
Coordinated Action of RTBV and RTSV Proteins Suppress Host RNA Silencing Machinery

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Small RNA Library Sequencing and Computational Analysis
2.3. Plasmid Constructs
2.4. Generation of GFP Silenced Tobacco Plants and Agro-Infiltration
2.5. cDNA Synthesis
2.6. Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
2.7. Northern Blot Analysis
2.8. Yeast Two-Hybrid Analysis
3. Results
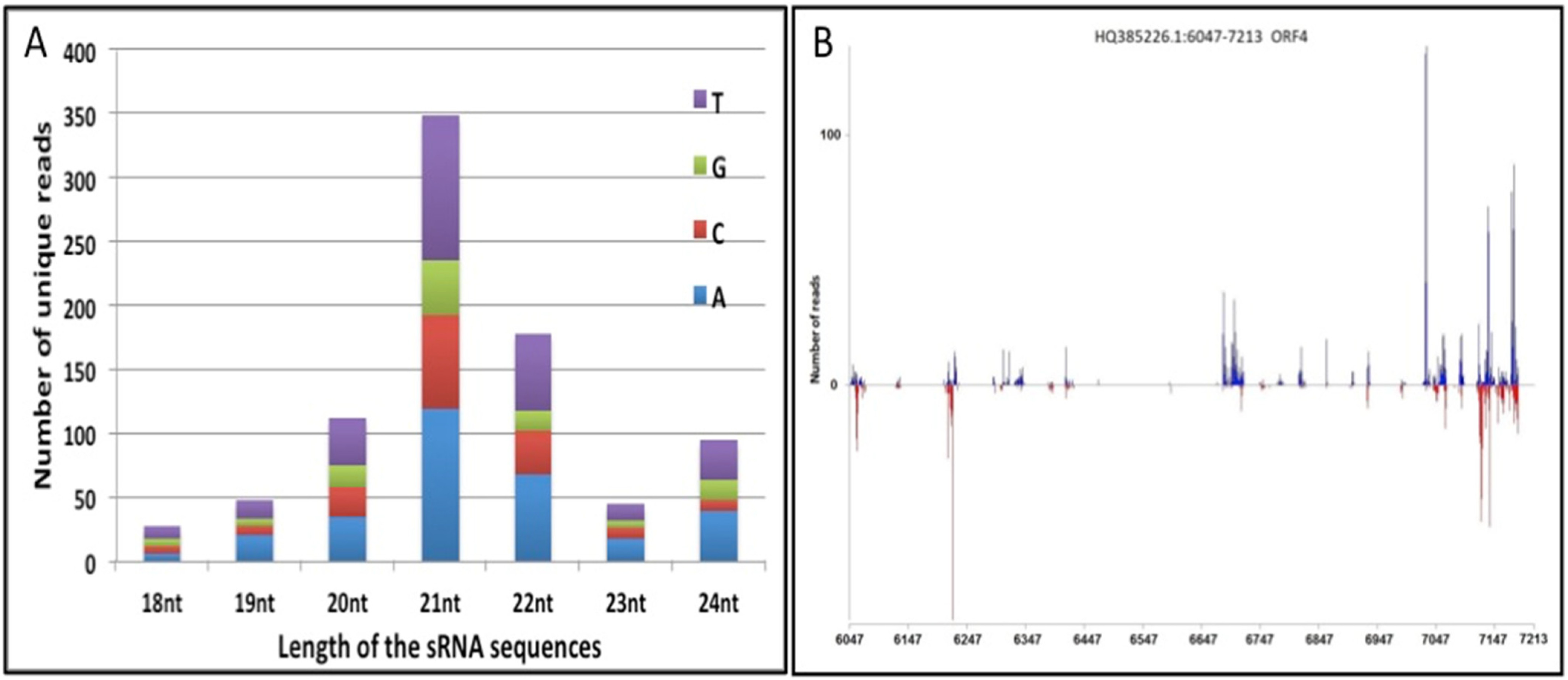
3.1. ORF-IV of RTBV Is a Potential Hotspot for siRNA Generation
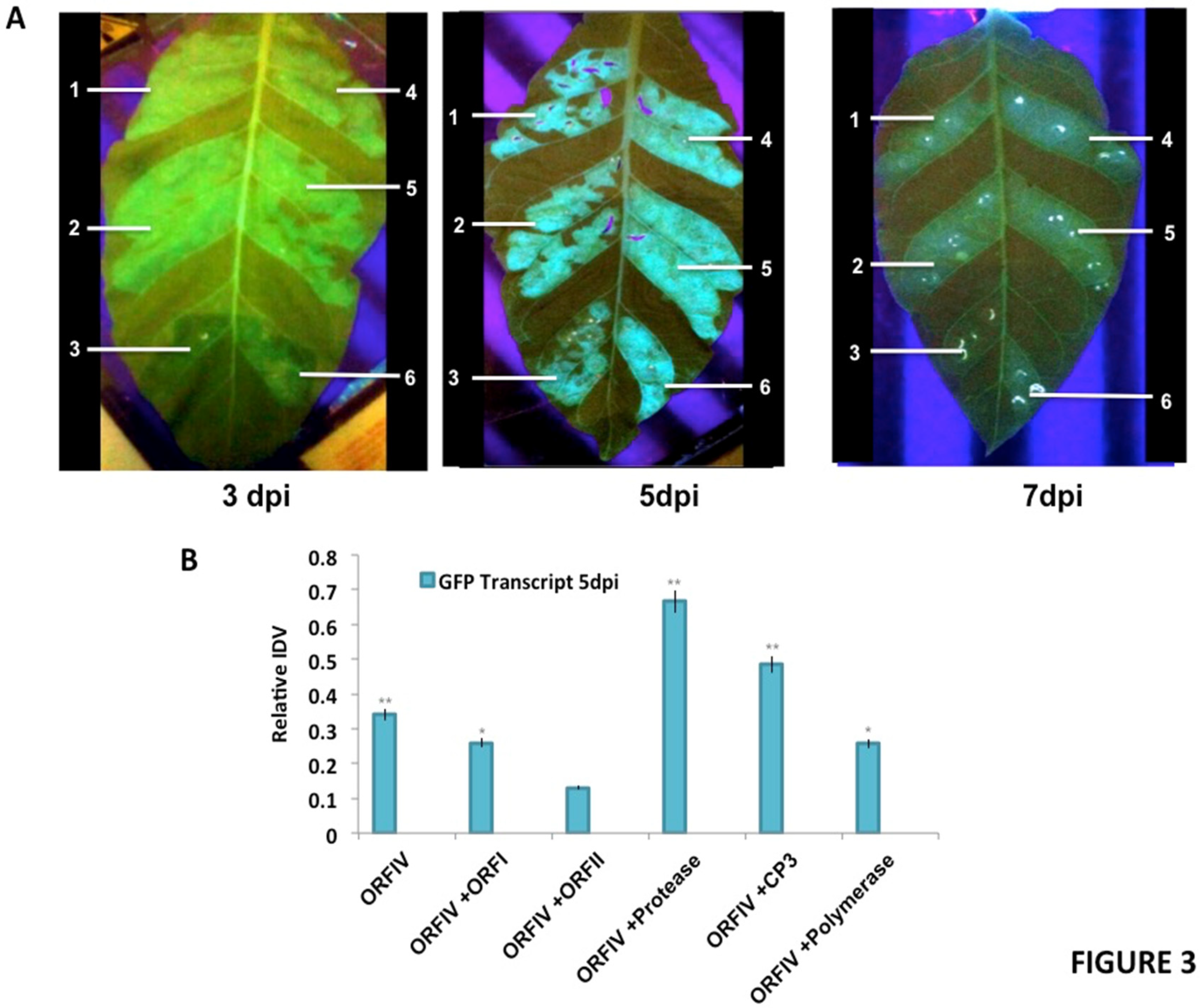
3.2. ORF-IV Can Suppress Pre-Established RNA Silencing
3.3. RTSV Coat Protein Enhances the Suppressor Activity of ORF-IV
3.4. RTBV ORF-IV Interacts with RTSV CP3 Protein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McKinney, H.H. Mosaic diseases of the Canary Islands; West Africa and Gibraltar. J. Agri. Res. 1929, 39, 557–578. [Google Scholar]
- Lindbo, J.A.; Silva-Rosales, L.; Proebsting, W.M.; Dougherty, W.G. Induction of a highly specific antiviral state in transgenic plants: Implications for regulation of gene expression and virus resistance. Plant Cell 1993, 5, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Beachy, R.N. Mechanisms and applications of pathogen-derived resistance in transgenic plants. Curr. Opin. Biotechnol. 1997, 8, 215–220. [Google Scholar] [CrossRef]
- Kyrychenko, A.M.; Kovalenko, O.G. Basic engineering strategies for virus-resistant plants. Cytol. Genet. 2018, 52, 213–221. [Google Scholar] [CrossRef]
- Ratcliff, F.; Harrison, B.D.; Baulcombe, D.C. A similarity between viral defense and gene silencing in plants. Science 1997, 276, 1558–1560. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Host factors against plant viruses. Mol. Plant Pathol. 2019, 20, 1588–1601. [Google Scholar] [CrossRef]
- Ramesh, S.V.; Yogindran, S.; Gnanasekaran, P.; Chakraborty, S.; Winter, S.; Pappu, H. RVirus and viroid-derived small RNAs as modulators of host gene expression: Molecular insights into pathogenesis. Front. Microbiol. 2021, 11, 614231. [Google Scholar] [CrossRef]
- Teixeira, R.M.; Ferreira, M.A.; Raimundo, G.; Fontes, E. Geminiviral triggers and suppressors of plant antiviral immunity. Microorganisms 2021, 9, 775. [Google Scholar] [CrossRef]
- Ding, S.W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef]
- Prasad, A.; Sharma, N.; Muthamilarasan, M.; Rana, S.; Prasad, M. Recent advances in small RNA mediated plant-virus interactions. Crit. Rev. Biotechnol. 2019, 39, 587–601. [Google Scholar] [CrossRef]
- Sanan-Mishra, N.; Jailani, A.A.K.; Mandal, B.; Mukherjee, S.K. Secondary siRNAs in plants: Biosynthesis; various functions; applications in virology. Front. Plant Sci. 2021, 12, 610283. [Google Scholar] [CrossRef]
- Sanan-Mishra, N.; Mukherjee, S.K. A peep into Plant miRNA world. Open Plant Sci. J. 2007, 1, 1–9. [Google Scholar] [CrossRef]
- Pelaez, P.; Sanchez, F. Small RNAs in plant defense responses during viral and bacterial interactions: Similarities and differences. Front. Plant Sci. 2013, 4, 343. [Google Scholar] [CrossRef] [Green Version]
- Sinha, V.; Anand, A.; Mukherjee, S.K.; Sanan-Mishra, N. RNAi based strategies for enhancing plant resistance to virus infection. In Advances in Biotechnology; Datta, A., Fakruddin, M., Iqbal, H.M.N., Abraham, J., Eds.; Open Access eBooks: Las Vegas, NV, USA, 2017; Volume 4. [Google Scholar]
- Qu, F.; Morris, T.J. Suppressors of RNA silencing encoded by plant viruses and their role in viral infections. FEBS Lett. 2005, 579, 5958–5964. [Google Scholar] [CrossRef] [Green Version]
- Anand, A.; Mukherjee, S.K.; Sanan-Mishra, N. Tools for pathogenicity: Virus encoded RNA silencing suppressors. In Recent Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Microbiology Book Series; Mendez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 751–763. [Google Scholar]
- Sanan-Mishra, N.; Chakraborty, S.; Gupta, D.; Mukherjee, S.K. RNAi suppressors: Biology and mechanisms. In Plant Epigenetics; RNA Technologies; Rajewsky, N., Jurga, S., Barciszewski, J., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2017; pp. 199–230. [Google Scholar]
- Carrington, J.C.; Kasschau, K.D.; Johansen, L.K. Activation and suppression of RNA silencing by plant viruses. Virology 2001, 281, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silhavy, D.; Molnar, A.; Lucioli, A.; Szittya, G.; Hornyik, C.; Tavazza, M.; Burgyan, J. A viral protein suppresses RNA silencing and binds silencing-generated; 21- to 25-nucleotide double-stranded RNAs. EMBO J. 2002, 21, 3070–3080. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Mishra, S.K.; Rehman, J.; Taneja, J.; Sundaresan, G.; Sanan-Mishra, N.; Mukherjee, S.K. Mungbean yellow mosaic Indian virus encoded AC2 protein suppresses RNA silencing by inhibiting Arabidopsis RDR6 and AGO1 activities. Virology 2015, 486, 158–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veluthambi, K.; Sunitha, S. Targets and mechanisms of geminivirus silencing suppressor protein AC2. Front. Microbiol. 2021, 12, 645419. [Google Scholar] [CrossRef]
- Anandalakshmi, R.; Pruss, G.J.; Ge, X.; Marathe, R.; Mallory, A.C.; Smith, T.H.; Vance, V.B. A viral suppressor of gene silencing in plants. Proc. Natl. Acad. Sci. USA 1998, 95, 13079–13084. [Google Scholar] [CrossRef] [Green Version]
- Pruss, G.; Ge, X.; Shi, X.M.; Carrington, J.C.; Vance, V.B. Plant viral synergism: The potyviral genome encodes a broad-range pathogenicity enhancer that transactivates replication of heterologous viruses. Plant Cell 1997, 9, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.W.; Lin, S.S.; Chen, K.C.; Yeh, S.D.; Chua, N.H. Discriminating mutations of HC-Pro of Zucchini yellow mosaic virus with differential effects on small RNA pathways involved in viral pathogenicity and symptom development. Mol. Plant-Microbe Interact. 2010, 23, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Palauqui, J.C.; Elmayan, T.; Pollien, J.M.; Vaucheret, H. Systemic acquired silencing: Transgene-specific post-transcriptional silencing is transmitted by grafting from silenced stocks to non-silenced scions. EMBO J. 1997, 16, 4738–4745. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Induction and suppression of RNA silencing: Insights from viral infections. Nat. Rev. Genet. 2005, 6, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Karjee, S.; Islam, M.N.; Mukherjee, S.K. Screening and identification of virus-encoded RNA silencing suppressors. Methods Mol. Biol. 2008, 442, 187–203. [Google Scholar] [PubMed]
- Azzam, O.; Chancellor, T.B. The biology; epidemiology; management of rice tungro disease in Asia. Plant Dis. 2002, 86, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Cabauatan, P.; Hibino, H. Transmission of rice tungro bacilliform and spherical viruses by Nephotettix virescens (Distant). Philipp. Phytopathol. 1985, 21, 103–109. [Google Scholar]
- Jones, M.; Gough, K.; Dasgupta, I.; Rao, B.S.; Cliffe, J.; Qu, R.; Shen, P.; Kaniewska, M.; Blakebrough, M.; Davies, J. Rice tungro disease is caused by an RNA and a DNA virus. J. Gen. Virol. 1991, 72, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Sanfaçon, H.; Wellink, J.; Le Gall, O.; Karasev, A.; van der Vlugt, R.; Wetzel, T. Secoviridae: A proposed family of plant viruses within the order Picornavirales that combines the families Sequiviridae and Comoviridae; the unassigned genera Cheravirus and Sadwavirus; the proposed genus Torradovirus. Arch. Virol. 2009, 154, 899–907. [Google Scholar] [CrossRef] [Green Version]
- Shen, P.; Kaniewska, M.; Smith, C.; Beachy, R. Nucleotide sequence and genomic organization of rice tungro spherical virus. Virology 1993, 193, 621–630. [Google Scholar] [CrossRef]
- Hull, R. Molecular biology of rice tungro viruses. Annu. Rev. Phytopathol. 1996, 34, 275–297. [Google Scholar] [CrossRef]
- Mangrauthia, S.K.; Malathi, P.; Agarwal, S.; Sailaja, B.; Singh, J.; Ramkumar, G.; Krishnaveni, D.; Balachandran, S.M. The molecular diversity and evolution of Rice tungro bacilliform virus from Indian perspective. Virus Genes 2012, 45, 126–138. [Google Scholar] [CrossRef]
- Kannan, M.; Saad, M.M.; Talip, N.; Baharum, S.N.; Bunawan, H. Complete genome sequence of Rice Tungro Bacilliform Virus infecting asian rice (Oryza sativa) in Malaysia. Microbiol. Resour. Announc. 2019, 8, e00262-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borah, B.K.; Sharma, S.; Kant, R.; Johnson, A.; Saigopal, D.V.R.; Dasgupta, I. Bacilliform DNA-containing plant viruses in the tropics: Commonalities within a genetically diverse group. Mol. Plant Pathol. 2013, 14, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, I.; Hull, R.; Eastop, S.; Poggi-Pollini, C.; Blakebrough, M.; Boulton, M.I.; Davies, J.W. Rice tungro bacilliform virus DNA independently infects rice after Agrobacterium-mediated transfer. J. Gen. Virol. 1991, 72, 1215–1221. [Google Scholar] [CrossRef]
- Srilatha, P.; Yousuf, F.; Methre, R.; Vishnukiran, T.; Agarwal, S.; Poli, Y.; Raghurami Reddy, M.; Vidyasagar, B.; Shanker, C.; Krishnaveni, D.; et al. Physical interaction of RTBV ORFI with D1 protein of Oryza sativa and Fe/Zn homeostasis play a key role in symptoms development during rice tungro disease to facilitate the insect mediated virus transmission. Virology 2019, 526, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, H.; Rajasubramaniam, S.; Rajam, M.V.; Dasgupta, I. RNA-interference in rice against Rice tungro bacilliform virus results in its decreased accumulation in inoculated rice plants. Transgenic Res. 2008, 17, 897–904. [Google Scholar] [CrossRef] [Green Version]
- Rajeswaran, R.; Golyaev, V.; Seguin, J.; Zvereva, A.S.; Farinelli, L.; Pooggin, M.M. Interactions of Rice Tungro Bacilliform Pararetrovirus and Its Protein P4 with Plant RNA-Silencing Machinery. Mol. Plant-Microbe Interact. 2014, 27, 1370–1378. [Google Scholar] [CrossRef] [Green Version]
- Mangrauthia, S.K.; Malathi, P.; Krishnaveni, D.; Reddy, C.S.; Viraktamath, B.C.; Balachandran, S.M.; Neeraja, C.N.; Biswal, S.K. Rapid detection of rice tungro spherical virus by RT-PCR and dot-blot hybridization. J. Mycol. Plant Pathol. 2010, 40, 445–449. [Google Scholar]
- Malathi, P.; Mangrauthia, S.K. Deciphering the multiplication behaviour of Rice tungro bacilliform virus by absolute quantitation through real-time PCR. Arch. Phytopathol. Plant Protect. 2013, 46, 2366–2375. [Google Scholar] [CrossRef]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid; transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]
- Das, S.S.; Sanan-Mishra, N. Comparative analysis of RNAi suppression activity of proteins from two disparate viruses. Am. J. Plant Sci. 2014, 5, 1789–1798. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 1989; Volume 1. [Google Scholar]
- Song, X.; Li, P.; Zhai, J.; Zhou, M.; Ma, L.; Liu, B.; Jeong, D.H.; Nakano, M.; Cao, S.; Liu, C. Roles of DCL4 and DCL3b in rice phased small RNA biogenesis. Plant J. 2012, 69, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, X.; Ding, S.W. Viral suppressors of RNA-based viral immunity: Host targets. Cell Host Microbe 2010, 8, 12–15. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Gu, L.; Song, X.; Cui, X.; Lu, Z.; Zhou, M.; Wang, L.; Hu, F.; Zhai, J.; Meyers, B.C. Dicer-like 3 produces transposable element-associated 24-nt siRNAs that control agricultural traits in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 3877–3882. [Google Scholar] [CrossRef] [Green Version]
- Llave, C. Virus-derived small interfering RNAs at the core of plant–virus interactions. Trends Plant Sci. 2010, 15, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Pantaleo, V. Plant RNA silencing in viral defence. Adv. Expt. Med. Biol. 2011, 722, 39–58. [Google Scholar]
- Rajeswaran, R.; Pooggin, M.M. RDR6-mediated synthesis of complementary RNA is terminated by miRNA stably bound to template RNA. Nuc. Acids Res. 2012, 40, 594–599. [Google Scholar] [CrossRef] [Green Version]
- Szittya, G.; Burgyán, J. RNA interference-mediated intrinsic antiviral immunity in plants. Curr. Top. Microbiol. Immunol. 2013, 371, 153–181. [Google Scholar] [PubMed]
- Hull, R. Plant Virology; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Aregger, M.; Borah, B.K.; Seguin, J.; Rajeswaran, R.; Gubaeva, E.G.; Zvereva, A.S.; Windels, D.; Vazquez, F.; Blevins, T.; Farinelli, L.; et al. Primary and secondary siRNAs in geminivirus-induced gene silencing. PLoS Pathog. 2012, 8, e1002941. [Google Scholar] [CrossRef] [PubMed]
- Rajeswaran, R.; Aregger, M.; Zvereva, A.S.; Borah, B.K.; Gubaeva, E.G.; Pooggin, M.M. Sequencing of RDR6-dependent double-stranded RNAs reveals novel features of plant siRNA biogenesis. Nuc. Acids Res. 2012, 40, 6241–6254. [Google Scholar] [CrossRef] [Green Version]
- Blevins, T.; Rajeswaran, R.; Shivaprasad, P.V.; Beknazariants, D.; Si-Ammour, A.; Park, H.S.; Vazquez, F.; Robertson, D.; Meins, F., Jr.; Hohn, T.; et al. Four plant Dicers mediate viral small RNA biogenesis and DNA virus induced silencing. Nuc. Acids Res. 2006, 34, 6233–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, A.; Voinnet, O.; Chappell, L.; Baulcombe, D. Two classes of short interfering RNA in RNA silencing. EMBO J. 2002, 21, 4671–4679. [Google Scholar] [CrossRef] [Green Version]
- Himber, C.; Dunoyer, P.; Moissiard, G.; Ritzenthaler, C.; Voinnet, O. Transitivity-dependent and independent cell-to-cell movement of RNA silencing. EMBO J. 2003, 22, 4523–4533. [Google Scholar] [CrossRef] [PubMed]
- Dunoyer, P.; Schott, G.; Himber, C.; Meyer, D.; Takeda, A.; Carrington, J.C.; Voinnet, O. Small RNA duplexes function as mobile silencing signals between plant cells. Science 2010, 328, 912–916. [Google Scholar] [CrossRef]
- Molnar, A.; Melnyk, C.W.; Bassett, A.; Hardcastle, T.J.; Dunn, R.; Baulcombe, D.C. Small silencing RNAs in plants are mobile and direct epigenetic modification in recipient cells. Science 2010, 328, 872–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fütterer, J.; Potrykus, I.; Brau, M.P.V.; Dasgupta, I.; Hull, R.; Hohn, T. Splicing in a plant Pararetrovirus. Virology 1994, 198, 663–670. [Google Scholar] [CrossRef]
- Wang, X.B.; Jovel, J.; Udomporn, P.; Wang, Y.; Wu, Q.; Li, W.X.; Gasciolli, V.; Vaucheret, H.; Ding, S.W. The 21-nucleotide; but not 22-nucleotide; viral secondary small interfering RNAs direct potent antiviral defense by two cooperative argonautes in Arabidopsis thaliana. Plant Cell 2011, 23, 1625–1638. [Google Scholar] [CrossRef] [Green Version]
- Blevins, T.; Rajeswaran, R.; Aregger, M.; Borah, B.K.; Schepetilnikov, M.; Baerlocher, L.; Farinelli, L.; Meins, F.; Hohn, T.; Pooggin, M.M. Massive production of small RNAs from a non-coding region of Cauliflower mosaic virus in plant defense and viral counter-defense. Nuc. Acids Res. 2011, 39, 5003–5014. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Sugiyama, K.; Nagano, H.; Mori, M.; Kaido, M.; Mise, K.; Tsuda, S.; Okuno, T. Identification of a novel RNA silencing suppressor; NSs protein of Tomato spotted wilt virus. FEBS Letts. 2002, 532, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Fusaro, A.F.; Matthew, L.; Smith, N.A.; Curtin, S.J.; Dedic-Hagan, J.; Ellacott, G.A.; Watson, J.M.; Wang, M.B.; Brosnan, C.; Carroll, B.J. RNA interference-inducing hairpin RNAs in plants act through the viral defence pathway. EMBO Rep. 2006, 7, 1168–1175. [Google Scholar] [CrossRef]
- Mallory, A.C.; Vaucheret, H. Functions of microRNAs and related small RNAs in plants. Nat. Genet. 2006, 38, S31–S36. [Google Scholar] [CrossRef]
- Moissiard, G.; Parizotto, E.A.; Himber, C.; Voinnet, O. Transitivity in Arabidopsis can be primed; requires the redundant action of the antiviral Dicer-like 4 and Dicer-like 2, is compromised by viral-encoded suppressor proteins. RNA 2007, 13, 1268–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. Plant Cell 1990, 2, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Di Serio, F.; Schöb, H.; Iglesias, A.; Tarina, C.; Bouldoires, E.; Meins, F. Sense-and antisense-mediated gene silencing in tobacco is inhibited by the same viral suppressors and is associated with accumulation of small RNAs. Proc. Natl. Acad. Sci. USA 2001, 98, 6506–6510. [Google Scholar] [CrossRef] [Green Version]
- Zvereva, A.S.; Pooggin, M.M. Silencing and innate immunity in plant defense against viral and non-viral pathogens. Viruses 2012, 4, 2578–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malathi, P.; Muzammil, S.A.; Krishnaveni, D.; Balachandran, S.M.; Mangrauthia, S.K. Coat protein 3 of Rice tungro spherical virus is the key target gene for development of RNAi mediated tungro disease resistance in rice. Agri Gene 2019, 12, 100084. [Google Scholar] [CrossRef]
- Mangrauthia, S.K.; Malathi, P.; Balachandran, S.M.; Reddy, C.S.; Viraktamath, B.C. Global analysis of Rice tungro spherical virus coat proteins reveals new roles in evolutionary consequences. J. Plant Biochem. Biotechnol. 2010, 19, 263–266. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anand, A.; Pinninti, M.; Tripathi, A.; Mangrauthia, S.K.; Sanan-Mishra, N. Coordinated Action of RTBV and RTSV Proteins Suppress Host RNA Silencing Machinery. Microorganisms 2022, 10, 197. https://doi.org/10.3390/microorganisms10020197
Anand A, Pinninti M, Tripathi A, Mangrauthia SK, Sanan-Mishra N. Coordinated Action of RTBV and RTSV Proteins Suppress Host RNA Silencing Machinery. Microorganisms. 2022; 10(2):197. https://doi.org/10.3390/microorganisms10020197
Chicago/Turabian StyleAnand, Abhishek, Malathi Pinninti, Anita Tripathi, Satendra Kumar Mangrauthia, and Neeti Sanan-Mishra. 2022. "Coordinated Action of RTBV and RTSV Proteins Suppress Host RNA Silencing Machinery" Microorganisms 10, no. 2: 197. https://doi.org/10.3390/microorganisms10020197
APA StyleAnand, A., Pinninti, M., Tripathi, A., Mangrauthia, S. K., & Sanan-Mishra, N. (2022). Coordinated Action of RTBV and RTSV Proteins Suppress Host RNA Silencing Machinery. Microorganisms, 10(2), 197. https://doi.org/10.3390/microorganisms10020197