



Role of Indigenous Bacteria in Corrosion of Two Types of Carbon Steel

Abstract
:1. Introduction
2. Materials and Methods
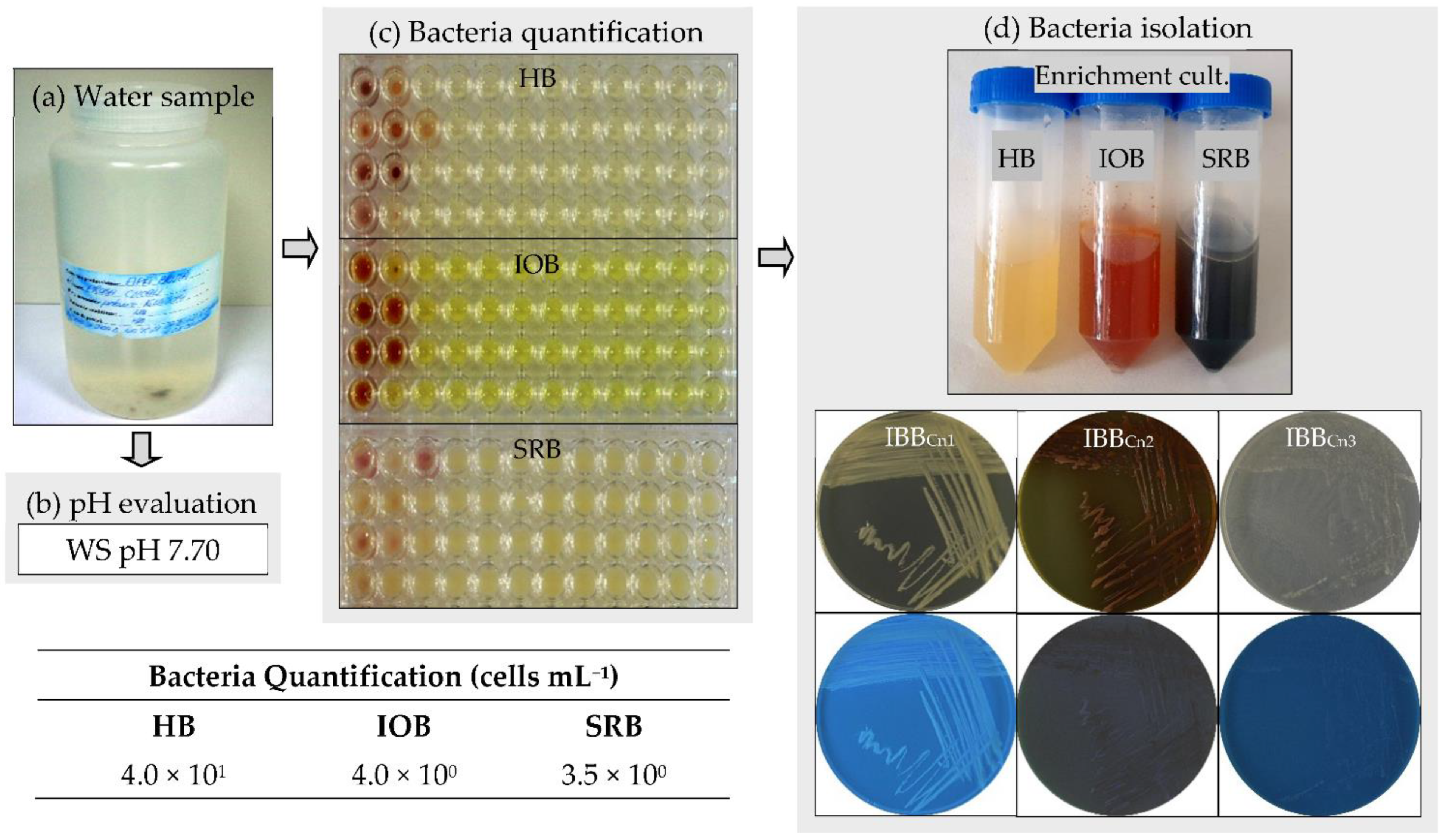
2.1. Bacteria Quantification in the Water Sample
2.2. Isolation and Identification of Bacteria from the Water Sample
2.2.1. Isolation of Bacteria
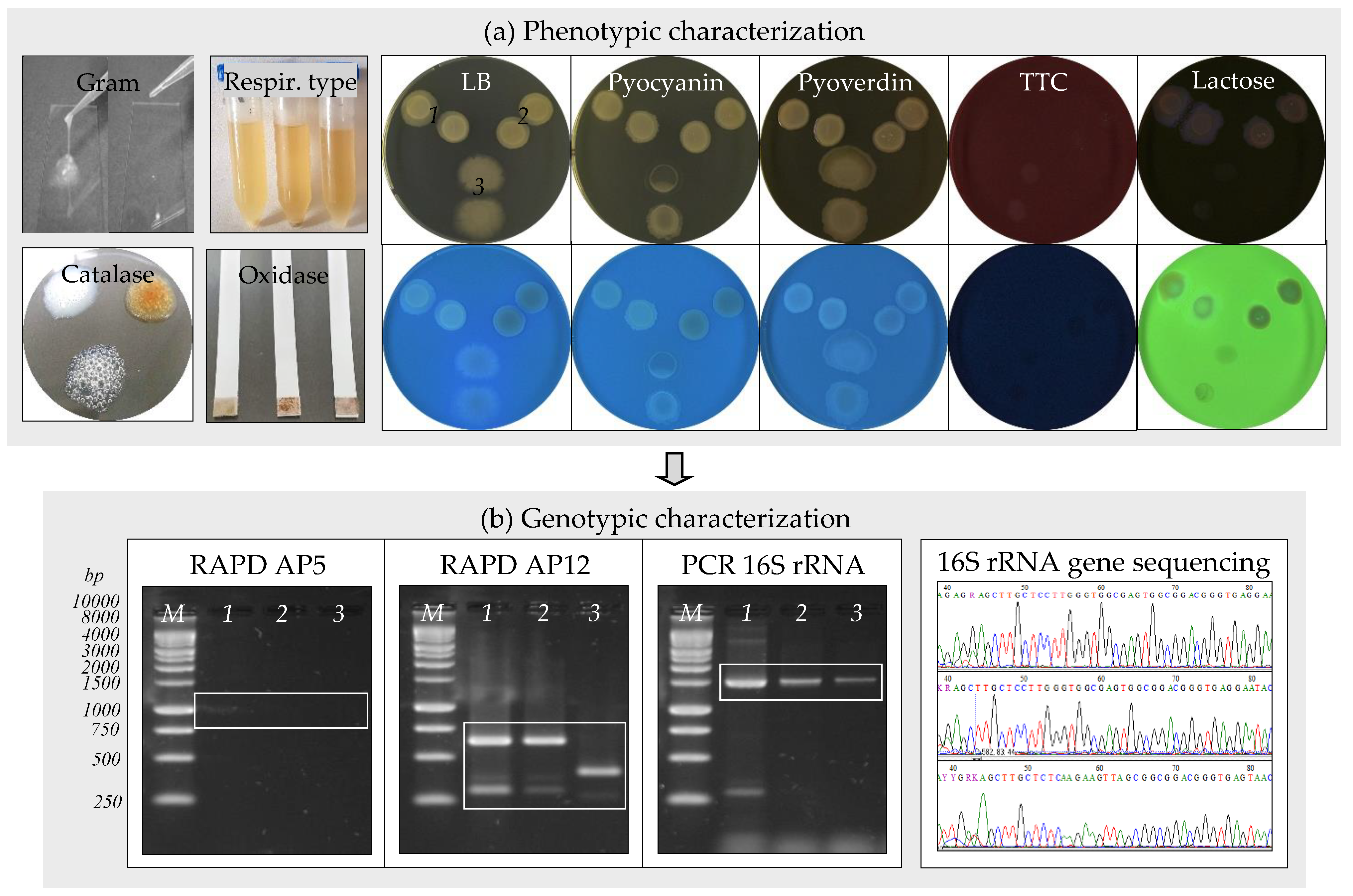
2.2.2. Identification of Bacteria
2.3. Biocorrosion of Carbon Steel Coupons Exposed to the Water Sample
2.3.1. Bacteria Quantification during the Corrosion Experiments
2.3.2. X-ray Fluorescence (XRF) Spectrometry
2.3.3. Isolation of Bacterial Consortia
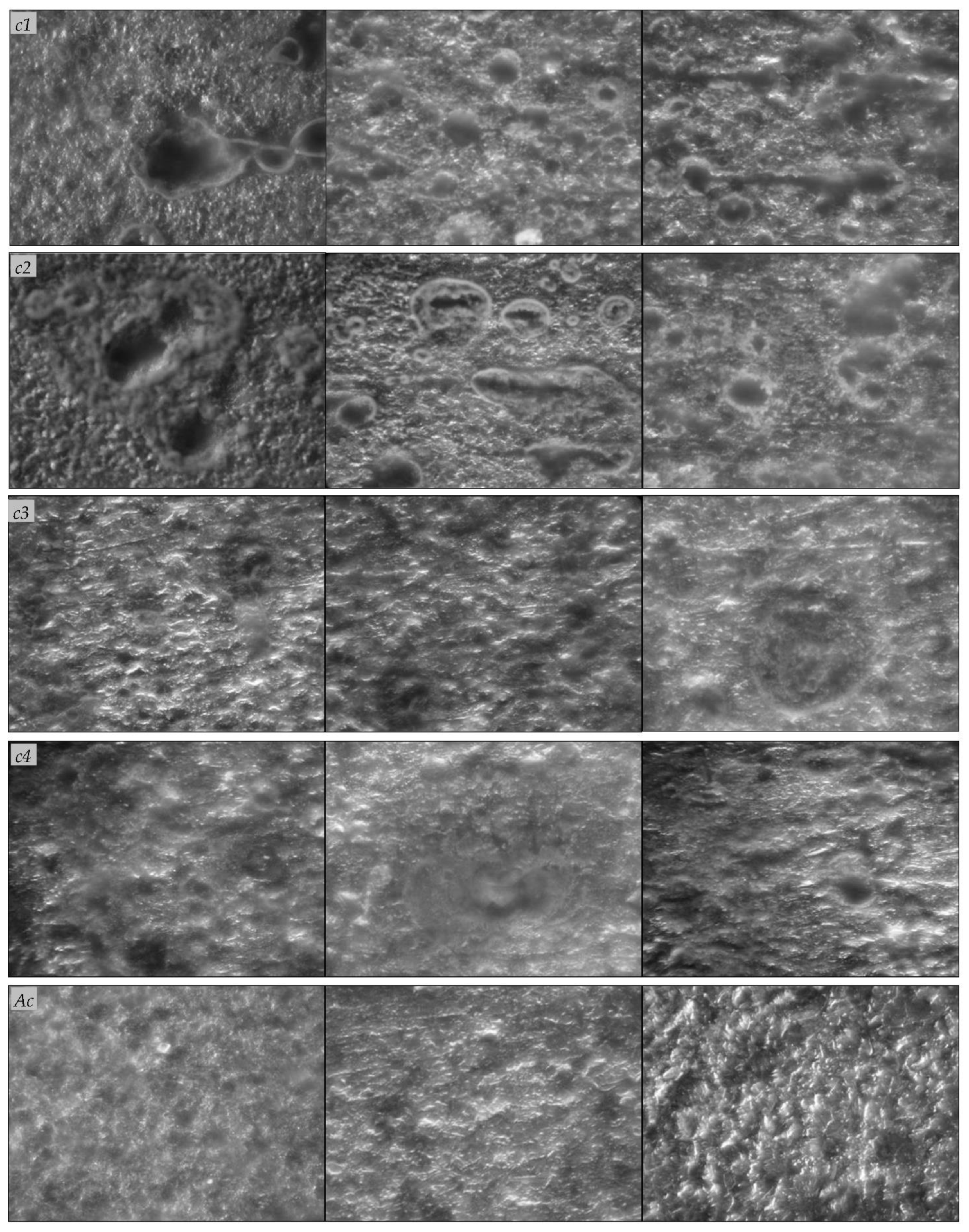
2.3.4. Optical Microscopy (OM), Scanning Electron Microscopy (SEM)
2.3.5. Weight Loss, Corrosion Rate
3. Results and Discussion
3.1. Bacteria Quantification in the Water Sample
3.2. Isolation and Identification of Bacteria from the Water Sample
3.3. Biocorrosion of Carbon Steel Coupons Exposed to the Water Sample
4. Conclusions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Melo, I.R.; Filho, S.L.U.; Oliveira, F.J.S.; de França, F.P. Formation of biofilms and biocorrosion on AISI-1020 carbon steel exposed to aqueous systems containing different concentrations of a diesel/biodiesel mixture. Int. J. Corros. 2011, 2011, 415920. [Google Scholar] [CrossRef] [Green Version]
- Zarasvand, K.A.; Rai, V.R. Microorganisms: Induction and inhibition of corrosion in metals. Int. Biodeterior. Biodegrad. 2014, 87, 66–74. [Google Scholar] [CrossRef]
- Refait, P.; Grolleau, A.-M.; Jeannin, M.; Rémazeilles, C.; Sabot, R. Corrosion of carbon steel in marine environments: Role of the corrosion product layer. Corros. Mater. Degrad. 2020, 1, 198–218. [Google Scholar] [CrossRef]
- Coetser, S.E.; Cloete, T.E. Biofouling and biocorrosion in industrial water systems. Crit. Rev. Microbiol. 2005, 31, 213–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, T. New understandings of biocorrosion mechanisms and their classifications. J. Microb. Biochem. Technol. 2012, 4, iii–vi. [Google Scholar] [CrossRef] [Green Version]
- Abdolahi, A.; Hamzah, E.; Ibrahim, Z.; Hashim, S. Microbially influenced corrosion of steels by Pseudomonas aeruginosa. Corros. Rev. 2014, 32, 129–141. [Google Scholar] [CrossRef]
- Černoušek, T.; Shrestha, R.; Kovářová, H.; Špánek, R.; Ševců, A.; Sihelská, K.; Kokinda, J.; Stoulil, J.; Steinová, J. Microbially influenced corrosion of carbon steel in the presence of anaerobic sulphate-reducing bacteria. Corros. Eng. Sci. Technol. 2019, 55, 127–137. [Google Scholar] [CrossRef]
- Phan, H.; Blackall, L.; Wade, S. Effect of multispecies microbial consortia on microbially influenced corrosion of carbon steel. Corros. Mater. Degrad. 2021, 2, 133–149. [Google Scholar] [CrossRef]
- Duan, J.; Wu, S.; Zhang, X.; Huang, G.; Du, M.; Hou, B. Corrosion of carbon steel influenced by anaerobic biofilm in natural seawater. Electrochim. Acta 2008, 54, 22–28. [Google Scholar] [CrossRef]
- McBeth, J.M.; Little, B.J.; Ray, R.I.; Farrar, K.M.; Emerson, D. Neutrophilic iron-oxidizing “Zetaproteobacteria” and mild steel corrosion in nearshore marine environments. Appl. Environ. Microb. 2011, 77, 1405–1412. [Google Scholar] [CrossRef]
- Li, X.; Duan, J.; Xiao, H.; Li, Y.; Liu, H.; Guan, F.; Zhai, X. Analysis of bacterial community composition of corroded steel immersed in Sanya and Xiamen Seawaters in China via method of Illumina MiSeq Sequencing. Front. Microbiol. 2017, 8, 1737. [Google Scholar] [CrossRef] [PubMed]
- Tribollet, B. Microbiologically influenced corrosion (MIC) in nuclear power plant systems and components. In Nuclear Corrosion Science and Engineering; Woodhead Publishing: Sawston, UK, 2012; pp. 230–261. [Google Scholar] [CrossRef]
- Féron, D.; Crusset, D. Microbial induced corrosion in French concept of nuclear waste underground disposal. Corros. Eng. Sci. Technol. 2014, 49, 540–547. [Google Scholar] [CrossRef]
- Lister, D.H.; Cook, W.G. Nuclear plant materials and corrosion. In The Essential CANDU; Garland, W.J., Ed.; UNENE: Canada, 2014; Available online: https://unene.ca/essentialcandu/pdf/14%20-%20Nuclear%20Plant%20Materials%20and%20Corrosion.pdf (accessed on 18 November 2022).
- Cai, D.; Wu, J.; Chai, K. Microbiologically influenced corrosion behavior of carbon steel in the presence of marine bacteria Pseudomonas sp. and Vibrio sp. ACS Omega 2021, 6, 3780–3790. [Google Scholar] [CrossRef] [PubMed]
- Rao, T.; Sairam, T.; Viswanathan, B.; Nair, K. Carbon steel corrosion by iron oxidising and sulphate reducing bacteria in a freshwater cooling system. Corros. Sci. 2000, 42, 1417–1431. [Google Scholar] [CrossRef]
- Stancu, M.M.; Grifoll, M. Multidrug resistance in hydrocarbon-tolerant Gram-positive and Gram-negative bacteria. J. Gen. Appl. Microbiol. 2011, 57, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Stancu, M.M. Isolation of bacterial consortia that induced corrosion of zirconium alloys. Water Air Soil Pollut. 2019, 230, 38. [Google Scholar] [CrossRef]
- Sambrook, J.; Russel, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Atlas, R.M. Handbook of Media for Environmental Microbiology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar] [CrossRef]
- Postgate, J.R. The Sulfate-Reducing Bacteria, 2nd ed.; Cambridge University Press: Cambridge, UK, 1984. [Google Scholar]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams and Wilkins: Baltimore, MD, USA, 1994. [Google Scholar]
- Pini, F.; Grossi, C.; Nereo, S.; Michaud, L.; Giudice, A.L.; Bruni, V.; Baldi, F.; Fani, R. Molecular and physiological characterisation of psychrotrophic hydrocarbon-degrading bacteria isolated from Terra Nova Bay (Antarctica). Eur. J. Soil Biol. 2007, 43, 368–379. [Google Scholar] [CrossRef]
- Michaud, L.; di Cello, F.; Brilli, M.; Fani, R.; Giudice, A.L.; Bruni, V. Biodiversity of cultivable Antarctic psychrotrophic marine bacteria isolated from Terra Nova Bay (Ross Sea). FEMS Microbiol. Lett. 2004, 230, 63–71. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Valencia-Cantero, E.; Peña-Cabriales, J.J. Effects of iron-reducing bacteria on carbon steel corrosion induced by thermophilic sulfate-reducing consortia. J. Microbiol. Biotechnol. 2014, 24, 280–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brânzoi, F.; Brânzoi, V.; Hărăbor, I. New friendly environment inhibitors for anticorrosive protection of cooling water systems. Rev. Roum. Chim. 2010, 55, 487–500. [Google Scholar]
- Chikere, C.B.; Okpokwasili, G.C.; Chikere, B.O. Monitoring of microbial hydrocarbon remediation in the soil. 3 Biotech 2011, 1, 117–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vartoukian, S.R.; Palmer, R.M.; Wade, W.G. Strategies for culture of ‘unculturable’ bacteria. FEMS Microbiol. Lett. 2010, 309, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Pedramfar, A.; Maal, K.B.; Mirdamadian, S.H. Characterization and identification of corrosive bacterium isolated from petroleum product transporting pipelines. Access Microbiol. 2019, 1, 130. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, H.; Karn, S.K.; Duan, J.; Guan, F.; Zhai, X.; Zhao, S.; Li, K.; Hou, B. Corrosion behavior of Q235 carbon steel in presence of H2S producing Bacillus sp. and a consortium of microbes isolated from inner rust layer. Int. J. Electrochem. Sci. 2017, 12, 2315–2328. [Google Scholar] [CrossRef]
- Procópio, L. Microbial community profiles grown on 1020 carbon steel surfaces in seawater-isolated microcosm. Ann. Microbiol. 2020, 70, 13. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Zhang, Y.; Cheng, G.; Zhu, W. Localized corrosion behavior of 316L stainless steel in the presence of sulfate-reducing and iron-oxidizing bacteria. Mater. Sci. Eng. A 2007, 443, 235–241. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Strain | |||
|---|---|---|---|---|
| IBBCn1 | IBBCn2 | IBBCn3 | ||
| Phenotypic | Color of colonies | creamy | red-yellow | white |
| Gram | − | − | + | |
| Shape | rods | rods | rods | |
| Motility | + | + | + | |
| Respiratory type | FAn | FAn | An | |
| Catalase | + | + | + | |
| Oxidase | − | − | − | |
| Hydrogen sulfide production | + | − | + | |
| Pyocyanin pigment production | − | − | − | |
| Pyoverdin pigment production | − | − | − | |
| Growth on TTC medium | − | − | − | |
| Lactose utilization | + | + | − | |
| Genotypic | RAPD using | |||
| AP5 primer (DFS, bp) | 1000 | − | − | |
| AP12 primer (DFS, bp) | 300, 400, 650 | 300, 400, 650 | 250, 420 | |
| PCR 16S rRNA gene using 27f-1492r primers (DFS, bp) | 1465 | 1465 | 1465 | |
| 16S rRNA gene sequence, sequence identity (%) | S. maltophilia, 99.78% | S. maltophilia, 99.78% | B. thuringiensis, 99.85% | |
| GenBank accession number | MT893712 | MT893713 | MT893714 | |
| CS Coupons | Bacteria Quantification (cells mL−1) | pH | Fe2O3 (wt.%) | CR (mm year−1) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| HB | IOB | SRB | |||||||||
| 0 Days | 120 Days | 0 Days | 120 Days | 0 Days | 120 Days | 0 Days | 120 Days | 0 Days | 120 Days | 120 Days | |
| c1 | 4.0 × 101 | 4.0 × 103 | 4.0 × 100 | 4.0 × 102 | 3.5 × 100 | 3.0 × 103 | 7.70 | 5.60 | 5.13 | 90.42 | 0.12 ± 0.03 |
| c2 | 4.0 × 101 | 3.0 × 103 | 4.0 × 100 | 3.0 × 102 | 3.5 × 100 | 4.0 × 104 | 7.70 | 5.80 | 5.13 | 86.97 | 0.13 ± 0.04 |
| c3 | 4.0 × 101 | 2.0 × 103 | 4.0 × 100 | 4.0 × 102 | 3.5 × 100 | 4.0 × 105 | 7.70 | 6.00 | 5.13 | 79.46 | 0.08 ± 0.01 |
| c4 | 4.0 × 101 | 4.0 × 102 | 4.0 × 100 | 3.0 × 102 | 3.5 × 100 | 5.0 × 105 | 7.70 | 6.02 | 5.13 | 90.19 | 0.10 ± 0.02 |
| Ac | − | − | − | − | − | − | 7.70 | 7.70 | 5.13 | 38.50 | 0.04 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stancu, M.M. Role of Indigenous Bacteria in Corrosion of Two Types of Carbon Steel. Microorganisms 2022, 10, 2451. https://doi.org/10.3390/microorganisms10122451
Stancu MM. Role of Indigenous Bacteria in Corrosion of Two Types of Carbon Steel. Microorganisms. 2022; 10(12):2451. https://doi.org/10.3390/microorganisms10122451
Chicago/Turabian StyleStancu, Mihaela Marilena. 2022. "Role of Indigenous Bacteria in Corrosion of Two Types of Carbon Steel" Microorganisms 10, no. 12: 2451. https://doi.org/10.3390/microorganisms10122451
APA StyleStancu, M. M. (2022). Role of Indigenous Bacteria in Corrosion of Two Types of Carbon Steel. Microorganisms, 10(12), 2451. https://doi.org/10.3390/microorganisms10122451