
Molecular Detection of Integrons, Colistin and β-lactamase Resistant Genes in Salmonella enterica Serovars Enteritidis and Typhimurium Isolated from Chickens and Rats Inhabiting Poultry Farms

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Salmonella serovars
2.2. Antibiotic Susceptibility Testing
2.3. Detection of Antibiotic Resistance Genes
2.4. Analysis of Antimicrobial Resistance Genes
2.4.1. Detection of Colistin (mcr)
2.4.2. Detection of β-lactamase Genes
2.4.3. Detection of Integrons (IntI) Genes
3. Results
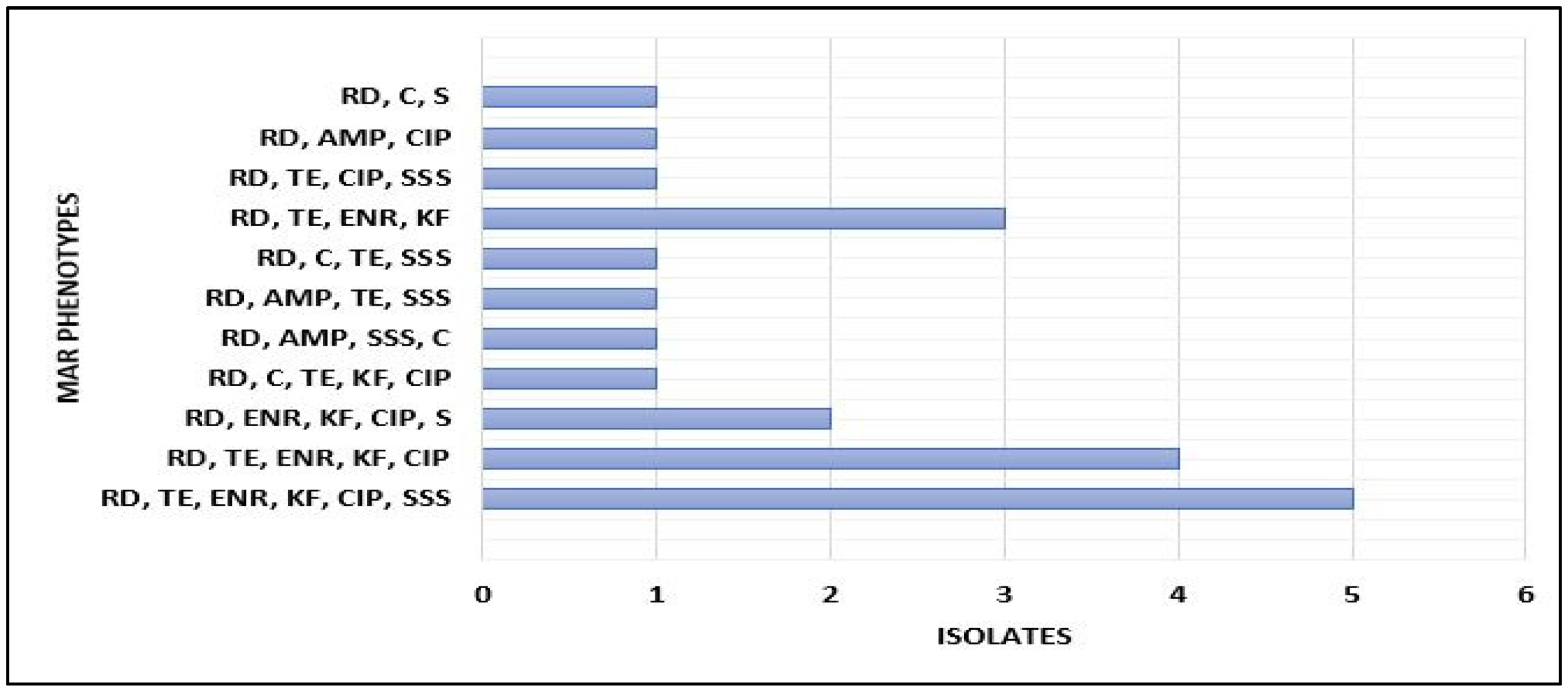
3.1. Antimicrobial Susceptibility Testing
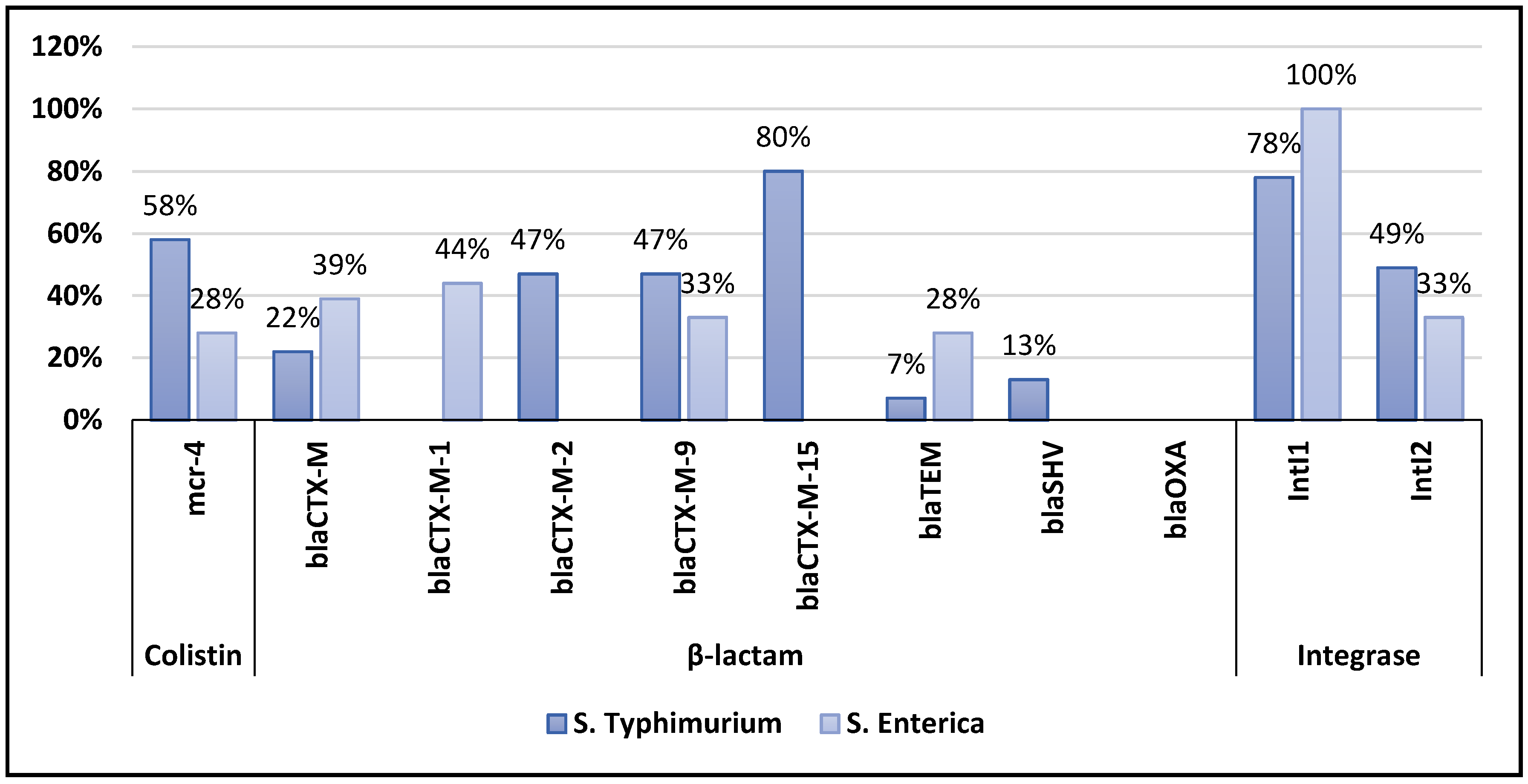
3.2. Detection of Antibiotic Resistance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waldman, J.; Souza, M.N.; Fonseca, A.S.K.; Ikuta, N.; Lunge, V.R. Direct detection of Salmonella from poultry samples by DNA isothermal amplification. Br. Poult. Sci. 2020, 61, 653–659. [Google Scholar] [CrossRef]
- Arkali, A.; Çetinkaya, B. Molecular identification and antibiotic resistance profiling of Salmonella species isolated from chickens in eastern Turkey. BMC Veter. Res. 2020, 16, 205. [Google Scholar] [CrossRef] [PubMed]
- Motladiile, T.W.; Tumbo, J.M.; Malumba, A.; Adeoti, B.; Masekwane, N.J.; Mokate, O.M.; Sebekedi, O.C. Salmonella food-poisoning outbreak linked to the National School Nutrition Programme, North West province, South Africa. S. Afr. J. Infect. Dis. 2019, 34, 6. [Google Scholar] [CrossRef] [PubMed]
- Keddy, K.H.; Takuva, S.; Musekiwa, A.; Puren, A.J.; Sooka, A.; Karstaedt, A.; Klugman, K.P.; Angulo, F.J. An association between decreasing incidence of invasive non-typhoidal salmonellosis and increased use of antiretroviral therapy, Gauteng Province, South Africa, 2003–2013. PLoS ONE 2017, 12, e0173091. [Google Scholar] [CrossRef]
- Shonhiwa, A.; Ntshoe, G.; Essel, V.; Thomas, J.; McCarthy, K. A review of foodborne diseases outbreaks reported to the outbreak response unit, national institute for communicable diseases, South Africa, 2013–2017. Int. J. Infect. Dis. 2019, 79, 73. [Google Scholar] [CrossRef] [Green Version]
- Esperón, F.; Sacristán, C.; Carballo, M.; de la Torre, A. Antimicrobial resistance genes in animal manure, manure-amended and nonanthropogenically impacted soils in Spain. Adv. Biosci. Biotechnol. 2018, 9, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Moreno, M.O.; Picó-Plana, E.; de Toro, M.; Grande-Armas, J.; Quiles-Fortuny, V.; Pons, M.J.; Gomes, C.; Sáenz, Y.; Torres, C.; Ruiz, J. β-Lactamases, transferable quinolone resistance determinants, and class 1 integron-mediated antimicrobial resistance in human clinical Salmonella enterica isolates of non-Typhimurium serotypes. Int. J. Med. Microbiol. 2013, 303, 25–31. [Google Scholar] [CrossRef]
- Ramatla, T.; Taioe, M.O.; Thekisoe, O.M.; Syakalima, M. Confirmation of Antimicrobial Resistance by Using Resistance Genes of Isolated Salmonella spp. in Chicken Houses of North West, South Africa. J. World’s Poult. Res. 2019, 9, 158–165. [Google Scholar] [CrossRef]
- Nghiem, M.N.; Nguyen, V.T.; Jeung, E.; Vo, T.T.B. Alternate antimicrobial resistance genes in multidrug resistant Salmonella spp. isolated from retail meats in Vietnam using RNA-sequencing analysis. J. Food Saf. 2019, 39, 12707. [Google Scholar] [CrossRef]
- Miró, E.; Vergés, C.; García, I.; Mirelis, B.; Navarro, F.; Coll, P.; Prats, G.; Martínez-Martínez, L. Resistance to quinolones and beta-lactams in Salmonella enterica due to mutations in topoisomerase-encoding genes, altered cell permeability and expression of an active efflux system. Enferm. Infecc. Microbiol. Clin. 2004, 22, 204–211. [Google Scholar] [CrossRef]
- Hasman, H.; Mevius, D.; Veldman, K.; Olesen, I.; Aarestrup, F. β-Lactamases among extended-spectrum β-lactamase (ESBL)-resistant Salmonella from poultry, poultry products and human patients in The Netherlands. J. Antimicrob. Chemother. 2005, 56, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Tzouvelekis, L.; Tzelepi, E.; Tassios, P.T.; Legakis, N. CTX-M-type β-lactamases: An emerging group of extended-spectrum enzymes. Int. J. Antimicrob. Agents 2000, 14, 137–142. [Google Scholar] [CrossRef]
- Olesen, I.; Hasman, H.; Møller Aarestrup, F. Prevalence of β-lactamases among ampicillin-resistant Escherichia coli and Salmonella isolated from food animals in Denmark. Microb. Drug Resist. 2004, 10, 334–340. [Google Scholar] [CrossRef]
- Zhao, S.; Blickenstaff, K.; Glenn, A.; Ayers, S.L.; Friedman, S.L.; Abbott, J.W.; McDermott, P.F. β-Lactam Resistance in Salmonella Strains Isolated from Retail Meats in the United States by the National Antimicrobial Resistance Monitoring System between 2002 and 2006. Appl. Environ. Microbiol. 2009, 75, 7624–7630. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Maamar, E.; Alonso, C.A.; Hamzaoui, Z.; Dakhli, N.; Abbassi, M.S.; Ferjani, S.; Saidani, M.; Boubaker, I.B.-B.; Torres, C. Emergence of plasmid-mediated colistin-resistance in CMY-2-producing Escherichia coli of lineage ST2197 in a Tunisian poultry farm. Int. J. Food Microbiol. 2018, 269, 60–63. [Google Scholar] [CrossRef] [PubMed]
- El Garch, F.; de Jong, A.; Bertrand, X.; Hocquet, D.; Sauget, M. mcr-1-like detection in commensal Escherichia coli and Salmonella spp. from food-producing animals at slaughter in Europe. Veter-Microbiol. 2018, 213, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Snyman, Y.; Whitelaw, A.C.; Barnes, J.M.; Maloba, M.R.B.; Newton-Foot, M. Characterisation of mobile colistin resistance genes (mcr-3 and mcr-5) in river and storm water in regions of the Western Cape of South Africa. Antimicrob. Resist. Infect. Control. 2021, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Anyanwu, M.U.; Jaja, I.F.; Oguttu, J.W.; Jaja, C.J.; Chah, K.F.; Shoyinka, V.S. Is Africa ready for mobile colistin resistance threat? Infect. Ecol. Epidemiol. 2021, 11, 1962781. [Google Scholar] [CrossRef]
- Kumar, H.; Chen, B.-H.; Kuca, K.; Nepovimova, E.; Kaushal, A.; Nagraik, R.; Bhatia, S.K.; Dhanjal, D.S.; Kumar, V.; Kumar, A.; et al. Understanding of Colistin Usage in Food Animals and Available Detection Techniques: A Review. Animals 2020, 10, 1892. [Google Scholar] [CrossRef] [PubMed]
- Drali, R.; Berrazeg, M.; Zidouni, L.L.; Hamitouche, F.; Abbas, A.A.; Deriet, A.; Mouffok, F. Emergence of mcr-1 plasmid-mediated colistin-resistant Escherichia coli isolates from seawater. Sci. Total Environ. 2018, 642, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance integrons: Class 1, 2 and 3 integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Tosini, F.; Giles, W.P.; Rupp, M.E.; Hinrichs, S.H.; Angulo, F.J.; Barrett, T.J.; Fey, P.D. Characterization of Plasmids Carrying CMY-2 from Expanded-Spectrum Cephalosporin-Resistant Salmonella Strains Isolated in the United States between 1996 and 1998. Antimicrob. Agents Chemother. 2002, 46, 1269–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kargar, M.; Mohammadalipour, Z.; Doosti, A.; Lorzadeh, S.; Japoni-Nejad, A. High Prevalence of Class 1 to 3 Integrons among Multidrug-Resistant Diarrheagenic Escherichia coli in Southwest of Iran. Osong Public Health Res. Perspect. 2014, 5, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Goudarzi, M.; Azimi, H. Dissemination of Classes 1, 2, and 3 Integrons in Acinetobacter baumannii Strains Recovered from Intensive Care Units Using Polymerase Chain Reaction-Restriction Fragment Length Polymorphism. Jundishapur J. Microbiol. 2017, 10, 193–198. [Google Scholar] [CrossRef]
- Faldynova, M.; Pravcova, M.; Sisak, F.; Havlickova, H.; Kolackova, I.; Cizek, A.; Karpiskova, R.; Rychlik, I. Evolution of Antibiotic Resistance in Salmonella enterica Serovar Typhimurium Strains Isolated in the Czech Republic between 1984 and 2002. Antimicrob. Agents Chemother. 2003, 47, 2002–2005. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Linke, L.; Doster, E.; Hyatt, D.; Burgess, B.A.; Magnuson, R.; Pabilonia, K.L.; Morley, P.S. Genomic diversity of class I integrons from antimicrobial resistant strains of Salmonella typhimurium isolated from livestock, poultry and humans. PLoS ONE 2020, 15, e0243477. [Google Scholar] [CrossRef]
- Ehuwa, O.; Jaiswal, A.; Jaiswal, S. Salmonella, Food Safety and Food Handling Practices. Foods 2021, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wang, C.; Ye, Y.; Liu, Y.; Wang, A.; Li, Y.; Zhou, X.; Pan, H.; Zhang, J.; Xu, X. Molecular Identification of Multidrug-Resistant Campylobacter Species from Diarrheal Patients and Poultry Meat in Shanghai, China. Front. Microbiol. 2018, 9, 1642. [Google Scholar] [CrossRef]
- Zhao, X.; Hu, M.; Zhang, Q.; Zhao, C.; Zhang, Y.; Li, L.; Qi, J.; Luo, Y.; Zhou, D.; Liu, Y. Characterization of integrons and antimicrobial resistance in Salmonella from broilers in Shandong, China. Poult. Sci. 2020, 99, 7046–7054. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Shimamoto, T. Genetic analysis of multiple antimicrobial resistance in Salmonella isolated from diseased broilers in Egypt. Microbiol. Immunol. 2012, 56, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Ramatla, T.; Mphuthi, N.; Gofaone, K.; Taioe, M.O.; Thekisoe, O.M.M.; Syakalima, M. Identification of Rodent Species That Infest Poultry Houses in Mafikeng, North West Province, South Africa. Int. J. Zool. 2019, 2019, 1280578. [Google Scholar] [CrossRef]
- Sekyere, J.O. Current State of Resistance to Antibiotics of Last-Resort in South Africa: A Review from a Public Health Perspective. Front. Public Health 2016, 4, 209. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance 2018, 23, 17-00672. [Google Scholar] [CrossRef] [PubMed]
- Firoozeh, F.; Zahraei-Salehi, T.; Shahcheraghi, F.; Karimi, V.; Aslani, M.M. Characterization of class I integrons among Salmonella enterica serovar Enteritidis isolated from humans and poultry. FEMS Immunol. Med. Microbiol. 2011, 64, 237–243. [Google Scholar] [CrossRef]
- Mobaraki, S.; Aghazadeh, M.; Barhaghi, M.H.S.; Memar, M.Y.; Goli, H.R.; Gholizadeh, P.; Kafil, H.S. Prevalence of integrons 1, 2, 3 associated with antibiotic resistance in Pseudomonas aeruginosa isolates from Northwest of Iran. BioMedicine 2018, 8, 12–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaja, I.F.; Bhembe, N.L.; Green, E.; Oguttu, J.; Muchenje, V. Molecular characterisation of antibiotic-resistant Salmonella enterica isolates recovered from meat in South Africa. Acta Trop. 2019, 190, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Gargano, V.; Sciortino, S.; Gambino, D.; Costa, A.; Agozzino, V.; Reale, S.; Alduina, R.; Vicari, D. Antibiotic Susceptibility Profile and Tetracycline Resistance Genes Detection in Salmonella spp. Strains Isolated from Animals and Food. Antibiotics 2021, 10, 809. [Google Scholar] [CrossRef]
- Aduah, M.; Adzitey, F.; Amoako, D.; Abia, A.; Ekli, R.; Teye, G.; Shariff, A.; Huda, N. Not All Street Food Is Bad: Low Prevalence of Antibiotic-Resistant Salmonella enterica in Ready-to-Eat (RTE) Meats in Ghana Is Associated with Good Vendors’ Knowledge of Meat Safety. Foods 2021, 10, 1011. [Google Scholar] [CrossRef] [PubMed]
- Mokgophi, T.; Gcebe, N.; Fasina, F.; Adesiyun, A. Antimicrobial Resistance Profiles of Salmonella Isolates on Chickens Processed and Retailed at Outlets of the Informal Market in Gauteng Province, South Africa. Pathogens 2021, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Eguale, T.; Birungi, J.; Asrat, D.; Njahira, M.N.; Njuguna, J.; Gebreyes, W.A.; Gunn, J.S.; Djikeng, A.; Engidawork, E. Genetic markers associated with resistance to beta-lactam and quinolone antimicrobials in non-typhoidal Salmonella isolates from humans and animals in central Ethiopia. Antimicrob. Resist. Infect. Control. 2017, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabry, M.A.; Abdel-Moein, K.A.; Abdel-Kader, F.; Hamza, E. Extended-spectrum β-lactamase-producing Salmonella serovars among healthy and diseased chickens and their public health implication. J. Glob. Antimicrob. Resist. 2020, 22, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Raseala, C.M.; Ekwanzala, M.D.; Momba, M.N.B. Shared Extended-Spectrum β-Lactamase-Producing Salmonella Serovars between Agricultural and Aquatic Environments Revealed through invA Amplicon Sequencing. Microorganisms 2020, 8, 1898. [Google Scholar] [CrossRef]
- Sow, A.G.; Wane, A.A.; Diallo, M.H.; Boye, C.S.-B.; Aïdara-Kane, A. Genotypic characterization of antibiotic-resistant Salmonella enteritidis isolates in Dakar, Senegal. J. Infect. Dev. Ctries. 2007, 1, 284–288. [Google Scholar] [PubMed]
- Boisramé, S.; Tandé, D.; Münck, M.-R.; Gouriou, S.; Nordmann, P.; Naas, T. Salmonella carriage in adopted children from Mali: 2001-08. J. Antimicrob. Chemother. 2011, 66, 2271–2276. [Google Scholar] [CrossRef]
- Adesiji, Y.O.; Deekshit, V.K.; Karunasagar, I. Antimicrobial-resistant genes associated with Salmonella spp. isolated from human, poultry, and seafood sources. Food Sci. Nutr. 2014, 2, 436–442. [Google Scholar] [CrossRef]
- Mendelson, M.; Brink, A.; Gouws, J.; Mbelle, N.; Naidoo, V.; Pople, T.; Schellack, N.; van Vuuren, M.; Rees, H.; Banoo, S.; et al. The One Health stewardship of colistin as an antibiotic of last resort for human health in South Africa. Lancet Infect. Dis. 2018, 18, e288–e294. [Google Scholar] [CrossRef]
- National Institute for Communicable Diseases. Antimicrobial Resistance National Surveillance Maps for South Africa. Available online: http://www.nicd.ac.za/index.php/httpscdwmicrostrategy-nhls-aczamicrostrategyaspmain-aspxservernhlssandmicsi03projectnhlsbiport0evt2048001srcmain-aspx-2048001documentid10c27d0e4b424b217efdc086730cd76dcurrentvie/ (accessed on 6 March 2018).
- Coetzee, J.; Corcoran, C.; Prentice, E.; Moodley, M.; Mendelson, M.; Poirel, L.; Nordmann, P.; Brink, A.J. Emergence of plasmid-mediated colistin resistance (MCR-1) among Escherichia coli isolated from South African patients. S. Afr. Med. J. 2016, 106, 449–450. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance 2017, 22, 30589. [Google Scholar] [CrossRef] [Green Version]
- Mthembu, T.; Zishiri, O.; El Zowalaty, M. Genomic Characterization of Antimicrobial Resistance in Food Chain and Livestock-Associated Salmonella Species. Animals 2021, 11, 872. [Google Scholar] [CrossRef] [PubMed]
- Hassell, J.M.; Ward, M.J.; Muloi, D.; Bettridge, J.; Robinson, T.P.; Kariuki, S.; Ogendo, A.; Kiiru, J.; Imboma, T.; Kang’Ethe, E.K.; et al. Clinically relevant antimicrobial resistance at the wildlife–livestock–human interface in Nairobi: An epidemiological study. Lancet Planet. Health 2019, 3, e259–e269. [Google Scholar] [CrossRef] [Green Version]
- Sonola, V.S.; Katakweba, A.S.; Misinzo, G.; Matee, M.I.N. Occurrence of Multi-Drug-Resistant Escherichia coli in Chickens, Humans, Rodents and Household Soil in Karatu, Northern Tanzania. Antibiotics 2021, 10, 1137. [Google Scholar] [CrossRef] [PubMed]
- Agyare, C.; Boamah, V.E.; Zumbi, C.N.; Osei, F.B. Antibiotic Use in Poultry Production and Its Effects on Bacterial Resistance. In Antimicrobial Resistance—A Global Threat; Kumar, Y., Ed.; IntechOpen: London, UK, 2018; pp. 33–51. [Google Scholar]
- Antunes, P.; Machado, J.; Peixe, L. Characterization of antimicrobial resistance and class 1 and 2 integrons in Salmonella enterica isolates from different sources in Portugal. J. Antimicrob. Chemother. 2006, 58, 297–304. [Google Scholar] [CrossRef]
- Lu, Y.; Zhao, H.; Sun, J.; Liu, Y.; Zhou, X.; Beier, R.C.; Wu, G.; Hou, X. Characterization of Multidrug-Resistant Salmonella enterica Serovars Indiana and Enteritidis from Chickens in Eastern China. PLoS ONE 2014, 9, e96050. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, F.E.L.; Dantas, F.G.D.S.; Grisolia, A.B.; Crispim, B.D.A.; Oliveira, K.M.P. Identification of class 1 and 2 integrons from clinical and environmental Salmonella isolates. J. Infect. Dev. Ctries. 2014, 8, 1518–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firoozeh, F.; Shahcheraghi, F.; Salehi, T.Z.; Karimi, V.; Aslani, M.M. Antimicrobial resistance profile and presence of class I integrongs among Salmonella enterica serovars isolated from human clinical specimens in Tehran, Iran. Iran. J. Microbiol. 2011, 3, 112–117. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Target Gene | Primer | Primer Sequence (5′ → 3′) | Conditions | Cycles | Size (bp) | References |
|---|---|---|---|---|---|---|
| Colistin resistance | ||||||
| mcr-1 | mcr-1-F mcr-1-R | TATCGCTATGTGCTAAAGCCTG CGTCTGCAGCCACTGGG | 94 °C for 5 Min, 94 °C for 30 s, 56 °C for 1 min, 72 °C for 1 min, 72 °C for 5 min. | 25 | 1139 | [34] |
| mcr-2 | mcr-2-F mcr-2-R | TATCGCTATGTGCTAAAGCCTG AAAATACTGCGTGGCAGGTAGC | 816 | |||
| mcr-3 | mcr-3-F mcr-3-R | CAATCGTTAGTTACACAATGATGAAG AACACATCTAGCAGGCCCTC | 676 | |||
| mcr-4 | mcr-4-F mcr-4-R | ATCCTGCTGAAGCATTGATG GCGCGCAGTTTCACC | 405 | |||
| mcr-5 | mcr-5-F mcr-5-R | GGTTGAGCGGCTATGAAC GAATGTTGACGTCACTACGG | 207 | |||
| β-lactamase resistance | ||||||
| blaCTX-M | blaCTX-M -F blaCTX-M- R | GTTACAATGTGTGAGAAGCAG CCGTTTCCGCTATTACAAAC | 94 °C for 5 min, 94 °C for 45 s, 55 °C for 30 s, 72 °C for 60 s, 72 °C for 10 min. | 35 | 550 | [35] |
| blaCTX-M-1 | blaCTX-M-1 -F blaCTX-M-1 -R | GTTACAATGTGTGAGAAGCAG CCGTTTCCGCTATTACAAAC | 1041 | |||
| blaCTX-M-2 | blaCTX-M-2 -F blaCTX-M-2 -R | ATGATGACTCAGAGCATTCGCCGC TCAGAAACCGTGGGTTACGATTT | 876 | |||
| blaCTX-M-15 | blaCTX-M-15 -F blaCTX-M-15 -R | CACACGTGGAATTTAGGGACT GCCGTCTAAGGCGATAAACA | 995 | |||
| blaTEM | blaTEM -F blaTEM -R | TTCTTGAAGACGAAAGGG C ACGCTCAGTGGAACGAAAAC | 1150 | |||
| blaSHV | blaSHV -F blaSHV -R | CACTCAAGGATGTATTGT G TTAGCGTTGCCAGTGCTCG | 885 | |||
| blaOXA | blaOXA- F blaOXA -R | ACACAATACATATCAACTTCGC AGTGTGTTTAGAATGGTGATC | 813 | |||
| Integrase Class 1, 2 and 3 | ||||||
| IntI1 | IntI1-F IntI1-R | GCCTTGCTGTTCTTCTACGG GATGCCTGCTTGTTCTACGG | 94 °C for 5 min, 30 s at 94 °C, 30 s, 55–60 °C, 2 min at 72 °C, 5 min at 72 °C. | 35 | 558 | [36] |
| IntI2 | IntI2-F IntI2-R | CACGGATATGCGACAAAAAGG TGTAGCAAACGAGTGACGAAATG | 94 °C for 5 min, 94 °C for 1 min, 60 °C for 1 min, 72 °C for 2 min, 72 °C for 10 min. | 32 | 740 | [25] |
| IntI3 | IntI3-F IntI3-R | GCCTCCGGCAGCGACTTTCAG ACGGATCTGCCAAACCTGACT | 94 °C for 10 min, 94 °C for 40 s, 59 °C for 50 s and 72 °C for 55 s 72 °C for 10 min. | 30 to 40 | 650 | [37] |
| No. Resistant (%) | |||||
|---|---|---|---|---|---|
| Antibiotic | Code | Conc. (μg) | S. typhimurium | S. enteritidis | Total |
| Ampicillin | AMP | 10 μg | 3 (6.7%) | − | 3 (4.8%) |
| Sulphonamides | SSS | 300 μg | 8 (17.8%) | 5 (27.8%) | 13 (20.6%) |
| Cephalothin | KF | 30 μg | 11 (24.4%) | 3 (16.7%) | 14 (22.2%) |
| Tetracycline | TE | 30 μg | 23 (51.1%) | 6 (33.3%) | 29 (46.0%) |
| Ciprofloxacin | NA | 30 μg | − | 2 (11.1%) | 2 (3.2%) |
| Nalidixic acid | C | 30 μg | 5 (11.1%) | 4 (22.2%) | 9 (14.3%) |
| Chloramphenicol | CA | 10 μg | − | − | − |
| Gentamicin | ENR | 5 μg | 6 (13.3%) | 5 (27.8%) | 11 (17.5%) |
| Enrofloxacin | RD | 5 μg | 26 (57.8%) | 13 (72.2%) | 39 (61.9%) |
| Rifampicin | S | 10 μg | 6 (13.3%) | − | 6 (9.5%) |
| Streptomycin | CIP | 5μg | 14 (31.1%) | 7 (38.9%) | 21 (33.3%) |
| Serovars | Sample ID | Accession Number | Antimicrobial-Resistant Genes Pattern | Integrase | ||
|---|---|---|---|---|---|---|
| IntI1 | IntI2 | IntI3 | ||||
| S. Typhimurium | R 1 | MH352147 | mcr-4, blaCTX-M-2, blaCTX-M-9 | + | − | − |
| R 3 | MH352149 | mcr-4, blaCTX-M, blaCTX-M-2, blaCTX-M-9, blaCTX-M-15 | + | + | − | |
| R 6 | MH352152 | mcr-4, blaCTX-M, blaCTX-M-2, blaCTX-M-9 | + | − | − | |
| R 7 | MH352153 | mcr-4, blaCTX-M-2, blaCTX-M-9 | + | + | − | |
| R 8 | MH352154 | mcr-4, blaCTX-M, blaCTX-M-2, blaCTX-M-9 | + | − | − | |
| R 9 | MH352155 | mcr-4, blaCTX-M-2,blaCTX-M-9 | + | − | − | |
| R 10 | MH352156 | mcr-4, blaCTX-M-2, blaCTX-M-9, blaTEM | + | + | − | |
| R 11 | MH352157 | blaCTX-M-2, blaCTX-M-9 | + | + | − | |
| R 12 | MH352158 | blaCTX-M-2, blaCTX-M-9 | + | + | − | |
| R 22 | MH352168 | mcr-4, blaCTX-M, blaCTX-M-9 | + | + | − | |
| R 25 | MH352171 | mcr-4, blaCTX-M-2, blaCTX-M-9 | + | + | − | |
| R 28 | MH352174 | mcr-4, blaCTX-M-2, blaCTX-M-9, blaTEM | + | + | − | |
| R 29 | MH352175 | mcr-4, blaCTX-M-9 | + | + | − | |
| R 30 | MH352176 | mcr-4, blaCTX-M-9 | + | + | − | |
| R 36 | MH352182 | mcr-4, blaCTX-M, blaCTX-M-9, blaCTX-M-15 | + | + | − | |
| R 37 | MH352183 | mcr-4, blaCTX-M-9 | + | − | − | |
| R 39 | MH352185 | mcr-4, blaCTX-M-2, blaCTX-M-9 | + | − | − | |
| R 43 | MH352189 | mcr-4, blaCTX-M-2, blaCTX-M-9, blaTEM | + | + | − | |
| R 44 | MH352190 | mcr-4, blaCTX-M-9 | + | − | − | |
| R 45 | MH352191 | mcr-4, blaCTX-M-9 | + | + | − | |
| R 46 | MH352192 | mcr-4, blaCTX-M-9 | + | − | − | |
| R 48 | MH352194 | blaCTX-M-2, blaCTX-M-9 | + | − | − | |
| R 49 | MH352195 | blaCTX-M-2, blaCTX-M-9, blaTEM | + | + | − | |
| R 51 | MH352197 | blaCTX-M-2, blaCTX-M-9 | + | − | − | |
| R 52 | MH352198 | blaCTX-M-2, blaCTX-M-9 | + | − | − | |
| R 53 | MH352199 | mcr-4, blaCTX-M, blaCTX-M-2, blaCTX-M-15 | + | + | − | |
| R 54 | MH352200 | mcr-4, blaCTX-M-2, blaCTX-M-9 | + | − | − | |
| R 56 | MH352202 | blaCTX-M-2, blaCTX-M-9 | + | − | − | |
| R 60 | MH352206 | blaCTX-M-9 | + | + | − | |
| R 65 | MH352211 | mcr-4, blaCTX-M-9 | + | + | − | |
| R 67 | MH352213 | mcr-4, blaCTX-M-2, blaCTX-M-9, blaTEM | + | − | − | |
| C 6 | MH356675 | mcr-4 | − | − | − | |
| C 7 | MH356676 | mcr-4, blaCTX-M | + | + | − | |
| C 11 | MH356680 | mcr-4, blaCTX-M, blaCTX-M-15 | − | − | − | |
| C 12 | MH356681 | − | − | + | − | |
| C 23 | MH356692 | − | + | − | − | |
| C 26 | MH356695 | blaCTX-M-15 | − | − | − | |
| C 28 | MH356697 | blaCTX-M, blaCTX-M-15 | − | + | − | |
| C 30 | MH356699 | blaCTX-M-15 | − | − | − | |
| C 32 | MH356701 | − | − | − | − | |
| C 34 | MH356703 | − | − | − | − | |
| C 34 | MH356704 | − | − | − | − | |
| C 36 | MH356705 | − | − | + | − | |
| C 37 | MH356706 | − | − | − | − | |
| C 41 | MH356710 | blaCTX-M-15 | + | − | − | |
| C 42 | MH356711 | − | − | − | − | |
| C 43 | MH356712 | blaCTX-M | − | − | − | |
| C 44 | MH356713 | − | − | − | − | |
| C 45 | MH356714 | − | + | + | − | |
| C 46 | MH356715 | blaCTX-M-15 | − | − | − | |
| S. Enteritidis | C 1 | MH356670 | blaCTX-M-15 | + | − | − |
| C 8 | MH356677 | mcr-4 | + | − | − | |
| C 20 | MH356689 | − | + | + | − | |
| C 22 | MH356691 | − | + | − | − | |
| C 29 | MH356698 | mcr-4 | + | − | − | |
| C 40 | MH356709 | mcr-4, blaCTX-M-1 | + | − | − | |
| R 2 | MH352148 | mcr-4, blaCTX-M-1 | + | − | − | |
| R 4 | MH352150 | blaCTX-M-1 | + | − | − | |
| R 7 | MH352153 | blaCTX-M-1 | + | + | − | |
| R 27 | MH352173 | blaCTX-M | + | − | − | |
| R 37 | MH352183 | blaCTX-M, blaCTX-M-15 | + | − | − | |
| R 50 | MH352196 | blaCTX-M, blaSHV | + | + | − | |
| R 57 | MH352203 | mcr-4, blaCTX-M, blaSHV | + | + | − | |
| R 58 | MH352204 | blaCTX-M, blaCTX-M-1, blaCTX-M-15, blaSHV | + | − | − | |
| R 59 | MH352205 | blaCTX-M, blaCTX-M-15 | + | − | − | |
| R 62 | MH352208 | blaCTX-M, blaCTX-M-1, blaCTX-M-15,blaSHV | + | − | − | |
| R 64 | MH352210 | blaCTX-M-1, blaCTX-M-15 | + | + | − | |
| R 68 | MH352214 | blaCTX-M, blaCTX-M-1 | + | + | − | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramatla, T.; Mileng, K.; Ndou, R.; Mphuti, N.; Syakalima, M.; Lekota, K.E.; Thekisoe, O.M.M. Molecular Detection of Integrons, Colistin and β-lactamase Resistant Genes in Salmonella enterica Serovars Enteritidis and Typhimurium Isolated from Chickens and Rats Inhabiting Poultry Farms. Microorganisms 2022, 10, 313. https://doi.org/10.3390/microorganisms10020313
Ramatla T, Mileng K, Ndou R, Mphuti N, Syakalima M, Lekota KE, Thekisoe OMM. Molecular Detection of Integrons, Colistin and β-lactamase Resistant Genes in Salmonella enterica Serovars Enteritidis and Typhimurium Isolated from Chickens and Rats Inhabiting Poultry Farms. Microorganisms. 2022; 10(2):313. https://doi.org/10.3390/microorganisms10020313
Chicago/Turabian StyleRamatla, Tsepo, Kealeboga Mileng, Rendani Ndou, Nthabiseng Mphuti, Michelo Syakalima, Kgaugelo E. Lekota, and Oriel M.M. Thekisoe. 2022. "Molecular Detection of Integrons, Colistin and β-lactamase Resistant Genes in Salmonella enterica Serovars Enteritidis and Typhimurium Isolated from Chickens and Rats Inhabiting Poultry Farms" Microorganisms 10, no. 2: 313. https://doi.org/10.3390/microorganisms10020313
APA StyleRamatla, T., Mileng, K., Ndou, R., Mphuti, N., Syakalima, M., Lekota, K. E., & Thekisoe, O. M. M. (2022). Molecular Detection of Integrons, Colistin and β-lactamase Resistant Genes in Salmonella enterica Serovars Enteritidis and Typhimurium Isolated from Chickens and Rats Inhabiting Poultry Farms. Microorganisms, 10(2), 313. https://doi.org/10.3390/microorganisms10020313