
Prevalence and Profiles of Antibiotic Resistance Genes mph(A) and qnrB in Extended-Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli Isolated from Dairy Calf Feces

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Calf Management and Treatment Protocols
2.3. Microbiological Processing for ESBL-Producing E. coli
2.4. Enterobacteriaceae and E. coli Confirmation
2.5. E. coli Enumeration by Colony Forming Units per Gram Feces (CFU/g)
2.6. DNA Extraction and Endpoint PCR for blaCTX-M, qnrB, and mph(A) Genes
2.7. Missing or Excluded Samples and Isolates
2.8. DNA Extraction and E. coli Whole-Genome Sequencing (WGS)
2.9. Bioinformatics Analysis
2.10. Data Analysis
2.11. Ethics Statement
3. Results
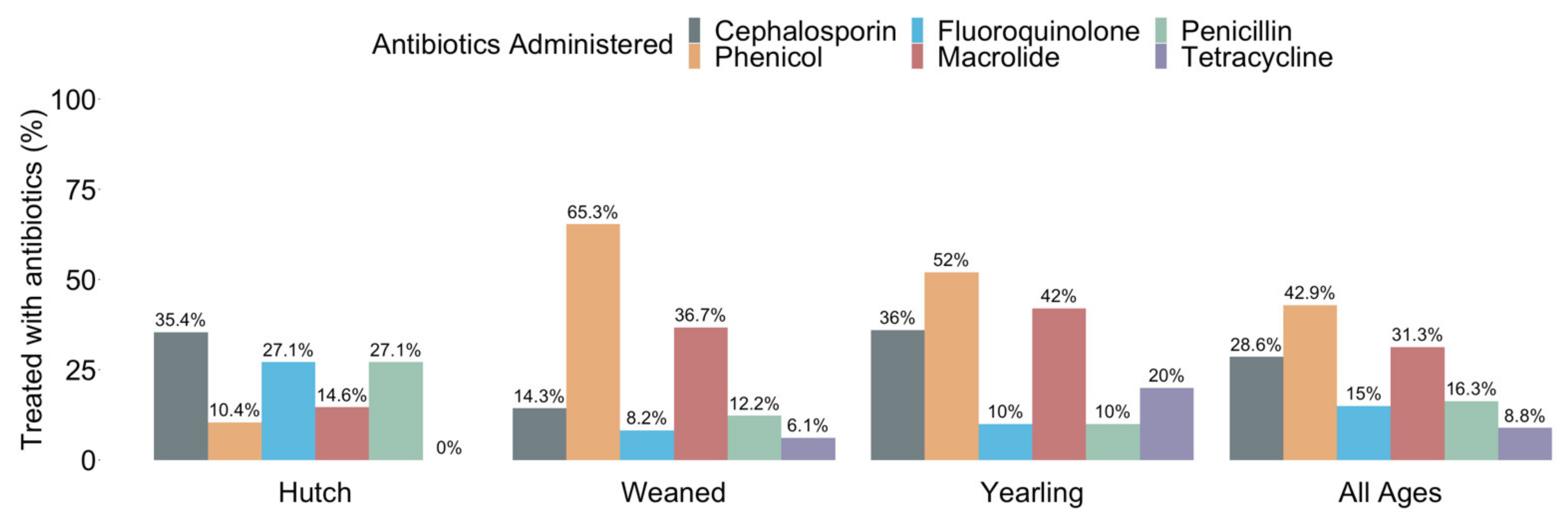
3.1. ESBL Prevalence and Antibiotic Use in Hutch, Weaned, and Yearling Calves
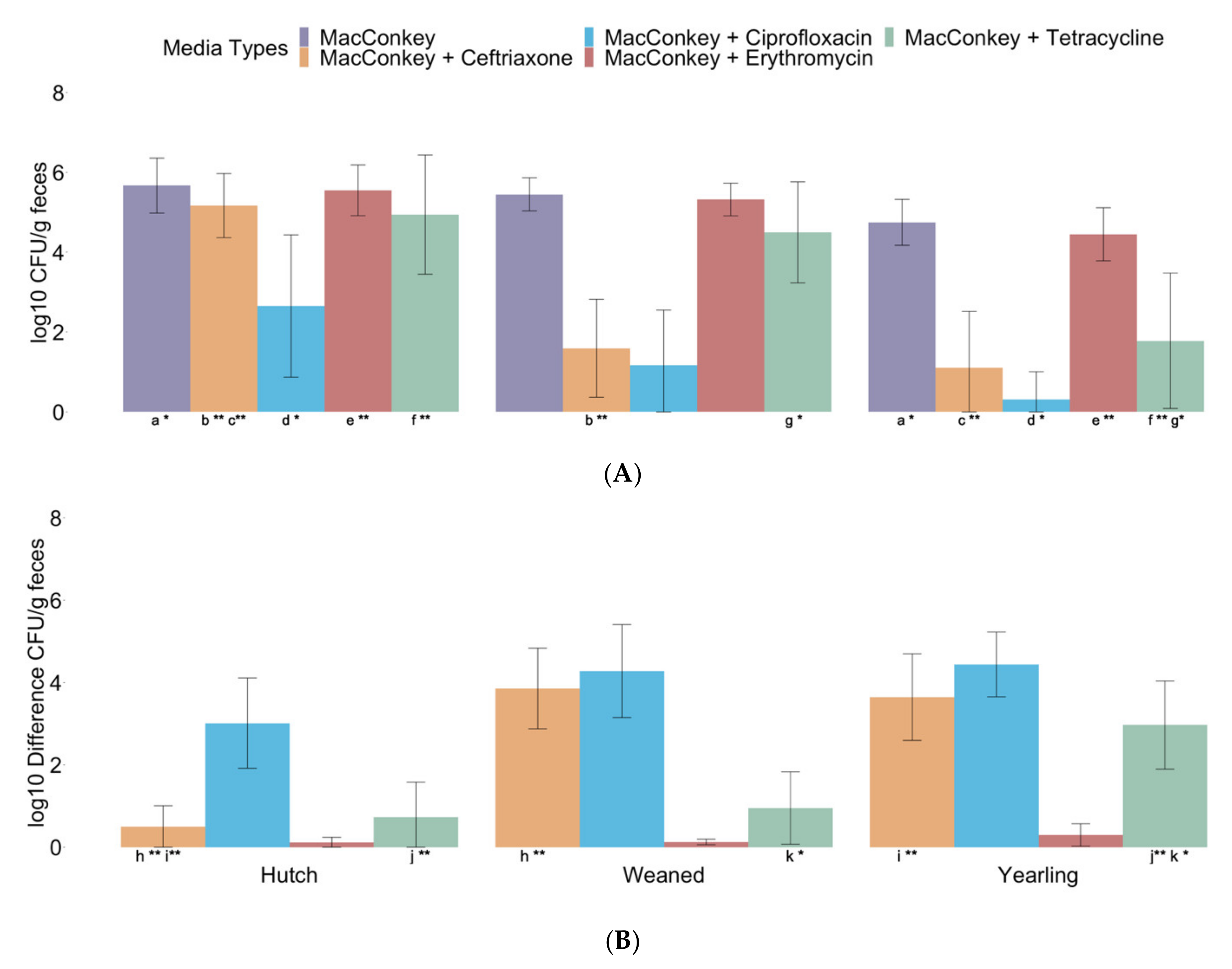
3.2. Enumeration of Antibiotic-Resistant E. coli in Hutch, Weaned, and Yearling Calves
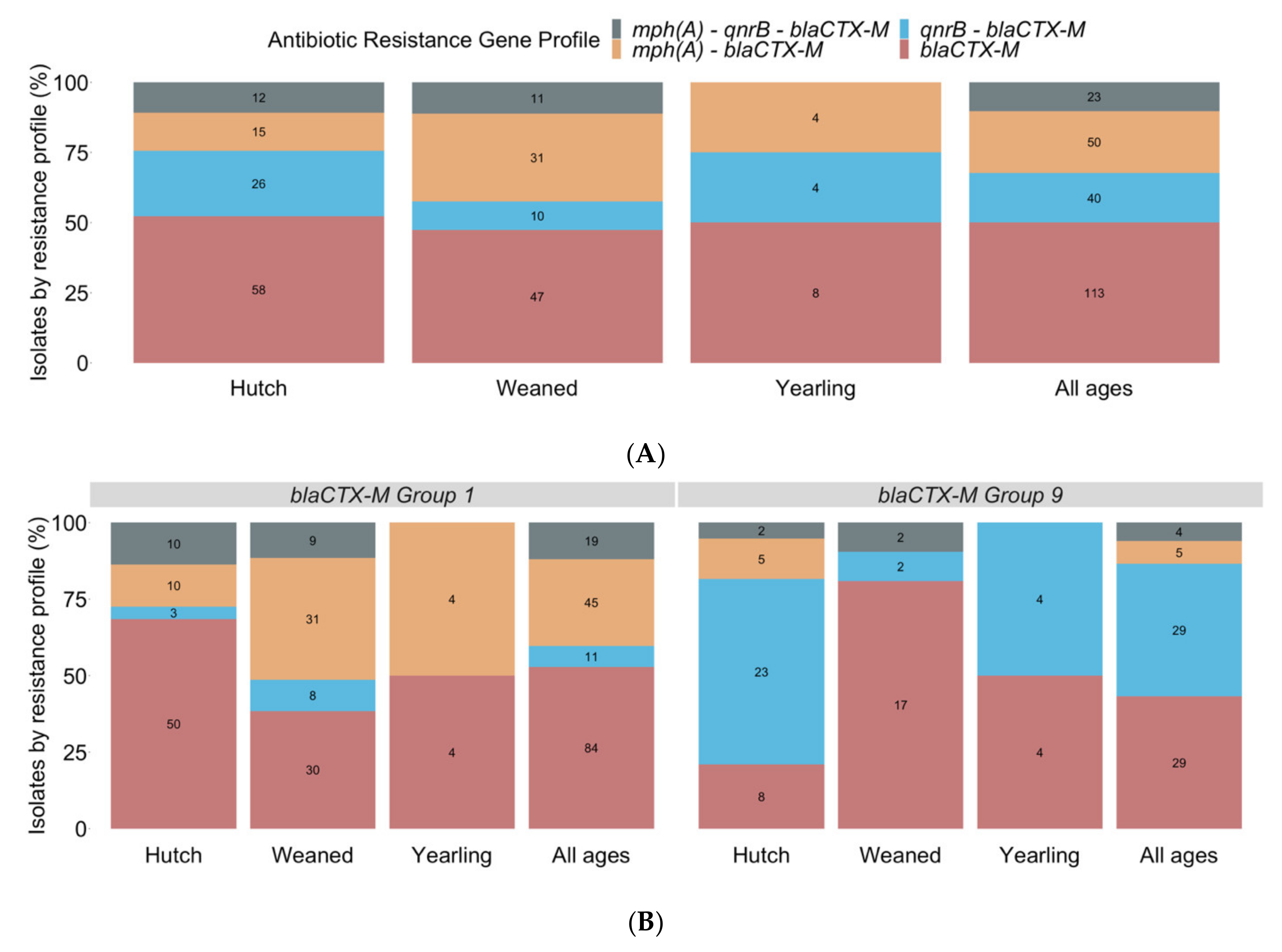
3.3. Resistance Profiles Identified by PCR for blaCTX-M, qnrB, and mph(A)
3.4. WGS of ESBL-Producing E. coli Isolates
4. Discussion
4.1. Age-Related Prevalence of ESBL-Producing E. coli
4.2. Antibiotic Use and Age-Related Growth of E. coli and Antibiotic-Resistant E. coli
4.3. Genotypic Resistance Profiles Identified by PCR for blaCTX-M, qnrB, and mph(A)
4.4. Diversity of ESBL-Producing E. coli
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019.
- WHO. Critically Important Antimicrobials for Human Medicine, 5th ed.; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Hammerum, A.M.; Heuer, O.E. Human health hazards from antimicrobial-resistant Escherichia coli of animal origin. Clin. Infect. Dis. 2009, 48, 916–921. [Google Scholar] [CrossRef] [Green Version]
- Tollefson, L.; Karp, B.E. Human health impact from antimicrobial use in food animals. Méd. Mal. Infect. 2004, 34, 514–521. [Google Scholar] [CrossRef]
- United States Food and Drug Administration. Animal Medicinal Drug Use Clarification Act of 1994 (AMDUCA). Available online: https://www.fda.gov/animal-veterinary/guidance-regulations/animal-medicinal-drug-use-clarification-act-1994-amduca (accessed on 20 January 2022).
- Catry, B.; Laevens, H.; Devriese, L.A.; Opsomer, G.; de Kruif, A. Antimicrobial resistance in livestock. J. Vet. Pharmacol. Ther. 2003, 26, 81–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, T.L.; Callaway, T.R.; Norman, K.N.; Scott, H.M.; Loneragan, G.H.; Ison, S.A.; Beier, R.C.; Harhay, D.M.; Norby, B.; Nisbet, D.J. Transferability of antimicrobial resistance from multidrug-resistant Escherichia coli isolated from cattle in the USA to E. coli and Salmonella Newport recipients. J. Glob. Antimicrob. Resist. 2017, 11, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Pradhan, A.K.; Karns, J.S.; Hovingh, E.; Wolfgang, D.R.; Vinyard, B.T.; Kim, S.W.; Salaheen, S.; Haley, B.J.; Van Kessel, J.A.S. Age-Associated Distribution of Antimicrobial-Resistant Salmonella enterica and Escherichia coli Isolated from Dairy Herds in Pennsylvania, 2013–2015. Foodborne Pathog. Dis. 2019, 16, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Tacao, M.; Moura, A.; Correia, A.; Henriques, I. Co-resistance to different classes of antibiotics among ESBL-producers from aquatic systems. Water Res. 2014, 48, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.A.; Ossa-Trujillo, C.; Vinasco, J.; Jordan, E.R.; Garcia Buitrago, J.A.; Hagevoort, R.; Norman, K.N.; Lawhon, S.D.; Pineiro, J.M.; Levent, G.; et al. Use of critically important antimicrobial classes early in life may adversely impact bacterial resistance profiles during adult years: Potential co-selection for plasmid-borne fluoroquinolone and macrolide resistance via extended-spectrum beta-lactam use in dairy cattle. Lett. Appl. Microbiol. 2021, 72, 220–224. [Google Scholar] [CrossRef]
- Waade, J.; Seibt, U.; Honscha, W.; Rachidi, F.; Starke, A.; Speck, S.; Truyen, U. Multidrug-resistant enterobacteria in newborn dairy calves in Germany. PLoS ONE 2021, 16, e0248291. [Google Scholar] [CrossRef]
- Singer, R.S.; Patterson, S.K.; Wallace, R.L. Effects of therapeutic ceftiofur administration to dairy cattle on Escherichia coli dynamics in the intestinal tract. Appl. Environ. Microbiol. 2008, 74, 6956–6962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, E.A.; Jordan, E.R.; Garcia, J.A.; Hagevoort, G.R.; Norman, K.N.; Lawhon, S.D.; Pineiro, J.M.; Scott, H.M. Effects of two-dose ceftiofur treatment for metritis on the temporal dynamics of antimicrobial resistance among fecal Escherichia coli in Holstein-Friesian dairy cows. PLoS ONE 2019, 14, e0220068. [Google Scholar] [CrossRef] [Green Version]
- Wellman, N.G.; O’Connor, A.M. Meta-analysis of treatment of cattle with bovine respiratory disease with tulathromycin. J. Vet. Pharmacol. Ther. 2007, 30, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, R. Mechanisms of Resistance to Macrolides and Lincosamides: Nature of the Resistance Elements and Their Clinical Implications. Clin. Infect. Dis. 2002, 34, 482–492. [Google Scholar] [CrossRef] [Green Version]
- Phuc Nguyen, M.C.; Woerther, P.L.; Bouvet, M.; Andremont, A.; Leclercq, R.; Canu, A. Escherichia coli as reservoir for macrolide resistance genes. Emerg. Infect. Dis. 2009, 15, 1648–1650. [Google Scholar] [CrossRef]
- Poirel, L.; Cattoir, V.; Nordmann, P. Plasmid-Mediated Quinolone Resistance; Interactions between Human, Animal, and Environmental Ecologies. Front. Microbiol. 2012, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Tran, J.H.; Jacoby, G.A. Mechanism of plasmid-mediated quinolone resistance. Proc. Natl. Acad. Sci. USA 2002, 99, 5638–5642. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.A.; Heaton, K.; Poisson, S.; Sischo, W.M. Dairy Calf Housing and Environment: The Science Behind Housing and On-Farm Assessments Dairy Calf Housing and Environment: The Science Behind Housing and On-Farm Assessments; Washington State University Extension: Pullman, WA, USA, 2012. [Google Scholar]
- Okello, E.; Williams, D.R.; ElAshmawy, W.R.; Adams, J.; Pereira, R.V.; Lehenbauer, T.W.; Aly, S.S. Survey on Antimicrobial Drug Use Practices in California Preweaned Dairy Calves. Front. Vet. Sci. 2021, 8, 636670. [Google Scholar] [CrossRef]
- Enroflox 100 Package Insert. 2017. Available online: https://www.norbrook.com/media/2873/package-insert-enroflox-100.pdf (accessed on 12 November 2021).
- Draxxin Cattle Prescribing Information. 2014. Available online: https://www.zoetisus.com/draxxin-25/PDF/Draxxin_25_Full_PI_R2.pdf (accessed on 12 November 2021).
- Excede Cattle Marketing Package Insert. 2020. Available online: https://www.zoetisus.com/products/pages/excede_dairy/documents/excede-cattle-marketing-package-insert.pdf (accessed on 12 November 2021).
- Liu, J.; Zhao, Z.; Orfe, L.; Subbiah, M.; Call, D.R. Soil-borne reservoirs of antibiotic-resistant bacteria are established following therapeutic treatment of dairy calves. Environ. Microbiol. 2016, 18, 557–564. [Google Scholar] [CrossRef]
- Pereira, R.V.; Siler, J.D.; Ng, J.C.; Davis, M.A.; Grohn, Y.T.; Warnick, L.D. Effect of on-farm use of antimicrobial drugs on resistance in fecal Escherichia coli of preweaned dairy calves. J. Dairy Sci. 2014, 97, 7644–7654. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testings: 25th Informational Supplement; Clinical Laboratory Standards Institute: Malvern, PA, USA, 2016. [Google Scholar]
- CDC. CDC National Antimicrobial Resistance Monitoring System for Enteric Bacteria: Antibiotics Tested by NARMS. Available online: https://www.cdc.gov/narms/antibiotics-tested.html (accessed on 27 December 2021).
- Sutcliffe, J.; Grebe, T.; Tait-Kamradt, A.; Wondrack, L. Detection of erythromycin-resistant determinants by PCR. Antimicrob. Agents Chemother. 1996, 40, 2562–2566. [Google Scholar] [CrossRef] [Green Version]
- Marti, E.; Jofre, J.; Balcazar, J.L. Prevalence of antibiotic resistance genes and bacterial community composition in a river influenced by a wastewater treatment plant. PLoS ONE 2013, 8, e78906. [Google Scholar] [CrossRef]
- Xu, L.; Ensor, V.; Gossain, S.; Nye, K.; Hawkey, P. Rapid and simple detection of blaCTX-M genes by multiplex PCR assay. J. Med. Microbiol. 2005, 54, 1183–1187. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewels, P.; Magnusson, M.; Lundin, S.; Kaller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Joensen, K.G.; Tetzschner, A.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and Easy In Silico Serotyping of Escherichia coli Isolates by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.J.; Taylor, B.; Keane, J.A. Multilocus sequence typing by blast from de novo assemblies against PubMLST. J. Open Source Softw. 2016, 8, 118. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Allesoe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- StataCorp. Stata Statistical Software: Release 17; StataCorp LLC: College Station, TX, USA, 2021. [Google Scholar]
- Hope, R.M. Rmisc: Rmisc: Ryan Miscellaneous; R Package Version 1.5; The Comprehensive R Archive Network: 2013. Available online: https://CRAN.R-project.org/package=Rmisc (accessed on 12 November 2021).
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots; R Package Version 0.3.0; The Comprehensive R Archive Network: 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 12 November 2021).
- Müller, K.; Wickham, H. tibble: Simple Data Frames; R Package Version 3.0.1; The Comprehensive R Archive Network: 2020. Available online: https://CRAN.R-project.org/package=tibble (accessed on 12 November 2021).
- RStudio Team. RStudio: Integrated Development for R; Version 2021.9.0.351; RStudio, PBC: Boston, MA, USA, 2021. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 12 November 2021).
- Wickham, H.; Bryan, J. readxl: Read. Excel Files, R Package Version 1.3.1; The Comprehensive R Archive Network: Vienna, Austria, 2019. Available online: https://CRAN.R-project.org/package=readxl(accessed on 12 November 2021).
- Xiao, N. ggsci: Scientific Journal and Sci-Fi Themed Color. Palettes for ‘ggplot2’, R Package Version 2.9; The Comprehensive R Archive Network: Vienna, Austria, 2018. Available online: https://CRAN.R-project.org/package=ggsci(accessed on 12 November 2021).
- Castanheira, M.; Simner, P.J.; Bradford, P.A. Extended-spectrum beta-lactamases: An update on their characteristics, epidemiology and detection. JAC Antimicrob. Resist. 2021, 3, dlab092. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Sharara, S.L.; Pana, Z.D.; Amoah, J.; Fisher, S.L.; Tekle, T.; Doi, Y.; Simner, P.J. Molecular Epidemiology of Ceftriaxone Non-Susceptible Enterobacterales Isolates in an Academic Medical Center in the United States. Open Forum Infect. Dis. 2019, 6, ofz353. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.D.; Johnson, S.J.; Cerniglia, C.E.; Erickson, B.D. Bovine intestinal bacteria inactivate and degrade ceftiofur and ceftriaxone with multiple beta-lactamases. Antimicrob. Agents Chemother. 2011, 55, 4990–4998. [Google Scholar] [CrossRef] [Green Version]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [Green Version]
- García Ovando, H.; Gorla, N.; Luders, C.; Poloni, G.; Errecalde, C.; Prieto, G.; Puelles, I. Comparative pharmacokinetics of enrofloxacin and ciprofloxacin in chickens. J. Vet. Pharmacol. Ther. 1999, 22, 209–212. [Google Scholar] [CrossRef]
- Constable, P.D. Antimicrobial Use in the Treatment of Calf Diarrhea. J. Vet. Intern. Med. 2004, 18, 8–17. [Google Scholar] [CrossRef]
- Chittum, H.S.; Champney, W.S. Erythromycin Inhibits the Assembly of the Large Ribosomal Subunit in Growing Escherichia coli Cells. Curr. Microbiol. 1995, 30, 273–279. [Google Scholar] [CrossRef]
- Edgar, R.; Bibi, E. MdfA, an Escherichia coli Multidrug Resistance Protein with an Extraordinarily Broad Spectrum of Drug Recognition. J. Bacteriol. 1997, 179, 2274–2280. [Google Scholar] [CrossRef] [Green Version]
- Zuprevo Product Information Insert. 2017. Available online: https://vet-advantage.com/wp-content/uploads/sites/5/2019/10/Merck-Zuprevo-SB.pdf (accessed on 27 December 2021).
- Tylan200 Injection (Tylosin Injection) Package Insert. Available online: https://www.elancolabels.com/us/tylan-200-injection-500ml (accessed on 27 December 2021).
- Noguchi, N.; Takada, K.; Katayama, J.; Emura, A.; Sasatsu, M. Regulation of Transcription of the mph(A) Gene for Macrolide 2?-Phosphotransferase I in Escherichia coli: Characterization of the Regulatory Gene mphR(A). J. Bacteriol. 2000, 182, 5052–5058. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, R. Growing group of extended-spectrum beta-lactamases: The CTX-M enzymes. Antimicrob. Agents Chemother. 2004, 48, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossolini, G.M.; D’Andrea, M.M.; Mugnaioli, C. The spread of CTX-M-type extended-spectrum b-lactamases. Eur. Soc. Clin. Microbiol. Infect. Dis. 2008, 14, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afema, J.A.; Ahmed, S.; Besser, T.E.; Jones, L.P.; Sischo, W.M.; Davis, M.A. Molecular Epidemiology of Dairy Cattle-Associated Escherichia coli Carrying bla CTX-M Genes in Washington State. Appl. Environ. Microbiol. 2018, 84, e02430-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, T.; Ur Rahman, S.; Zhang, L.; Shahid, M.; Han, D.; Gao, J.; Zhang, S.; Ruegg, P.L.; Saddique, U.; Han, B. Characteristics and genetic diversity of multi-drug resistant extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli isolated from bovine mastitis. Appl. Environ. Microbiol. 2018, 8, 90144–90163. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Fratamico, P.M.; Feng, P. Pathogenic Escherichia coli. In Compendium of Methods for the Microbiological Examination of Foods; American Public Health Association: Washington, DC, USA, 2013. [Google Scholar]
- Ison, S.A.; Delannoy, S.; Bugarel, M.; Nightingale, K.K.; Webb, H.E.; Renter, D.G.; Nagaraja, T.G.; Loneragan, G.H.; Fach, P. Genetic Diversity and Pathogenic Potential of Attaching and Effacing Escherichia coli O26:H11 Strains Recovered from Bovine Feces in the United States. Appl. Environ. Microbiol. 2015, 81, 3671–3678. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence | Product | Annealing | Source |
|---|---|---|---|---|
| mph(A) | F 5′-AAC TGT ACG CAC TTG C-3′ R 5′-GGT ACT CTT CGT TAC C-3′ | 837 bp | 52 °C | [28] |
| qnrB (1–6) | F 5′ -GGM ATH GAA ATT CGC CAC TG-3′ R 5′ -TTY GCB GYY CGC CAG TCG AA-3′ | 264 bp | 54 °C | [29] |
| blaCTX-M-Group-1 | F 5′-GCG TGA TAC CAC TTC ACC TC-3′ R 5′-TGA AGT AAG TGA CCA GAA TC-3′ | 260 bp | 55 °C | [30] |
| blaCTX-M-Group-9 | F 5′-ATC AAG CCT GCC GAT CTG GTT A-3′ R 5′-GTA AGC TGA CGC AAC GTC TGC-3′ | 293 bp | 55 °C | [31] |
| qnrB19 | F 5′-CAC ATT GCG TGA CCA ATT-3′ R 5′-GAT GCC TGG TAG CTG TCT AAC-3′ | 90 bp | 60 °C | This study |
| Item | Hutch | Weaned | Yearling | Total |
|---|---|---|---|---|
| No. of Calves | 48 | 49 | 50 | 147 |
| Mean Age (Range) * | 1.8 (1.6–3.0) | 5.0 (3.5–6.9) | 12.9 (12.0–16.5) | 6.6 (1.6–16.5) |
| No. Confirmed ESBL | 37 | 30 | 4 | 71 |
| % ESBL (95%CI) | 77.1% (62.7–88.0%) | 61.2% (46.2–74.8%) | 8.0% (2.2–19.2%) | 48.3% (40.0–56.7%) |
| No. ESBL Isolates ** | 111 | 99 | 16 | 226 |
| Isolate | Age Group | Resistance Genes by Antibiotic Class | Serotype | Sequence Type (ST) | Plasmids | PCR Profile (blaCTX-M Group) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Beta-Lactam | Aminoglycoside | Trimethoprim | Sulfonamide | MLS | Phenicol | Quinolone | Tetracycline | Point Mutation | ||||||
| 18-H-6-Ecoli-Feb2020-2 | weaned | blaCTX-M-1 blaTEM-1A | aph(3″)-Ib aph(6)-Id | sul2 | mdf(A) mph(A) | floR | qnrB19 | tet(A) | O40:H4 | 226 | IncY, IncR, Col440I | mph(A), qnrB, blaCTX-M (1) | ||
| 19-H-6-Ecoli-Feb2020-1 | weaned | blaCTX-M-32 | dfrA1 | sul1 | mdf(A) mph(A) | floR | tet(A) | O134:H38 | 154 | IncFIB, IncR | mph(A), blaCTX-M (1) | |||
| 21-B-6-Ecoli-Feb2020-2 | yearling | blaCTX-M-102 * blaCTX-M-174 * | aph(6)-Id | sul2 | mdf(A) | qnrB19 | O26:H11 | 29 | IncFIA, IncFIB, Col(MG828), Col440I, Col156 | qnrB, blaCTX-M (9) | ||||
| 34-H-6-Ecoli-Feb2020-1 | weaned | blaCTX-M-15 blaTEM-1B | aph(6)-Id aph(3″)-Ib | sul2 | mdf(A) | qnrB19 qnrS1 | O4:H11 | 641 | IncFIB, IncY, Col440I | qnrB, blaCTX-M (1) | ||||
| 5-B-3-Ecoli-Feb2020-1 | yearling | blaCTX-M-27 | aph(3″)-Ib aph(6)-Id | sul2 | mdf(A) | floR | tet(A) | O8:H30 | 58 | IncFIA, IncFIB, IncFII | blaCTX-M (9) | |||
| 5-H-6-Ecoli-Feb2020-3 | weaned | blaCTX-M-27 | mdf(A) | tet(A) | O70:H2 | 10 | IncFIB, IncFII | blaCTX-M (1) | ||||||
| 13-W-3-Ecoli-Feb2020-3 | hutch | blaCTX-M-65 blaCARB-2 blaTEM-1A | aph(3′)-IIa aph(6)-Id aph(3″)-Ib aph(3′)-Ia | sul1 sul2 | mdf(A) mph(A) | qnrA1 | gyrA p.S83L ** | O100:H30 | 993 | IncI1α, IncX1, IncHI2, IncHI2A | mph(A), blaCTX-M (9) | |||
| 47-W-3-Ecoli-Feb2020-2 | hutch | blaCTX-M-65 | dfrA1 | sul1 sul2 | mdf(A) mph(A) * | floR | qnrB19 | O103:H2 | 1967 | IncHI2A, IncHI2, IncFII, Col440I, Col156 | mph(A), qnrB, blaCTX-M (9) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carey, A.M.; Capik, S.F.; Giebel, S.; Nickodem, C.; Piñeiro, J.M.; Scott, H.M.; Vinasco, J.; Norman, K.N. Prevalence and Profiles of Antibiotic Resistance Genes mph(A) and qnrB in Extended-Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli Isolated from Dairy Calf Feces. Microorganisms 2022, 10, 411. https://doi.org/10.3390/microorganisms10020411
Carey AM, Capik SF, Giebel S, Nickodem C, Piñeiro JM, Scott HM, Vinasco J, Norman KN. Prevalence and Profiles of Antibiotic Resistance Genes mph(A) and qnrB in Extended-Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli Isolated from Dairy Calf Feces. Microorganisms. 2022; 10(2):411. https://doi.org/10.3390/microorganisms10020411
Chicago/Turabian StyleCarey, Alexis M., Sarah F. Capik, Sarah Giebel, Colette Nickodem, Juan M. Piñeiro, Harvey Morgan Scott, Javier Vinasco, and Keri N. Norman. 2022. "Prevalence and Profiles of Antibiotic Resistance Genes mph(A) and qnrB in Extended-Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli Isolated from Dairy Calf Feces" Microorganisms 10, no. 2: 411. https://doi.org/10.3390/microorganisms10020411
APA StyleCarey, A. M., Capik, S. F., Giebel, S., Nickodem, C., Piñeiro, J. M., Scott, H. M., Vinasco, J., & Norman, K. N. (2022). Prevalence and Profiles of Antibiotic Resistance Genes mph(A) and qnrB in Extended-Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli Isolated from Dairy Calf Feces. Microorganisms, 10(2), 411. https://doi.org/10.3390/microorganisms10020411