
Squalamine and Its Aminosterol Derivatives: Overview of Biological Effects and Mechanisms of Action of Compounds with Multiple Therapeutic Applications

 , ,
, , 
Abstract
:1. Introduction
2. Structure of Squalamine and Its Derivatives
3. Aminosterols and Their Mode of Action
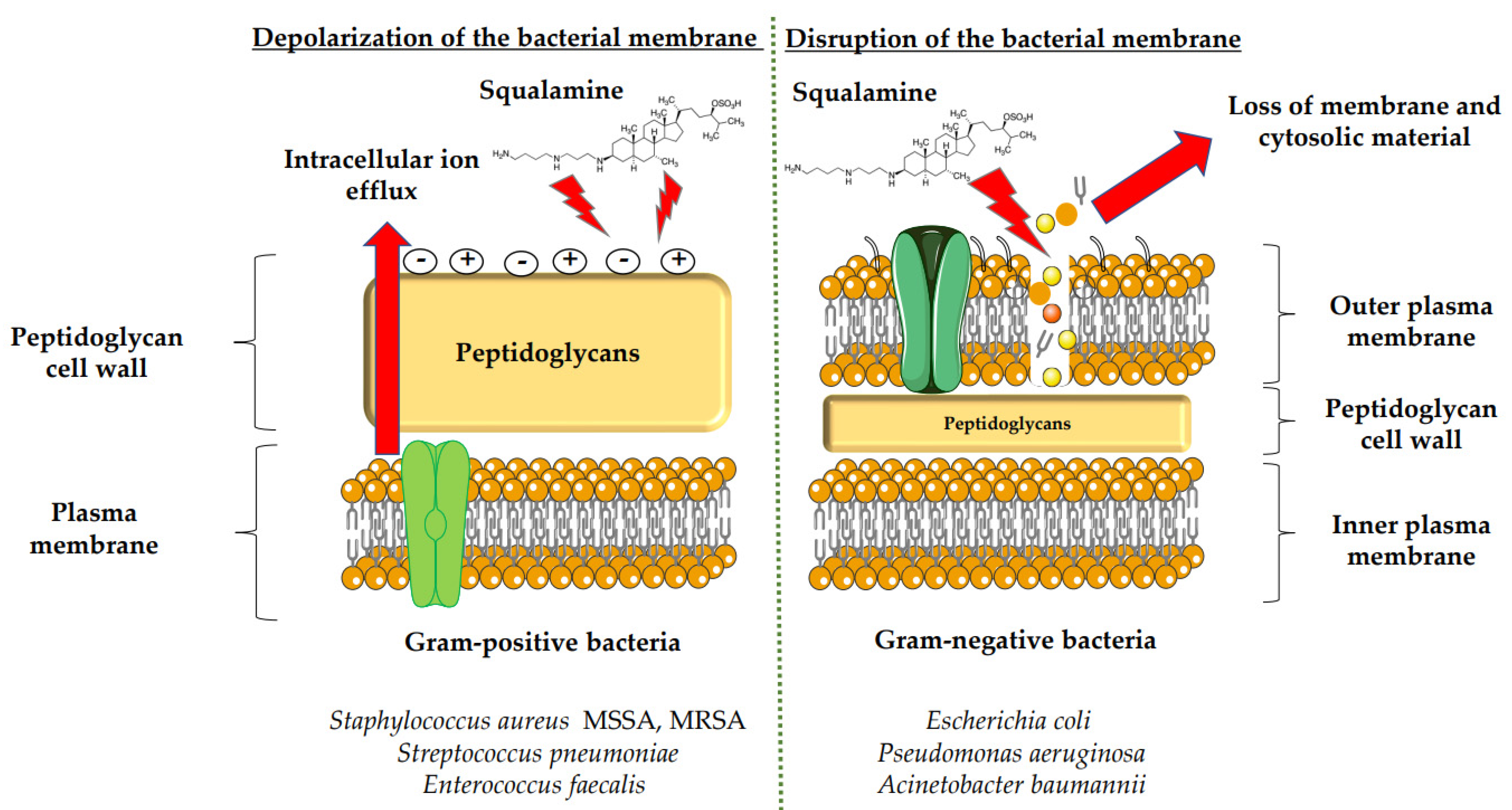
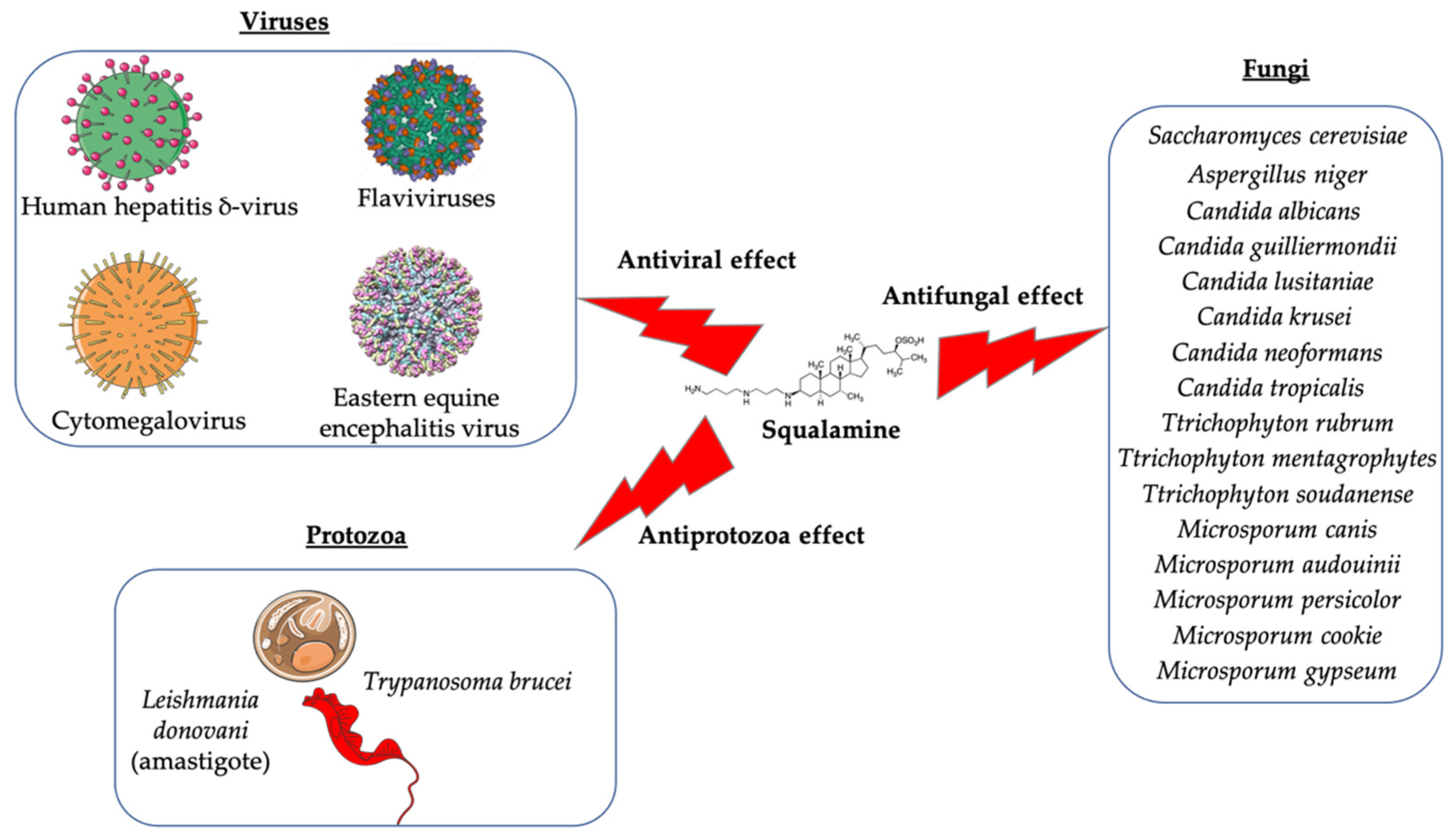
3.1. Antimicrobial Activity of Squalamine and Its Derivatives
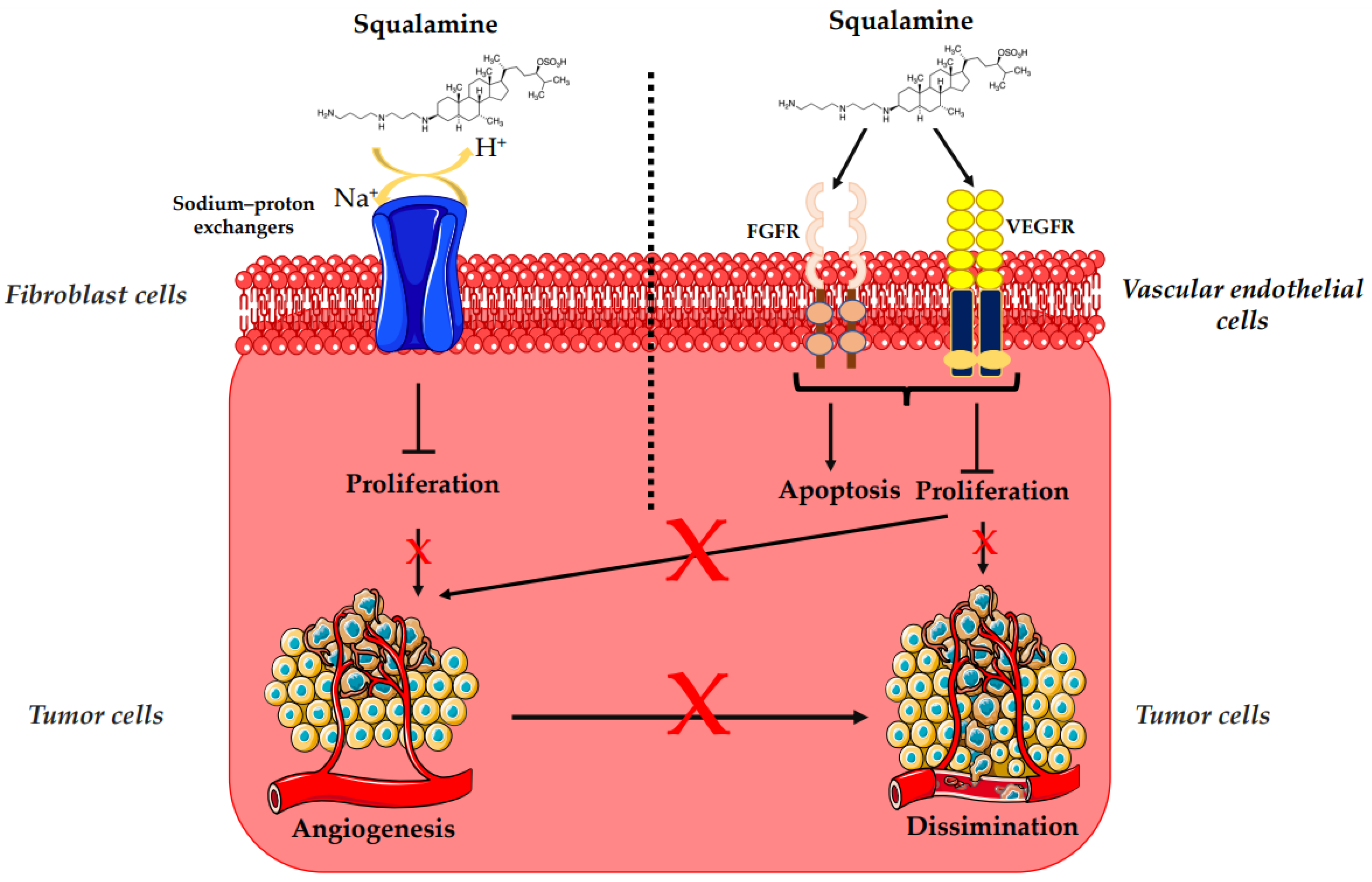
3.2. Antiangiogenesis and Antitumor Activity of Squalamine and Its Derivatives
3.3. Potential Effects of Squalamine on Neurodegenerative Diseases and Other Pathologies
3.3.1. Parkinson’s Disease
3.3.2. Retinopathies and Ocular Neovascularization
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kijjoa, A.; Sawangwong, P. Drugs and Cosmetics from the Sea. Mar. Drugs 2004, 2, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Criscitiello, M.F. What the Shark Immune System Can and Cannot Provide for the Expanding Design Landscape of Immunotherapy. Expert Opin. Drug Discov. 2014, 9, 725–739. [Google Scholar] [CrossRef] [PubMed]
- Malve, H. Exploring the Ocean for New Drug Developments: Marine Pharmacology. J. Pharm. Bioallied Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef]
- Alhanout, K.; Rolain, J.M.; Brunel, J. Squalamine as an Example of a New Potent Antimicrobial Agents Class: A Critical Review. Curr. Med. Chem. 2010, 17, 3909–3917. [Google Scholar] [CrossRef]
- Márquez-Garbán, D.C.; Gorrín-Rivas, M.; Chen, H.-W.; Sterling, C.; Elashoff, D.; Hamilton, N.; Pietras, R.J. Squalamine Blocks Tumor-Associated Angiogenesis and Growth of Human Breast Cancer Cells with or without HER-2/Neu Overexpression. Cancer Lett. 2019, 449, 66–75. [Google Scholar] [CrossRef]
- Boman, H.G. Antibacterial Peptides: Key Components Needed in Immunity. Cell 1991, 65, 205–207. [Google Scholar] [CrossRef]
- Moore, K.S.; Wehrli, S.; Roder, H.; Rogers, M.; Forrest, J.N.; McCrimmon, D.; Zasloff, M. Squalamine: An Aminosterol Antibiotic from the Shark. Proc. Natl. Acad. Sci. USA 1993, 90, 1354–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhanout, K.; Brunel, J.M.; Ranque, S.; Rolain, J.M. In Vitro Antifungal Activity of Aminosterols against Moulds Isolated from Cystic Fibrosis Patients. J. Antimicrob. Chemother. 2010, 65, 1307–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhanout, K.; Malesinki, S.; Vidal, N.; Peyrot, V.; Rolain, J.M.; Brunel, J.M. New Insights into the Antibacterial Mechanism of Action of Squalamine. J. Antimicrob. Chemother. 2010, 65, 1688–1693. [Google Scholar] [CrossRef]
- Rao, M.N.; Shinnar, A.E.; Noecker, L.A.; Chao, T.L.; Feibush, B.; Snyder, B.; Sharkansky, I.; Sarkahian, A.; Zhang, X.; Jones, S.R.; et al. Aminosterols from the Dogfish Shark Squalus Acanthias. J. Nat. Prod. 2000, 63, 631–635. [Google Scholar] [CrossRef]
- Zasloff, M.; Adams, A.P.; Beckerman, B.; Campbell, A.; Han, Z.; Luijten, E.; Meza, I.; Julander, J.; Mishra, A.; Qu, W.; et al. Squalamine as a Broad-Spectrum Systemic Antiviral Agent with Therapeutic Potential. Proc. Natl. Acad. Sci. USA 2011, 108, 15978–15983. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Williams, J.I.; Pietras, R.J. Squalamine and Cisplatin Block Angiogenesis and Growth of Human Ovarian Cancer Cells with or without HER-2 Gene Overexpression. Oncogene 2002, 21, 2805–2814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmi, C.; Loncle, C.; Vidal, N.; Letourneux, Y.; Fantini, J.; Maresca, M.; Taïeb, N.; Pagès, J.-M.; Brunel, J.M. Squalamine: An Appropriate Strategy against the Emergence of Multidrug Resistant Gram-Negative Bacteria? PLoS ONE 2008, 3, e2765. [Google Scholar] [CrossRef] [PubMed]
- Kazakova, O.; Giniyatullina, G.; Babkov, D.; Wimmer, Z. From Marine Metabolites to the Drugs of the Future: Squalamine, Trodusquemine, Their Steroid and Triterpene Analogues. Int. J. Mol. Sci. 2022, 23, 1075. [Google Scholar] [CrossRef] [PubMed]
- PubChem Squalamine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/72495 (accessed on 6 January 2022).
- Savage, P.B.; Li, C. Cholic Acid Derivatives: Novel Antimicrobials. Expert Opin. Investig. Drugs 2005, 9, 263–272. [Google Scholar] [CrossRef]
- Chen, W.-H.; Shao, X.-B.; Moellering, R.; Wennersten, C.; Regen, S.L. A Bioconjugate Approach toward Squalamine Mimics: Insight into the Mechanism of Biological Action. Bioconjugate Chem. 2006, 17, 1582–1591. [Google Scholar] [CrossRef]
- Lavigne, J.-P.; Brunel, J.-M.; Chevalier, J.; Pagès, J.-M. Squalamine, an Original Chemosensitizer to Combat Antibiotic-Resistant Gram-Negative Bacteria. J. Antimicrob. Chemother. 2010, 65, 799–801. [Google Scholar] [CrossRef]
- Alhanout, K.; Brunel, J.-M.; Raoult, D.; Rolain, J.-M. In Vitro Antibacterial Activity of Aminosterols against Multidrug-Resistant Bacteria from Patients with Cystic Fibrosis. J. Antimicrob. Chemother. 2009, 64, 810–814. [Google Scholar] [CrossRef]
- Djouhri-Bouktab, L.; Alhanout, K.; Andrieu, V.; Stremler, N.; Dubus, J.C.; Raoult, D.; Rolain, J.M.; Brunel, J.M. Soluble Squalamine Tablets for the Rapid Disinfection of Home Nebulizers of Cystic Fibrosis Patients. J. Cyst. Fibros. 2012, 11, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Alhanout, K.; Djouhri, L.; Vidal, N.; Brunel, J.M.; Piarroux, R.; Ranque, S. In Vitro Activity of Aminosterols against Yeasts Involved in Blood Stream Infections. Med. Mycol. 2011, 49, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Hraiech, S.; Brégeon, F.; Brunel, J.-M.; Rolain, J.-M.; Lepidi, H.; Andrieu, V.; Raoult, D.; Papazian, L.; Roch, A. Antibacterial Efficacy of Inhaled Squalamine in a Rat Model of Chronic Pseudomonas Aeruginosa Pneumonia. J. Antimicrob. Chemother. 2012, 67, 2452–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicol, M.; Mlouka, M.A.B.; Berthe, T.; Di Martino, P.; Jouenne, T.; Brunel, J.-M.; Dé, E. Anti-Persister Activity of Squalamine against Acinetobacter Baumannii. Int. J. Antimicrob. Agents 2019, 53, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Djouhri-Bouktab, L.; Alhanout, K.; Andrieu, V.; Raoult, D.; Rolain, J.M.; Brunel, J.M. Squalamine Ointment for Staphylococcus aureus Skin Decolonization in a Mouse Model. J. Antimicrob. Chemother. 2011, 66, 1306–1310. [Google Scholar] [CrossRef] [Green Version]
- Coulibaly, O.; Alhanout, K.; L’Ollivier, C.; Brunel, J.-M.; Thera, M.A.; Djimdé, A.A.; Doumbo, O.K.; Piarroux, R.; Ranque, S. In Vitro Activity of Aminosterols against Dermatophytes. Med. Mycol. 2013, 51, 309–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khabnadideh, S.; Tan, C.L.; Croft, S.L.; Kendrick, H.; Yardley, V.; Gilbert, I.H. Squalamine Analogues as Potential Anti-Trypanosomal and Anti-Leishmanial Compounds. Bioorg. Med. Chem. Lett. 2000, 10, 1237–1239. [Google Scholar] [CrossRef]
- Sills, A.K.; Williams, J.I.; Tyler, B.M.; Epstein, D.S.; Sipos, E.P.; Davis, J.D.; McLane, M.P.; Pitchford, S.; Cheshire, K.; Gannon, F.H.; et al. Squalamine Inhibits Angiogenesis and Solid Tumor Growth in Vivo and Perturbs Embryonic Vasculature. Cancer Res. 1998, 58, 2784–2792. [Google Scholar] [PubMed]
- Teicher, B.A.; Williams, J.I.; Takeuchi, H.; Ara, G.; Herbst, R.S.; Buxton, D. Potential of the Aminosterol, Squalamine in Combination Therapy in the Rat 13,762 Mammary Carcinoma and the Murine Lewis Lung Carcinoma. Anticancer Res. 1998, 18, 2567–2573. [Google Scholar]
- Bhargava, P.; Marshall, J.L.; Dahut, W.; Rizvi, N.; Trocky, N.; Williams, J.I.; Hait, H.; Song, S.; Holroyd, K.J.; Hawkins, M.J. A Phase I and Pharmacokinetic Study of Squalamine, a Novel Antiangiogenic Agent, in Patients with Advanced Cancers. Clin. Cancer Res. 2001, 7, 3912–3919. [Google Scholar] [PubMed]
- Hao, D.; Hammond, L.A.; Eckhardt, S.G.; Patnaik, A.; Takimoto, C.H.; Schwartz, G.H.; Goetz, A.D.; Tolcher, A.W.; McCreery, H.A.; Mamun, K.; et al. A Phase I and Pharmacokinetic Study of Squalamine, an Aminosterol Angiogenesis Inhibitor. Clin. Cancer Res. 2003, 9, 2465–2471. [Google Scholar]
- Herbst, R.S.; Hammond, L.A.; Carbone, D.P.; Tran, H.T.; Holroyd, K.J.; Desai, A.; Williams, J.I.; Bekele, B.N.; Hait, H.; Allgood, V.; et al. A Phase I/IIA Trial of Continuous Five-Day Infusion of Squalamine Lactate (MSI-1256F) Plus Carboplatin and Paclitaxel in Patients with Advanced Non-Small Cell Lung Cancer. Clin. Cancer Res. 2003, 9, 4108–4115. [Google Scholar]
- Williams, J.I.; Weitman, S.; Gonzalez, C.M.; Jundt, C.H.; Marty, J.; Stringer, S.D.; Holroyd, K.J.; Mclane, M.P.; Chen, Q.; Zasloff, M.; et al. Squalamine Treatment of Human Tumors in Nu/Nu Mice Enhances Platinum-Based Chemotherapies. Clin. Cancer Res. 2001, 7, 724–733. [Google Scholar] [PubMed]
- Akhter, S.; Nath, S.K.; Tse, C.M.; Williams, J.; Zasloff, M.; Donowitz, M. Squalamine, a Novel Cationic Steroid, Specifically Inhibits the Brush-Border Na+/H+ Exchanger Isoform NHE3. Am. J. Physiol. 1999, 276, C136–C144. [Google Scholar] [CrossRef] [PubMed]
- Pietras, R.J.; Weinberg, O.K. Antiangiogenic Steroids in Human Cancer Therapy. Evid. Based Complement. Alternat. Med. 2005, 2, 49–57. [Google Scholar] [CrossRef] [Green Version]
- West, C.L.; Mao, Y.-K.; Delungahawatta, T.; Amin, J.Y.; Farhin, S.; McQuade, R.M.; Diwakarla, S.; Pustovit, R.; Stanisz, A.M.; Bienenstock, J.; et al. Squalamine Restores the Function of the Enteric Nervous System in Mouse Models of Parkinson’s Disease. J. Parkinsons Dis. 2020, 10, 1477–1491. [Google Scholar] [CrossRef] [PubMed]
- Ciulla, T.A.; Criswell, M.H.; Danis, R.P.; Williams, J.I.; McLane, M.P.; Holroyd, K.J. Squalamine Lactate Reduces Choroidal Neovascularization in a Laser-Injury Model in the Rat. Retina 2003, 23, 808–814. [Google Scholar] [CrossRef]
- Genaidy, M.; Kazi, A.A.; Peyman, G.A.; Passos-Machado, E.; Farahat, H.G.; Williams, J.I.; Holroyd, K.J.; Blake, D.A. Effect of Squalamine on Iris Neovascularization in Monkeys. Retina 2002, 22, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Higgins, R.D.; Sanders, R.J.; Yan, Y.; Zasloff, M.; Williams, J.I. Squalamine Improves Retinal Neovascularization. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1507–1512. [Google Scholar]
- Carmona, S.; Brunel, J.-M.; Bonier, R.; Sbarra, V.; Robert, S.; Borentain, P.; Lombardo, D.; Mas, E.; Gerolami, R. A Squalamine Derivative, NV669, as a Novel PTP1B Inhibitor: In Vitro and In Vivo Effects on Pancreatic and Hepatic Tumor Growth. Oncotarget 2019, 10, 6651–6667. [Google Scholar] [CrossRef] [Green Version]
- Gallo, S.W.; Donamore, B.K.; Pagnussatti, V.E.; Ferreira, C.A.S.; de Oliveira, S.D. Effects of Meropenem Exposure in Persister Cells of Acinetobacter Calcoaceticus-Baumannii. Future Microbiol. 2017, 12, 131–140. [Google Scholar] [CrossRef]
- Chung, E.S.; Wi, Y.M.; Ko, K.S. Variation in Formation of Persister Cells against Colistin in Acinetobacter Baumannii Isolates and Its Relationship with Treatment Failure. J. Antimicrob. Chemother. 2017, 72, 2133–2135. [Google Scholar] [CrossRef]
- Bhargava, N.; Sharma, P.; Capalash, N. Pyocyanin Stimulates Quorum Sensing-Mediated Tolerance to Oxidative Stress and Increases Persister Cell Populations in Acinetobacter baumannii. Infect. Immun. 2014, 82, 3417–3425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cushnie, T.P.T.; Cushnie, B.; Lamb, A.J. Alkaloids: An Overview of Their Antibacterial, Antibiotic-Enhancing and Antivirulence Activities. Int. J. Antimicrob. Agents 2014, 44, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Tsubery, H.; Ofek, I.; Cohen, S.; Eisenstein, M.; Fridkin, M. Modulation of the Hydrophobic Domain of Polymyxin B Nonapeptide: Effect on Outer-Membrane Permeabilization and Lipopolysaccharide Neutralization. Mol. Pharmacol. 2002, 62, 1036–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clausell, A.; Rabanal, F.; Garcia-Subirats, M.; Asunción Alsina, M.; Cajal, Y. Synthesis and Membrane Action of Polymyxin B Analogues. Luminescence 2005, 20, 117–123. [Google Scholar] [CrossRef]
- Di Pasquale, E.; Salmi-Smail, C.; Brunel, J.-M.; Sanchez, P.; Fantini, J.; Maresca, M. Biophysical Studies of the Interaction of Squalamine and Other Cationic Amphiphilic Molecules with Bacterial and Eukaryotic Membranes: Importance of the Distribution Coefficient in Membrane Selectivity. Chem. Phys. Lipids 2010, 163, 131–140. [Google Scholar] [CrossRef]
- Kaptain, S.; Tan, L.K.; Chen, B. Her-2/Neu and Breast Cancer. Diagn. Mol. Pathol. 2001, 10, 139–152. [Google Scholar] [CrossRef]
- Dore-Savard, L.; Lee, E.; Kakkad, S.; Popel, A.S.; Bhujwalla, Z.M. The Angiogenic Secretome in VEGF Overexpressing Breast Cancer Xenografts. Sci. Rep. 2016, 6, 39460. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann-La Roche. A Randomized, Open-Label Study to Compare the Effect of First-Line Treatment with Avastin in Combination with Herceptin/Docetaxel and Herceptin/Docetaxel Alone on Progression-Free Survival in Patients with HER2 Positive Locally Recurrent or Metastatic Breast Cancer. 2015. Available online: clinicaltrials.gov (accessed on 16 September 2021).
- Gianni, L.; Romieu, G.H.; Lichinitser, M.; Serrano, S.V.; Mansutti, M.; Pivot, X.; Mariani, P.; Andre, F.; Chan, A.; Lipatov, O.; et al. AVEREL: A Randomized Phase III Trial Evaluating Bevacizumab in Combination with Docetaxel and Trastuzumab as First-Line Therapy for HER2-Positive Locally Recurrent/Metastatic Breast Cancer. J. Clin. Oncol. 2013, 31, 1719–1725. [Google Scholar] [CrossRef]
- Zhao, M.; Pan, X.; Layman, R.; Lustberg, M.B.; Mrozek, E.; Macrae, E.R.; Wesolowski, R.; Carothers, S.; Puhalla, S.; Shapiro, C.L.; et al. A Phase II Study of Bevacizumab in Combination with Trastuzumab and Docetaxel in HER2 Positive Metastatic Breast Cancer. Investig. New Drugs 2014, 32, 1285–1294. [Google Scholar] [CrossRef]
- Limbocker, R.; Errico, S.; Barbut, D.; Knowles, T.P.J.; Vendruscolo, M.; Chiti, F.; Zasloff, M. Squalamine and Trodusquemine: Two Natural Products for Neurodegenerative Diseases, from Physical Chemistry to the Clinic. Nat. Prod. Rep. 2022, 39, 742–753. [Google Scholar] [CrossRef]
- Salmi, C.; Loncle, C.; Vidal, N.; Laget, M.; Letourneux, Y.; Michel Brunel, J. Antimicrobial Activities of 3-Amino- and Polyaminosterol Analogues of Squalamine and Trodusquemine. J. Enzyme Inhib. Med. Chem. 2008, 23, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Djouhri-Bouktab, L.; Vidal, N.; Rolain, J.M.; Brunel, J.M. Synthesis of New 3,20-Bispolyaminosteroid Squalamine Analogues and Evaluation of Their Antimicrobial Activities. J. Med. Chem. 2011, 54, 7417–7421. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-H.; Wennersten, C.; Moellering, R.C.; Regen, S.L. Towards Squalamine Mimics: Synthesis and Antibacterial Activities of Head-to-Tail Dimeric Sterol-Polyamine Conjugates. Chem. Biodivers. 2013, 10, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent Bacterial Infections and Persister Cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef]
- Möhrenschlager, M.; Seidl, H.P.; Ring, J.; Abeck, D. Pediatric Tinea Capitis: Recognition and Management. Am. J. Clin. Dermatol. 2005, 6, 203–213. [Google Scholar] [CrossRef]
- Coulibaly, O.; Thera, M.A.; Koné, A.K.; Siaka, G.; Traoré, P.; Djimdé, A.A.; Brunel, J.-M.; Gaudart, J.; Piarroux, R.; Doumbo, O.K.; et al. A Double-Blind Randomized Placebo-Controlled Clinical Trial of Squalamine Ointment for Tinea Capitis Treatment. Mycopathologia 2015, 179, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Alexander, R.T.; Jaumouillé, V.; Yeung, T.; Furuya, W.; Peltekova, I.; Boucher, A.; Zasloff, M.; Orlowski, J.; Grinstein, S. Membrane Surface Charge Dictates the Structure and Function of the Epithelial Na+/H+ Exchanger. EMBO J. 2011, 30, 679–691. [Google Scholar] [CrossRef] [Green Version]
- Wroblewski, J.J.; Hu, A.Y. Topical Squalamine 0.2% and Intravitreal Ranibizumab 0.5 Mg as Combination Therapy for Macular Edema Due to Branch and Central Retinal Vein Occlusion: An Open-Label, Randomized Study. Ophthalmic Surg. Lasers Imaging Retina 2016, 47, 914–923. [Google Scholar] [CrossRef]
- Connolly, B.; Desai, A.; Garcia, C.A.; Thomas, E.; Gast, M.J. Squalamine Lactate for Exudative Age-Related Macular Degeneration. Ophthalmol. Clin. N. Am. 2006, 19, 381–391, vi. [Google Scholar] [CrossRef]
- Hussain, R.M.; Ciulla, T.A. Emerging Vascular Endothelial Growth Factor Antagonists to Treat Neovascular Age-Related Macular Degeneration. Expert Opin. Emerg. Drugs 2017, 22, 235–246. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Squalamine and Derivatives | Structure | Pharmacological Activity | References |
|---|---|---|---|
| Squalamine |  | Antibacterial activity: Escherichia coli (ATCC 25922, ATCC 54127); Pseudomonas aeruginosa (ATCC 27853, strain PAO1, ATCC 1569, ATCC 15442); Staphylococcus aureus (ATCC 25923, ATCC 6538); Streptococcus pneumoniae (a clinical isolate); Acinetobacter baumannii Antifungal activity: Candida albicans (ATCC 10231, (ATCC 90028); Aspergillus niger (ATCC 16404); Candida glabrata (ATCC 90030); Candida krusei (ATCC 6258); Candida parapsilosis (ATCC 22019) Bloodstream yeast isolates: C. albicans; C. glabrata; C. guilliermondii; C. krusei; C. lusitaniae; C. parapsilosis; C. tropicalis, Cryptococcus neoformans; Trichophyton rubrum; T. mentagrophytes; T. soudanense; Microsporum canis; M. audouinii; M. persicolor; M. cookie; M. gypseum; Tinea capitis Antiviral activity: Dengue virus; Human hepatitis B virus; Human hepatitis δ-virus; Yellow fever virus; Eastern equine encephalitis virus; Murine cytomegalovirus Antiprotozoa activity: Trypanosoma brucei; Leishmania donovani Eucaryote cells: Eukaryote cell (Wehi-231 cells) Antiangiogenic activity: Chicken embryo; MCF-7 (Michigan Cancer Foundation-7); Human breast cancer cell line; Pancreatic (BxPC-3, MiaPaCa-2) and hepatic (HepG2, Huh7) cancer cells; Rat mammary carcinoma and a murine Lewis lung carcinoma; Xenograft models using the chemoresistant MV-522 human non-small cell lung carcinoma and the SD human neuroblastoma lines; PS120/NHE3 fibroblasts cells; Rat brain endothelial (RBE-4) cell; Rabbit VX2 tumor cells Clinical trials for cancer cases: Patients with advanced cancers: Patients with metastasis to the central nervous system; Patients with liver metastasis; Patients with a histologically confirmed diagnosis of nonleukemic malignancy refractory; Patients with advanced solid malignancies; Patients with advanced non-small cell lung cancer Parkinson’s disease: Effect on the gastrointestinal tract (constipation) and neuron motility Retinopathies: Rat choroidal neovascular membrane; Iris neovascularization in cynomolgus monkeys; Ocular neovascularization Clinical trials for Retinopathies: Patients with macular edema | [5,8,9,11,15,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52] |
| Synthesized aminosterol derivatives (ASD) ASD 1 [7-(1,4-diaminobutane)-cholest-5-ene-3β-ol] |  | Antibacterial activity: E. coli (ATCC 25922); P. aeruginosa (ATCC 27853); S. aureus (ATCC 25923); S. pneumoniae isolates Antifungal: C. albicans (ATCC 90028); C. glabrata (ATCC 90030); C. krusei (ATCC 6258); C. parapsilosis (ATCC 22019) Bloodstream yeast isolates: C. albicans; C. glabrata; C. guilliermondii, C. krusei; C. lusitaniae ; C. parapsilosis; C. tropicali; C. neoformans | [8,18,20] |
| ASD 2 [7β-(1,4-diaminobutane)-cholestan-3β-ol] |  | Antibacterial activity: E. coli (ATCC 25922); P. aeruginosa (ATCC 27853); S. aureus (ATCC 25923); S. pneumoniae isolates | [18,32] |
| 3-amino- and polyaminosterol analogues of squalamine and trodusquemine 4b, 4e, 4n, 4r, 6b, 8b, 8c, 8d, 8e |  | Antibacterial activity: E. coli (ATCC 10536); S. aureus (ATCC 6538); Enterococcus faecalis (CIP 103015) Antifungal activity: C. albicans (ATCC 90029); C. tropicalis (CIP 2031); Saccharomyces cerevisiae (ATCC 28383) | [53] |
| 3, 20-amino- and polyaminosteroid analogues of squalamine and trodusquemine |  | Antibacterial activity: S. aureus; P. aeruginosa; Inquilinus limosus; Burkholderia cepacia | [54] |
| Dimeric sterol-polyamine conjugates (2, 4a, 4b, 5, 6a, 6b, 6c, 7a, 7b) |  | Antibacterial activity: E. coli (ATCC 25922 and ESBL clinical isolates); K. pneumoniae (clinical isolates); Acinetobacter spp. (clinical isolates); P. aeruginosa (clinical isolates); group A Streptococcus (clinical isolates); coag. neg. Staphylococcus (clinical isolates); S. aureus (A8115 MSSA, A8816 MRSA, A5948 MRSA 32); Enterococcus faecium (ATCC 29212, ATCC 51299, and clinical isolates) | [17] |
| Squalamine Mimics: Head-to-Tail Dimeric SterolPolyamine Conjugates (1–8) |  | Antibacterial activity: E. coli (ATCC 25922 and ESBL clinical isolates); K. pneumoniae (clinical isolates); Acinetobacter spp. (clinical isolates); P. aeruginosa (clinical isolates); E. faecalis (ATCC 29212, ATCC 51299, and clinical isolates); E. faecium (clinical isolates); S. aureus (ATCC 29213, MSSA, and MRSA clinical isolates); Coag. Neg. Staphylococcus (clinical isolates); Group A Streptococcus (clinical isolates) | [55] |
| Squalamine mimics |  | Antibacterial activity: E. coli (ATCC 25922); K. pneumoniae (ATCC 13883); P. aeruginosa (ATCC 27853); E. faecalis (ATCC29212); S. aureus (ATCC 25923); S. pyogenes (ATCC 19615); Antifungal activity: C. albicans (ATCC 90028) | [16] |
| Squalamine analogues |  | Antiprotozoa activity: Leishmania donovani | [26] |
| NV669 |  | Antiangiogenic activity: Pancreatic (BxPC-3, MiaPaCa-2) and hepatic (HepG2, Huh7) cancer cells | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mammari, N.; Salles, E.; Beaussart, A.; El-Kirat-Chatel, S.; Varbanov, M. Squalamine and Its Aminosterol Derivatives: Overview of Biological Effects and Mechanisms of Action of Compounds with Multiple Therapeutic Applications. Microorganisms 2022, 10, 1205. https://doi.org/10.3390/microorganisms10061205
Mammari N, Salles E, Beaussart A, El-Kirat-Chatel S, Varbanov M. Squalamine and Its Aminosterol Derivatives: Overview of Biological Effects and Mechanisms of Action of Compounds with Multiple Therapeutic Applications. Microorganisms. 2022; 10(6):1205. https://doi.org/10.3390/microorganisms10061205
Chicago/Turabian StyleMammari, Nour, Elsa Salles, Audrey Beaussart, Sofiane El-Kirat-Chatel, and Mihayl Varbanov. 2022. "Squalamine and Its Aminosterol Derivatives: Overview of Biological Effects and Mechanisms of Action of Compounds with Multiple Therapeutic Applications" Microorganisms 10, no. 6: 1205. https://doi.org/10.3390/microorganisms10061205
APA StyleMammari, N., Salles, E., Beaussart, A., El-Kirat-Chatel, S., & Varbanov, M. (2022). Squalamine and Its Aminosterol Derivatives: Overview of Biological Effects and Mechanisms of Action of Compounds with Multiple Therapeutic Applications. Microorganisms, 10(6), 1205. https://doi.org/10.3390/microorganisms10061205