
Uptake of Levilactobacillus brevis JCM 1059 by THP-1 Cells via Interaction between SlpB and CAP-1 Promotes Cytokine Production



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Fermentation
2.2. Induction of DC-SIGN and CAP-1 Expression on Differentiated THP-1 Cells
2.3. Sulfo-Cyanine3 Labelling
2.4. Preparation of Slps Coupled Microbeads
2.5. Uptake of LAB and Slps-Microbeads by THP-1 DC
2.6. Cytokine Measurement
2.7. Purification of the SlpB Receptor on THP-1 Cells
2.8. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis and Western Blotting Analysis
2.9. Protein Identification
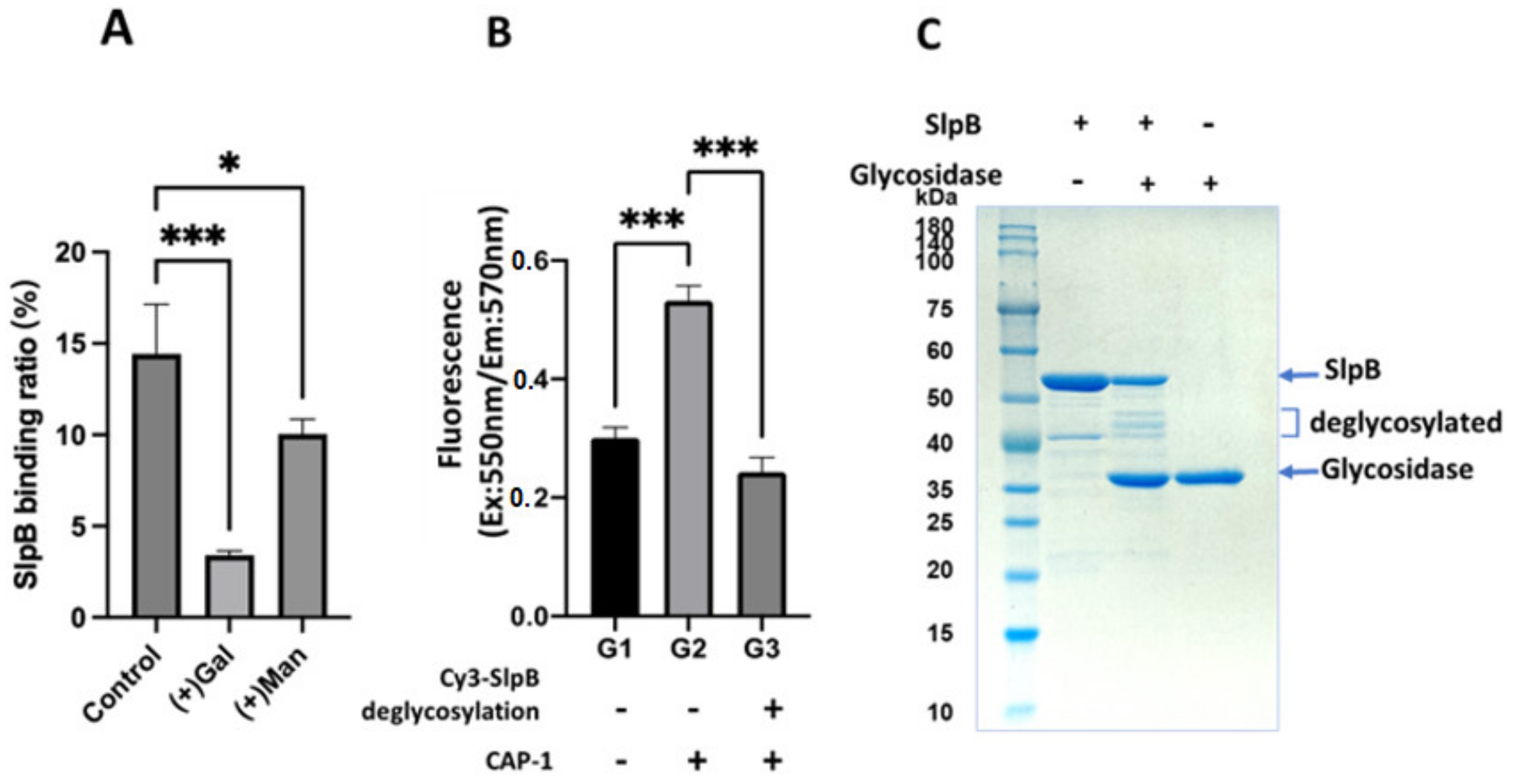
2.10. Binding of Glycan and SlpB to THP-1
2.11. Binding of Deglycosylated SlpB to THP-1
2.12. Statistical Analysis
3. Results
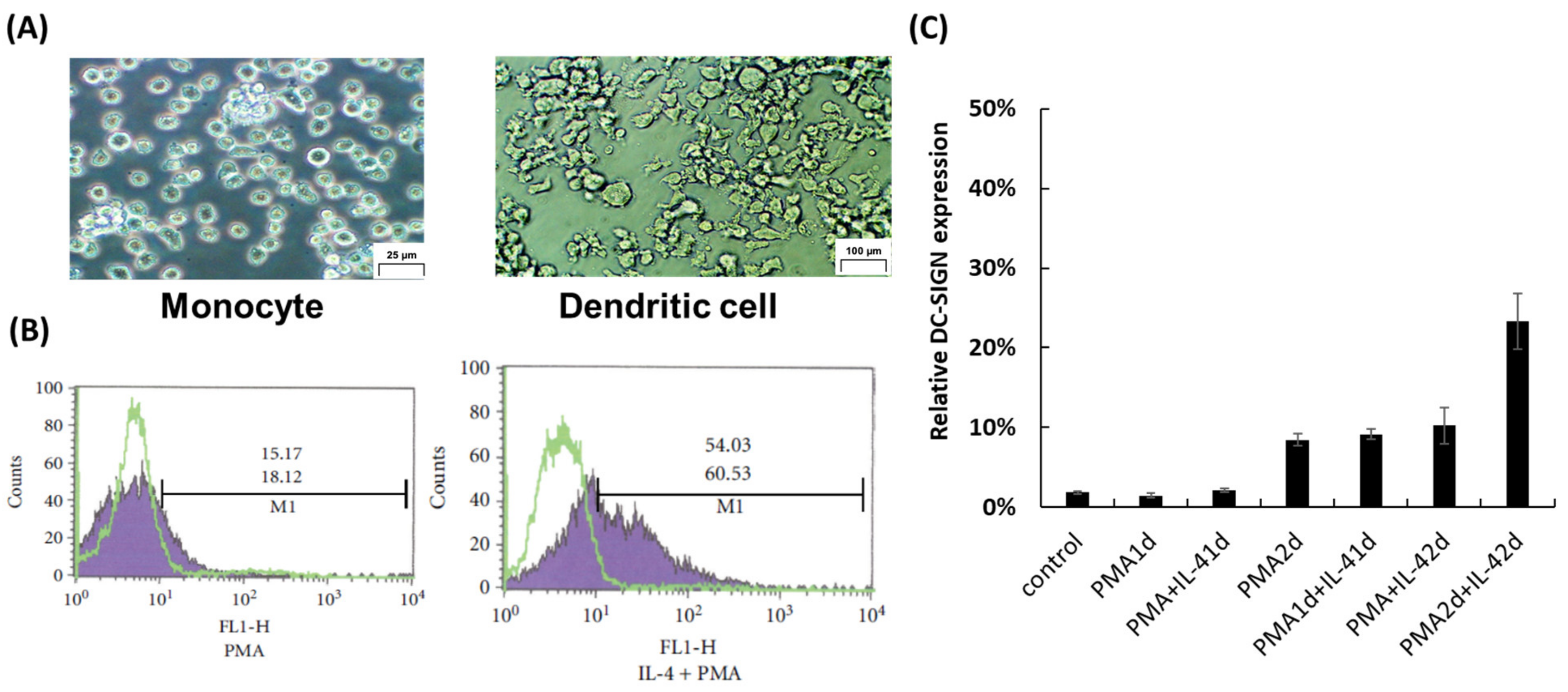
3.1. Induction of DC-SIGN on THP-1 Cells
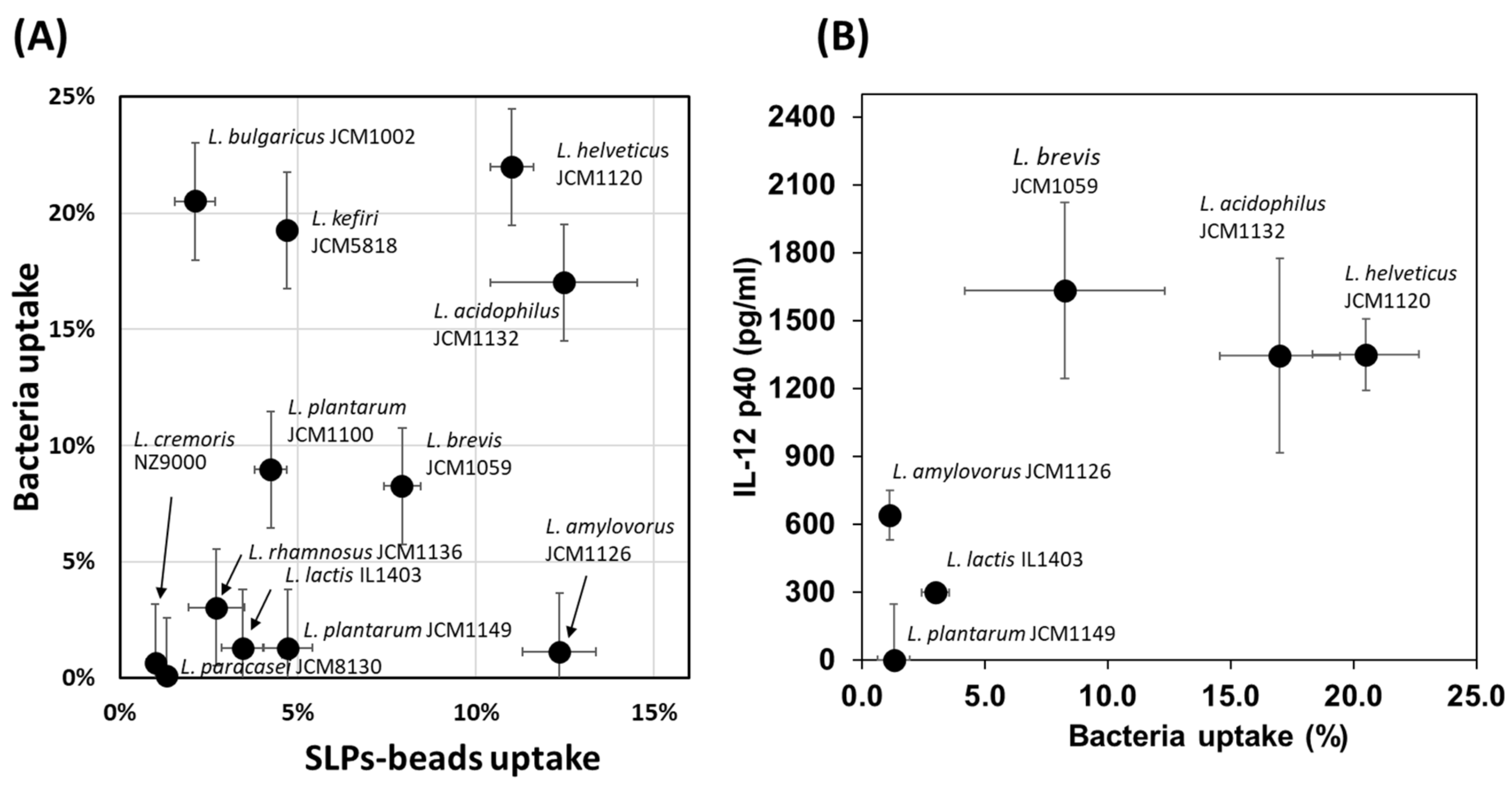
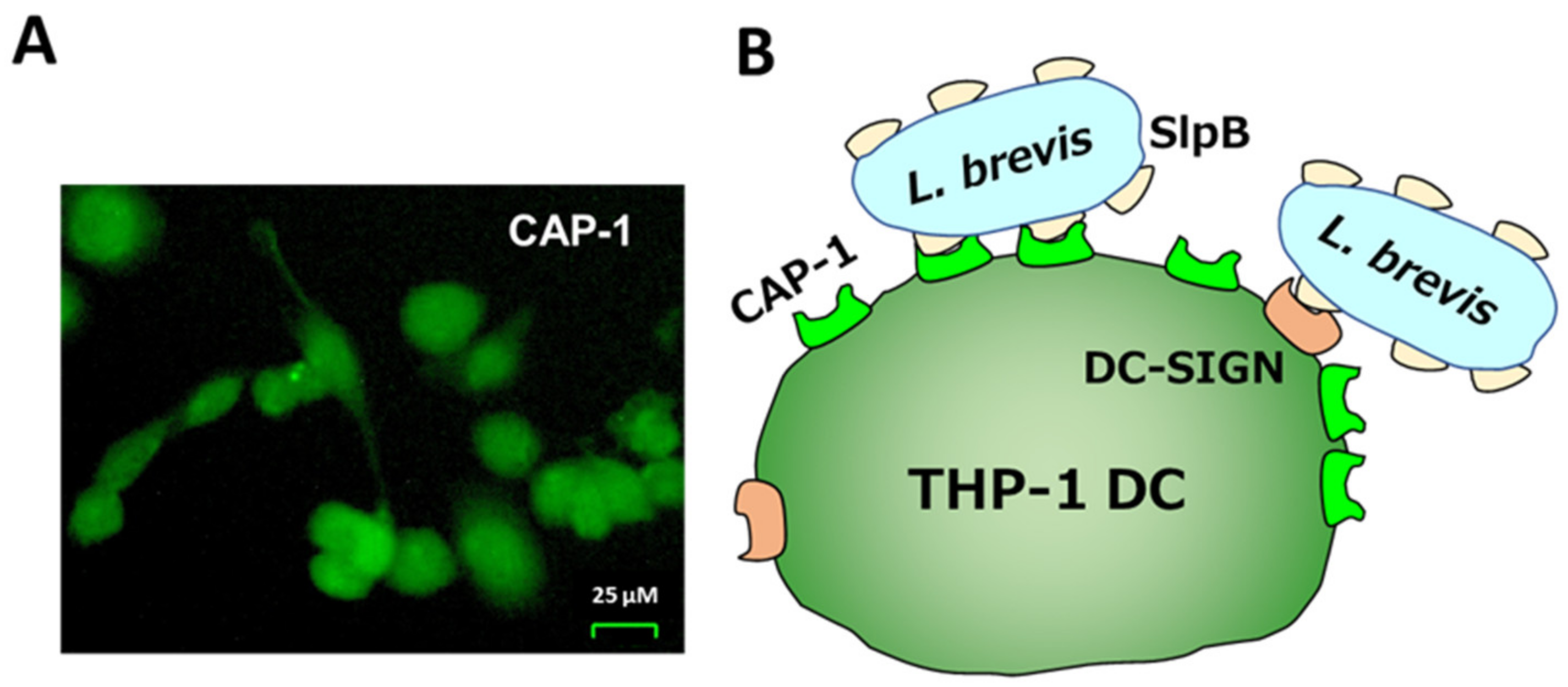
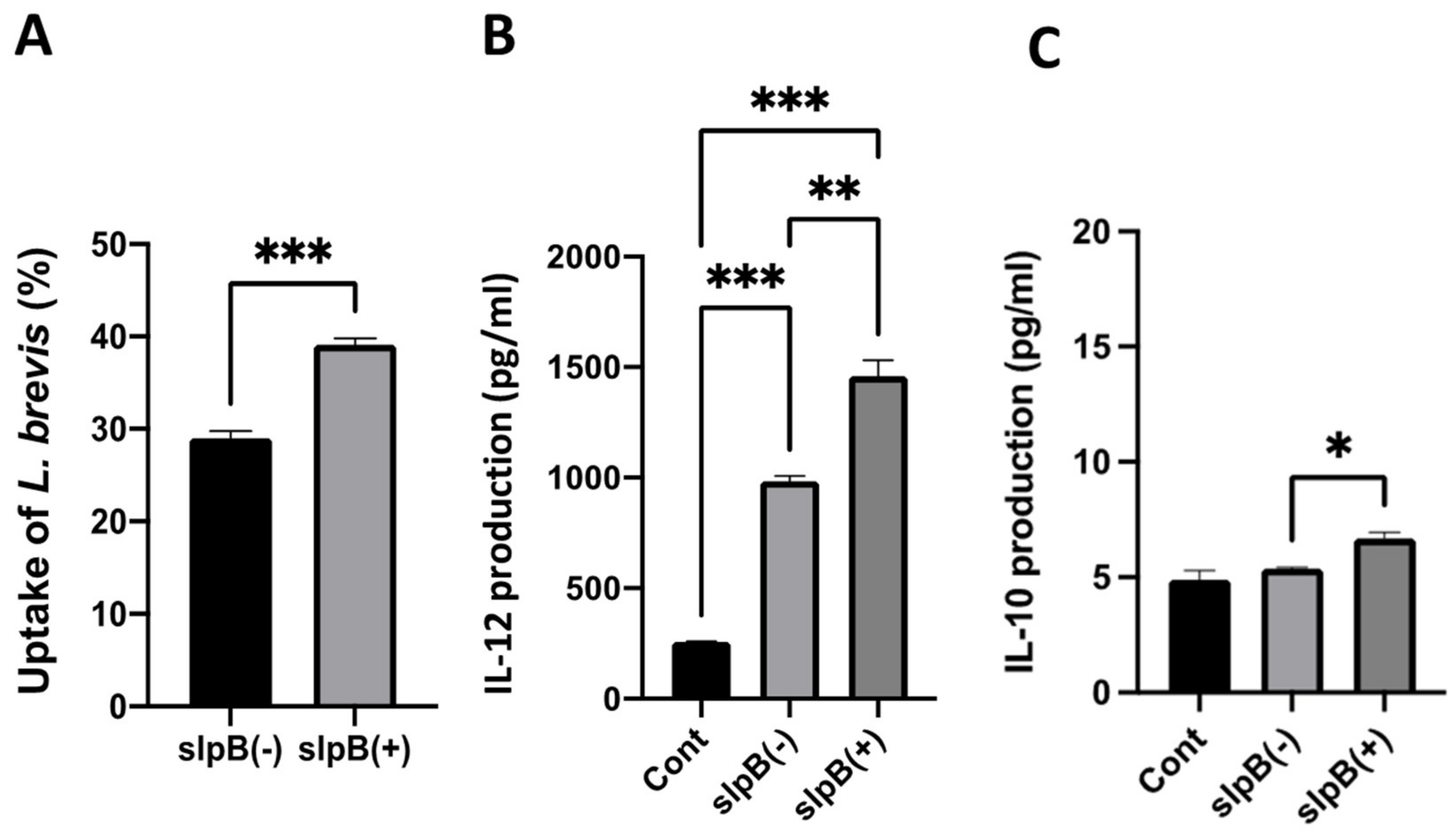
3.2. Uptake of LAB by THP-1 DC and IL-12 Induction
3.3. Slps from Various LAB Strains and Binding to DC-SIGN
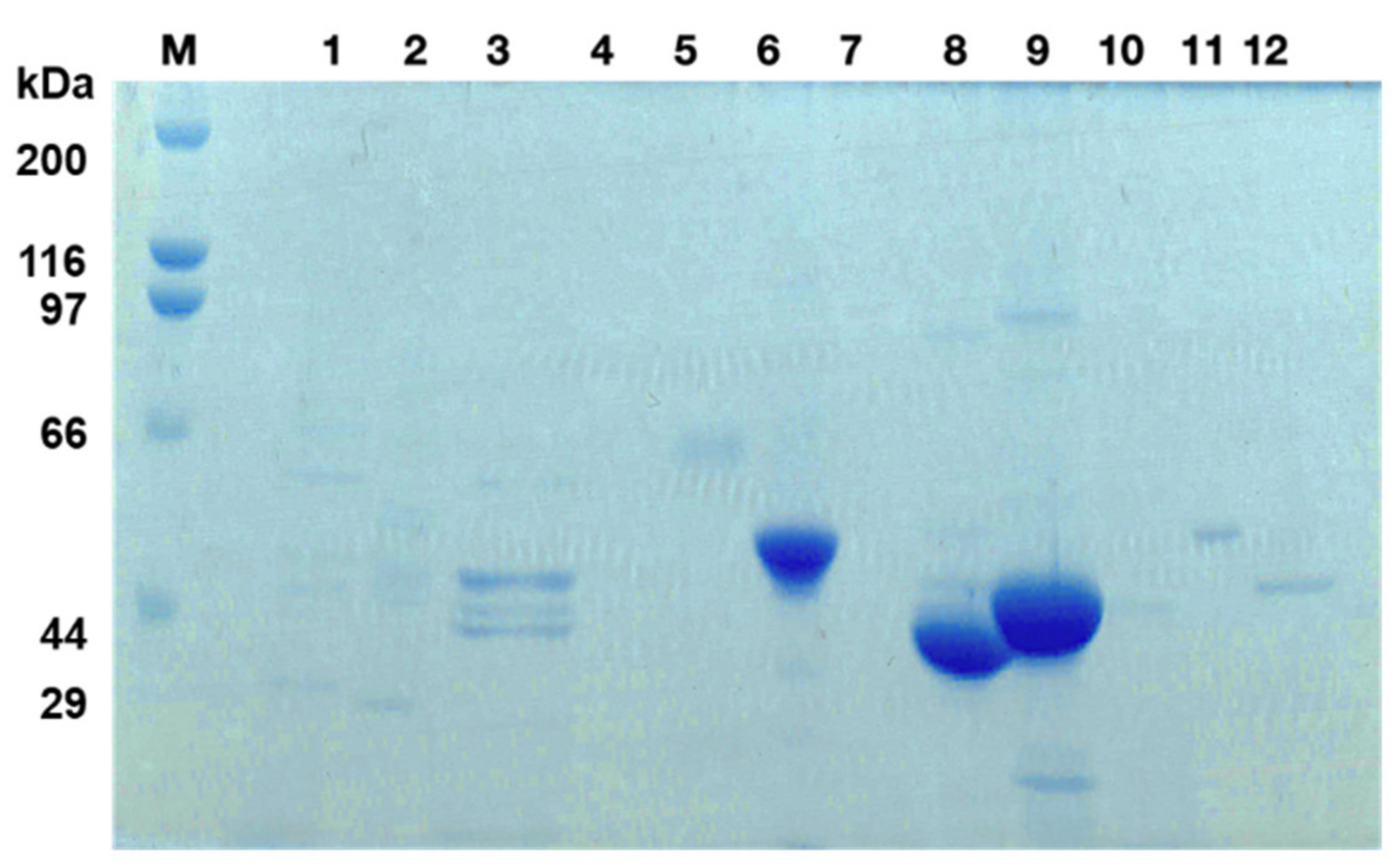
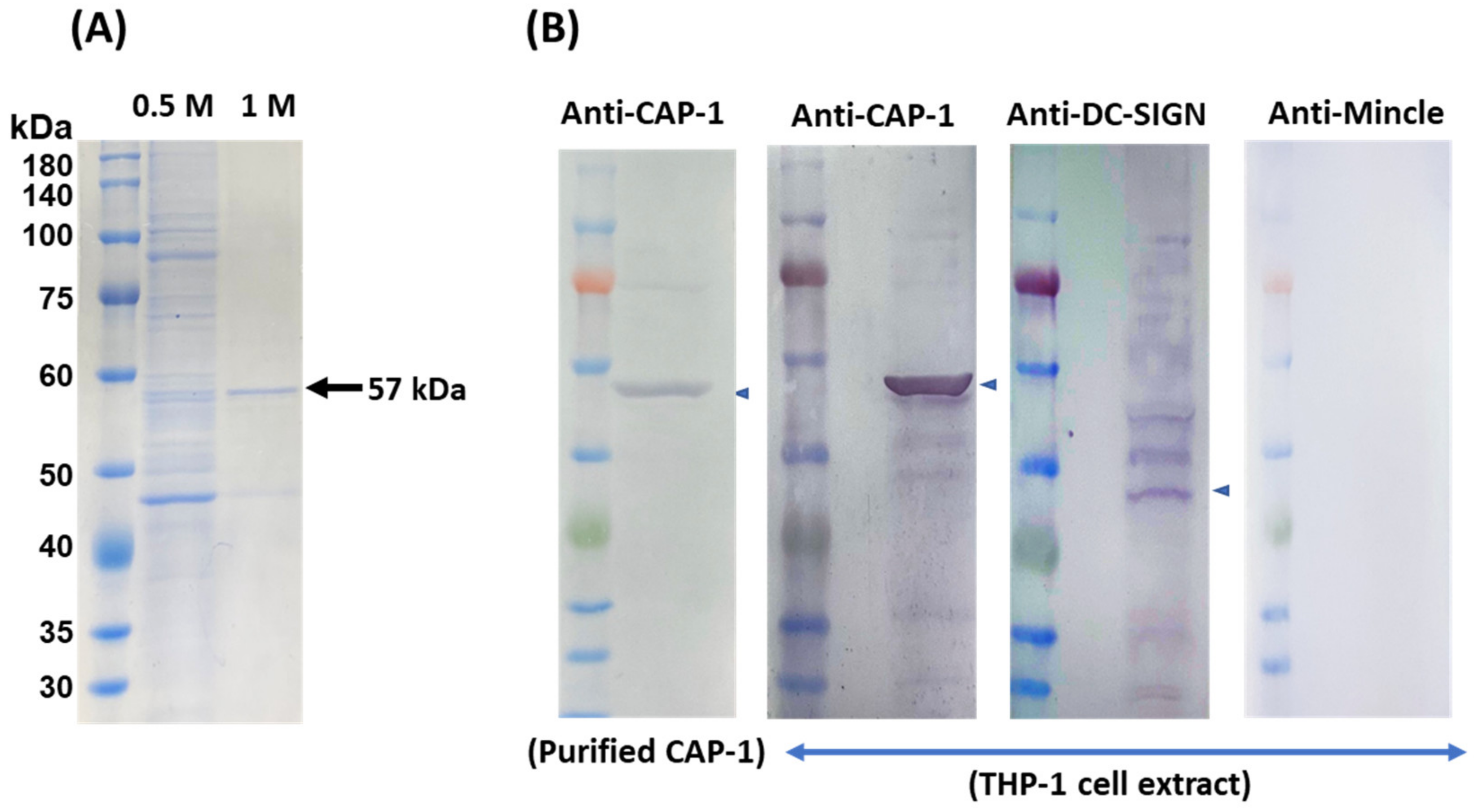
3.4. Purification and Identification of the Receptor for SlpB from THP-1 Cells
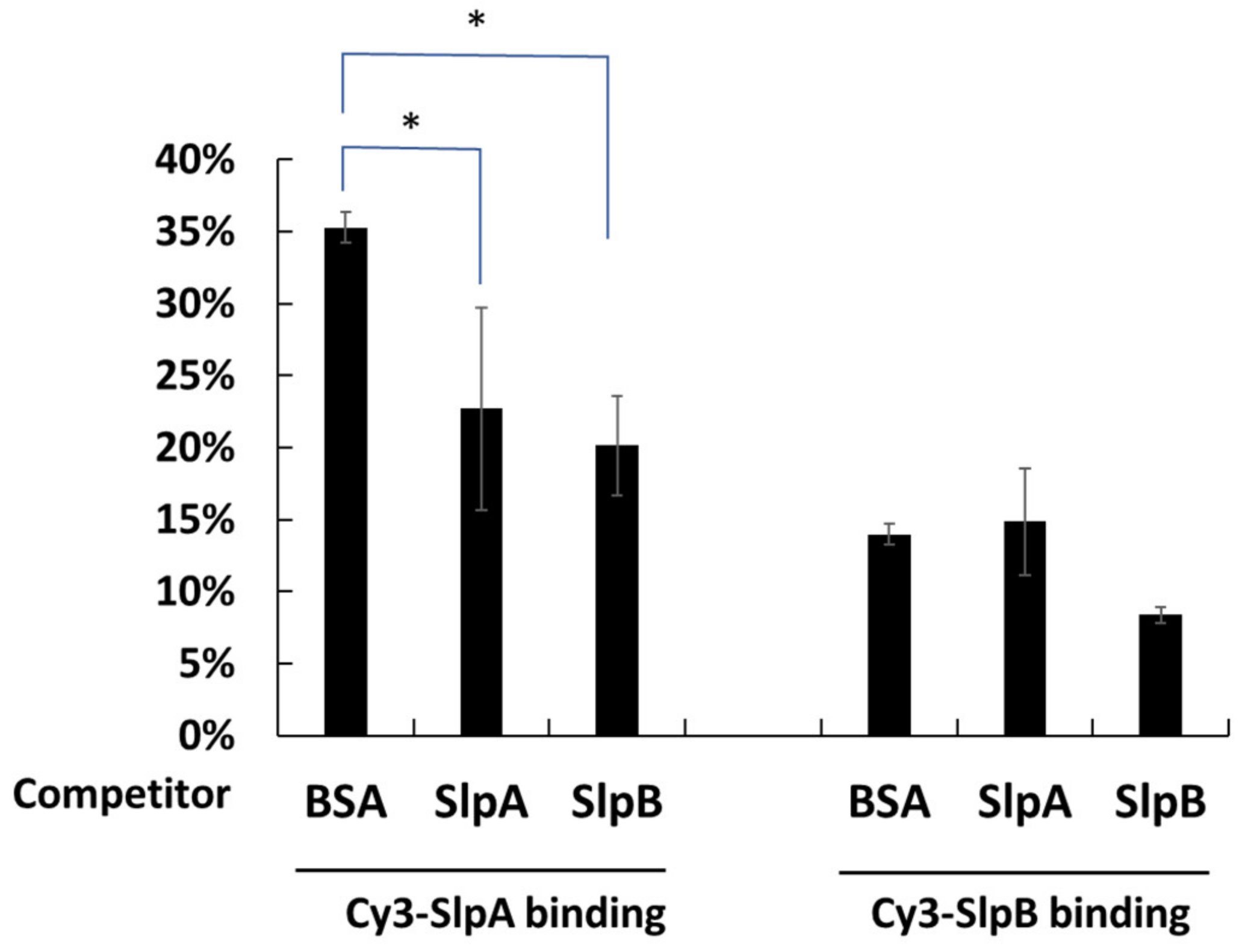
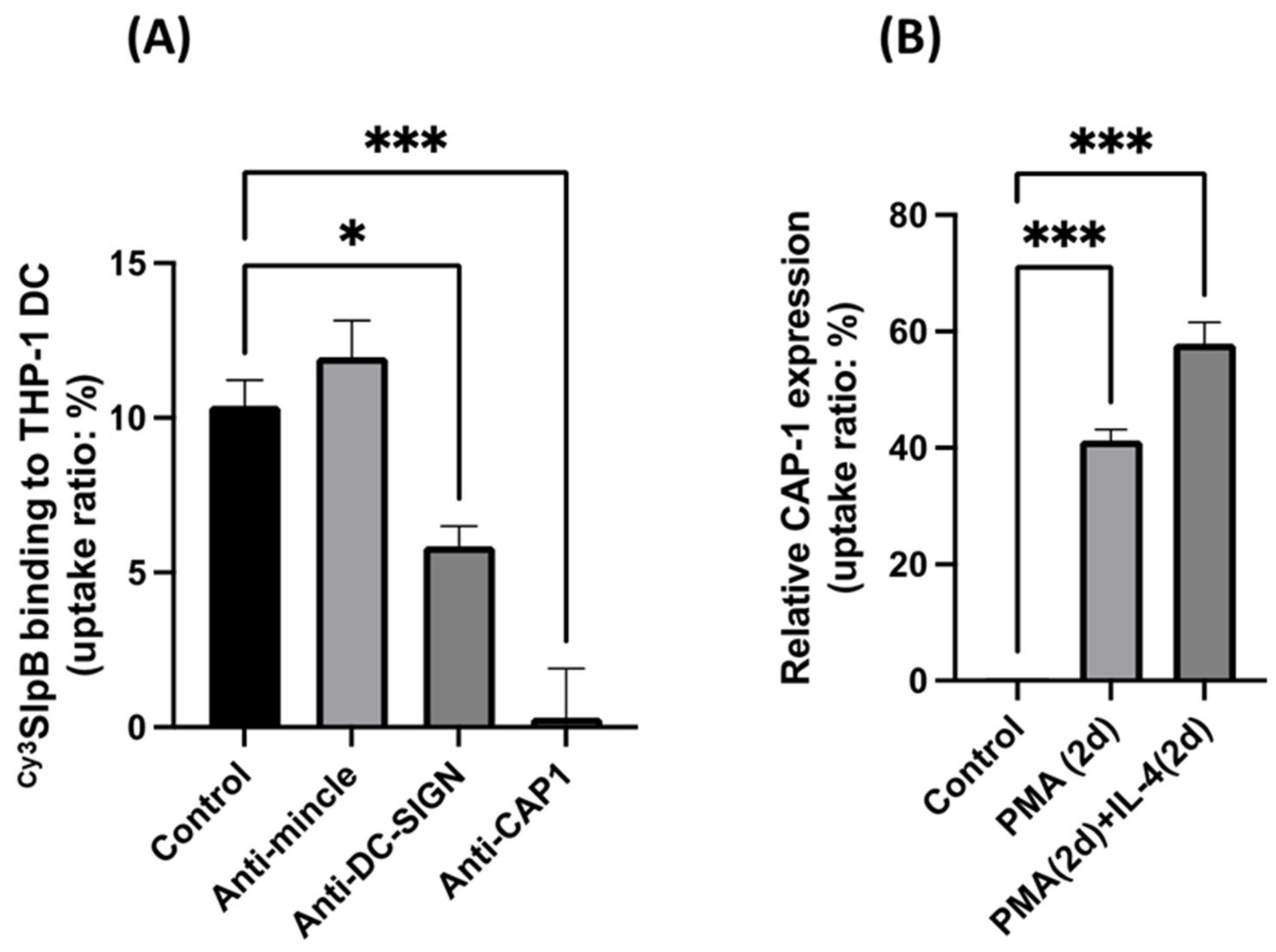
3.5. Inhibition of SlpB Binding to THP-1 DC by Anti-CAP-1, Anti-DC-SIGN, and Anti-Mincle Antibodies
3.6. Cytokine Production in THP-1 DC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Perez, N.G.; de Moreno de LeBlanc, A.; Gomez-Gutierrez, J.; LeBlanc, J.G.; Bermúdez-Humarán, L.G. Probiotics and trained immunity. Biomolecules 2021, 24, 1402. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A long debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Shida, K.; Kiyoshima-Shibata, J.; Nagaoka, M.; Watanabe, K.; Nanno, M. Induction of interleukin-12 by Lactobacillus strains having a rigid cell wall resistant to intracellular digestion. J. Dairy Sci. 2006, 89, 3306–3317. [Google Scholar] [CrossRef] [Green Version]
- Taverniti, V.; Guglielmetti, S. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: Proposal of paraprobiotic concept). Genes Nutr. 2011, 6, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.H.; Kim, S.H.; Jeon, J.H.; Kim, E.B.; Lee, N.K.; Beck, S.; Choi, Y.J.; Kang, S.K. Cytoplasmic expression of a model antigen with M Cell-Targeting moiety in lactic acid bacteria and implication of the mechanism as a mucosal vaccine via oral route. Vaccine 2021, 39, 4072–4081. [Google Scholar] [CrossRef]
- Rahimi, N. C-type lectin CD209L/L-SIGN and CD209/DC-SIGN: Cell adhesion molecules turned to pathogen recognition receptors. Biology 2020, 22, 1. [Google Scholar] [CrossRef]
- Feinberg, H.; Jégouzo, S.A.F.; Rex, M.J.; Drickamer, K.; Weis, W.I.; Taylor, M.E. Mechanism of pathogen recognition by human dectin-2. J. Biol. Chem. 2017, 292, 13402–13414. [Google Scholar] [CrossRef] [Green Version]
- Butcher, M.J.; Zhu, J. Recent advances in understanding the Th1/Th2 effector choice. Fac. Rev. 2021, 10, 30. [Google Scholar] [CrossRef]
- Underhill, D.M.; Ozinsky, A. Toll-like receptors: Key mediators of microbe detection. Curr. Opin. Immunol. 2002, 14, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Thoma-Uszynski, S.; Stenger, S.; Takeuchi, O.; Ochoa, M.T.; Engele, M.; Sieling, P.A.; Barnes, P.F.; Rollinghoff, M.; Bolcskei, P.L.; Wagner, M.; et al. Induction of direct antimicrobial activity through mammalian Toll-like receptors. Science 2001, 291, 1544–1547. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Adema, G.J.; van Kooyk, Y.; Figdor, C.G. Identification of DC-SIGN, a novel dendritic cell-specific ICAM-3 receptor that supports primary immune responses. Cell 2000, 100, 575–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akdis, M.; Verhagen, J.; Taylor, A.; Karamloo, F.; Karagiannidis, C.; Crameri, R.; Thunberg, S.; Deniz, G.; Valenta, R.; Fiebig, H.; et al. Immune responses in healthy and allergic individuals are characterized by a fine balance between allergen-specific T regulatory 1 and T helper 2 cells. J. Exp. Med. 2004, 199, 1567–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaiss, C.A.; Levy, M.; Suez, J.; Elinav, E. The interplay between the innate immune system and the microbiota. Curr. Opin. Immunol. 2014, 26, 41–48. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.Y.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Snyder, S.; Feng, P.; Azadi, P.; Zhang, S.; Bulgheresi, S.; Sanderson, K.E.; He, J.; Klena, J.; Chen, T. Role of N-acetylglucosamine within core lipopolysaccharide of several species of gram-negative bacteria in targeting the DC-SIGN (CD209). J. Immunol. 2006, 177, 4002–4011. [Google Scholar] [CrossRef] [Green Version]
- Iyori, M.; Ohtani, M.; Hasebe, A.; Totsuka, Y.; Shibata, K.-I. A role of the Ca2+ binding site of DC-SIGN in the phagocytosis of E. coli. Biochem. Biophys. Res. Commun. 2008, 377, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Klena, J.; Zhang, P.; Schwartz, O.; Hull, S.; Chen, T. The core lipopolysaccharide of Escherichia coli is a ligand for the dendritic-cell-specific intercellular adhesion molecule nonintegrin CD209 receptor. J. Bacteriol. 2005, 187, 1710–1715. [Google Scholar] [CrossRef] [Green Version]
- Takahara, K.; Yashima, Y.; Omatsu, Y.; Yoshida, H.; Kimura, Y.; Kang, Y.-S.; Steinman, R.M.; Gyu, C.P.; Inaba, K. Functional comparison of the mouse DC-SIGN, SIGNR1, SIGNR3 and Langerin, C-type lectins. Int. Immunol. 2004, 16, 819–829. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Smidt, H.; de Vos, H.M.; Bruijns, S.C.; Singh, S.K.; Valence, F.; Molle, D.; Lortal, S.; Altermann, E.; Klaenhammer, T.R.; et al. S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc. Natl. Acad. Sci. USA 2008, 105, 19474–19479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, M.G.; Prado, A.M.; Candurra, N.A.; Ruzal, S.M. S-layer proteins of Lactobacillus acidophilus inhibits JUNV infection. Biochem. Biophys. Res. Commun. 2012, 422, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Malamud, M.; Cavallero, G.J.; Casabuono, A.C.; Lepenies, B.; Serradell, M.L.A.; Couto, A.S. Immunostimulation by Lactobacillus kefiri S-layer proteins with distinct glycosylation patterns requires different lectin partners. J. Biol. Chem. 2020, 295, 14430–14444. [Google Scholar] [CrossRef] [PubMed]
- Prado Acosta, M.; Goyette-Desjardins, G.; Scheffel, J.; Dudeck, A.; Ruland, J.; Lepenies, B.A. S-Layer from Lactobacillus brevis modulates antigen-presenting cell functions via the Mincle-Syk-Card9 axis. Front. Immunol. 2021, 12, 602067. [Google Scholar] [CrossRef] [PubMed]
- Hynönen, U.; Palva, A. Lactobacillus surface layer proteins: Structure, function and applications. Appl. Microbiol. Biotechnol. 2013, 97, 5225–5243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scolari, G.; Vescovo, M.; Zacconi, C.; Vescovi, F. Extraction and partial characterization of proteolytic activities from the cell surface of Lactobacillus helveticus Zuc2. J. Dairy Sci. 2006, 89, 3800–3809. [Google Scholar] [CrossRef] [Green Version]
- Lemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Jin, C.; Wu, L.; Li, J.; Fang, M.; Cheng, L.; Wu, N. Multiple signaling pathways are involved in the interleukine-4 regulated expression of DC-SIGN in THP-1 cell line. J. Biomed. Biotechnol. 2012, 2012, 357060. [Google Scholar] [CrossRef] [Green Version]
- Puig-Kröger, A.; Serrano-Gómez, D.; Caparrós, E.; Domínguez-Soto, A.; Relloso, M.; Colmenares, M.; Martínez-Muñoz, L.; Longo, N.; Sánchez-Sánchez, N.; Rincon, M.; et al. Regulated expression of the pathogen receptor dendritic cell-specific intercellular adhesion molecule 3 (ICAM-3)-grabbing nonintegrin in THP-1 human leukemic cells, monocytes, and macrophages. J. Biol. Chem. 2004, 279, 25680–25688. [Google Scholar] [CrossRef] [Green Version]
- Prado Acosta, M.; Sandra, M.R.; Sandra, M.C. S-layer proteins from Lactobacillus sp. inhibit bacterial infection by blockage of DC-SIGN cell receptor. Int. J. Biol. Macromol. 2016, 92, 998–1005. [Google Scholar] [CrossRef]
- Lee, S.; Lee, H.C.; Kwon, Y.W.; Lee, S.E.; Cho, Y.; Kim, J.; Lee, S.; Kim, J.Y.; Lee, J.; Yang, H.M.; et al. Adenylyl cyclase-associated protein 1 is a receptor for human resistin and mediates inflammatory actions of human monocytes. Cell Metab. 2014, 19, 484–497. [Google Scholar] [CrossRef] [Green Version]
- Saba, L.A.; McComb, M.E.; Potts, D.L.; Costello, C.E.; Amar, S. Proteomic mapping of stimulus-specific signaling pathways involved in THP-1 cells exposed to Porphyromonas gingivalis or its purified components. J. Proteome Res. 2007, 6, 2211. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.T.; Sharma, V.; Elmén, L.; Peterson, S.N. Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota. Clin. Exp. Immunol. 2015, 179, 363–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Zhang, J. Role of intestinal microbiota and metabolites on gut homeostasis and human diseases. BMC Immunol. 2017, 18, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gad, M.; Ravn, P.; Søborg, D.A.; Lund-Jensen, K.; Ouwehand, A.C.; Jensen, S.S. Regulation of the IL-10/IL-12 axis in human dendritic cells with probiotic bacteria. FEMS Immunol. Med. Microbiol. 2011, 3, 93–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malamud, M.; Carasi, P.; Assandri, M.H.; Freire, T.; Lepenies, B.; Serradell, M.L.Á. S-layer glycoprotein from Lactobacillus kefiri exerts its immunostimulatory activity through glycan recognition by Mincle. Front. Immunol. 2019, 26, 1422. [Google Scholar] [CrossRef]
- Hatakeyama, T.; Kamiya, T.; Kusunoki, M.; Nakamura-Tsuruta, S.; Hirabayashi, J.; Goda, S.; Unno, H. Galactose recognition by a tetrameric C-type lectin, CEL-IV, containing the EPN carbohydrate recognition motif. J. Biol. Chem. 2011, 286, 10305–10315. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.T.; Hsu, T.-L.; Ku, S.; Hsieh, S.-L.; Wang, C.-H.; Lee, Y.C. Survey of immune-related, mannose/fucose-binding C-type lectin receptors reveals widely divergent sugar-binding specificities. Glycobiology 2011, 21, 512–520. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Temperature (°C) | Origin |
|---|---|---|
| Lactobacillus helveticus JCM 1120 | 37 | Cheese |
| Lactobacillus acidophilus JCM 1132 | 37 | Human feces |
| Lactobacillus amylovorus JCM 1126 | 37 | Cattle waste |
| Lacticaseibacillus rhamnosus JCM 1136 | 37 | Unknown |
| Lactobacillus delbrueckii subsp. bulgaricus JCM 1002 | 37 | Bulgarian yogurt |
| Levilactobacillus brevis JCM 1059 | 30 | Human feces |
| Lactiplantibacillus plantarum JCM 1100 | 30 | Unknown |
| Lentilactobacillus kefiri JCM 5818 | 30 | Kefir grains |
| Lacticaseibacillus paracasei subsp. paracasei JCM 8130 | 30 | Milk product |
| Lactiplantibacillus plantarum subsp. plantarum JCM 1149 | 30 | Pickled cabbage |
| Lactococcus lactis subsp. cremoris NZ9000 | 30 | Mutant of L. lactis MG1363 |
| Lactococcus lactis subsp. lactis IL1403 | 30 | Cheese |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, T.; Zhang, X.; Iwatani, S.; Miyanaga, K.; Yamamoto, N. Uptake of Levilactobacillus brevis JCM 1059 by THP-1 Cells via Interaction between SlpB and CAP-1 Promotes Cytokine Production. Microorganisms 2023, 11, 247. https://doi.org/10.3390/microorganisms11020247
Yin T, Zhang X, Iwatani S, Miyanaga K, Yamamoto N. Uptake of Levilactobacillus brevis JCM 1059 by THP-1 Cells via Interaction between SlpB and CAP-1 Promotes Cytokine Production. Microorganisms. 2023; 11(2):247. https://doi.org/10.3390/microorganisms11020247
Chicago/Turabian StyleYin, Tingyu, Xiaoxi Zhang, Shun Iwatani, Kazuhiko Miyanaga, and Naoyuki Yamamoto. 2023. "Uptake of Levilactobacillus brevis JCM 1059 by THP-1 Cells via Interaction between SlpB and CAP-1 Promotes Cytokine Production" Microorganisms 11, no. 2: 247. https://doi.org/10.3390/microorganisms11020247
APA StyleYin, T., Zhang, X., Iwatani, S., Miyanaga, K., & Yamamoto, N. (2023). Uptake of Levilactobacillus brevis JCM 1059 by THP-1 Cells via Interaction between SlpB and CAP-1 Promotes Cytokine Production. Microorganisms, 11(2), 247. https://doi.org/10.3390/microorganisms11020247