
Plasmid Composition, Antimicrobial Resistance and Virulence Genes Profiles of Ciprofloxacin- and Third-Generation Cephalosporin-Resistant Foodborne Salmonella enterica Isolates from Russia

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Species Identification, and Antibiotic Susceptibility Detection
2.2. DNA Isolation and Whole Genome Sequencing
2.3. Analysis of Sequencing Data and Genome Assembly
3. Results
3.1. Typing and Classification
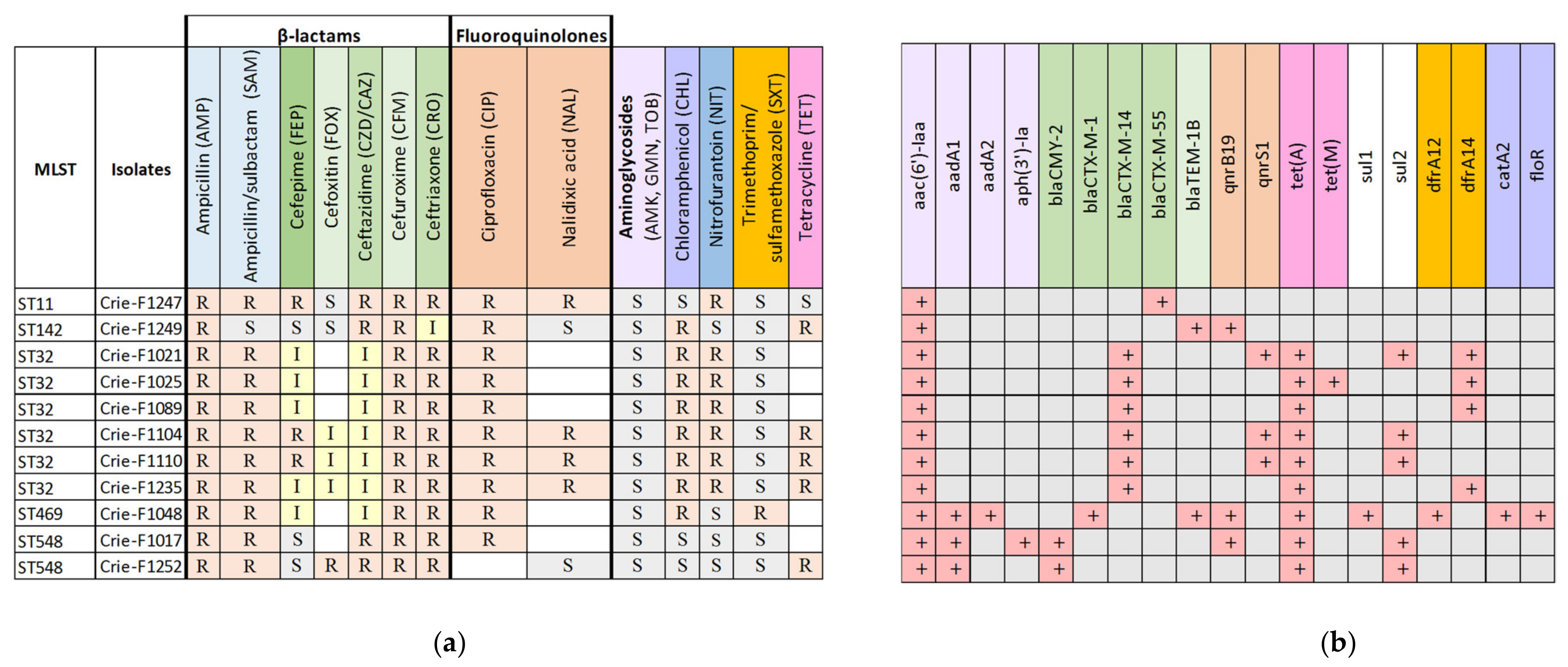
3.2. Antibiotic Resistance Determination
3.3. Virulence Genes
3.4. Plasmids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529–563. [Google Scholar] [CrossRef]
- Pires, S.M.; Desta, B.N.; Mughini-Gras, L.; Mmbaga, B.T.; Fayemi, O.E.; Salvador, E.M.; Gobena, T.; Majowicz, S.E.; Hald, T.; Hoejskov, P.S.; et al. Burden of foodborne diseases: Think global, act local. Curr. Opin. Food Sci. 2021, 39, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Odeyemi, O.A. Public health implications of microbial food safety and foodborne diseases in developing countries. Food Nutr. Res. 2016, 60, 29819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirone, M.; Visciano, P. Trends of Major Foodborne Outbreaks in the European Union during the Years 2015–2019. Hygiene 2021, 1, 106–119. [Google Scholar] [CrossRef]
- Li, X.; Singh, N.; Havelaar, A.H.; Blackburn, J.K. Geographical distribution and space-time clustering of human illnesses with major Salmonella serotypes in Florida, USA, 2017–2018. Epidemiol. Infect. 2022, 150, e175. [Google Scholar] [CrossRef] [PubMed]
- Rakitin, A.L.; Yushina, Y.K.; Zaiko, E.V.; Bataeva, D.S.; Kuznetsova, O.A.; Semenova, A.A.; Ermolaeva, S.A.; Beletskiy, A.V.; Kolganova, T.V.; Mardanov, A.V.; et al. Evaluation of Antibiotic Resistance of Salmonella Serotypes and Whole-Genome Sequencing of Multiresistant Strains Isolated from Food Products in Russia. Antibiotics 2021, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Alba, P.; Leekitcharoenphon, P.; Carfora, V.; Amoruso, R.; Cordaro, G.; Di Matteo, P.; Ianzano, A.; Iurescia, M.; Diaconu, E.L.; Study Group, E.N.; et al. Molecular epidemiology of Salmonella Infantis in Europe: Insights into the success of the bacterial host and its parasitic pESI-like megaplasmid. Microb. Genom. 2020, 6, e000365. [Google Scholar] [CrossRef] [PubMed]
- Arthur, T.M.; Brichta-Harhay, D.M.; Bosilevac, J.M.; Guerini, M.N.; Kalchayanand, N.; Wells, J.E.; Shackelford, S.D.; Wheeler, T.L.; Koohmaraie, M. Prevalence and characterization of Salmonella in bovine lymph nodes potentially destined for use in ground beef. J. Food Prot. 2008, 71, 1685–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechesso, A.F.; Moon, D.C.; Kim, S.J.; Song, H.J.; Kang, H.Y.; Na, S.H.; Choi, J.H.; Kim, H.Y.; Yoon, S.S.; Lim, S.K. Nationwide surveillance on serotype distribution and antimicrobial resistance profiles of non-typhoidal Salmonella serovars isolated from food-producing animals in South Korea. Int. J. Food Microbiol. 2020, 335, 108893. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Antimicrobial Resistance Surveillance System (GLASS): Molecular Methods for Antimicrobial Resistance (AMR) Diagnostics to Enhance the Global Antimicrobial Resistance Surveillance System; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Medalla, F.; Gu, W.; Mahon, B.E.; Judd, M.; Folster, J.; Griffin, P.M.; Hoekstra, R.M. Estimated Incidence of Antimicrobial Drug-Resistant Nontyphoidal Salmonella Infections, United States, 2004–2012. Emerg. Infect. Dis. 2016, 23, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Fontana, J.; Crowe, C.; Bolstorff, B.; Stout, A.; Van Duyne, S.; Hoekstra, M.P.; Whichard, J.M.; Barrett, T.J.; Angulo, F.J.; et al. Emergence of multidrug-resistant Salmonella enterica serotype Newport infections resistant to expanded-spectrum cephalosporins in the United States. J. Infect. Dis. 2003, 188, 1707–1716. [Google Scholar] [CrossRef] [Green Version]
- Weill, F.X.; Demartin, M.; Fabre, L.; Grimont, P.A. Extended-spectrum-beta-lactamase (TEM-52)-producing strains of Salmonella enterica of various serotypes isolated in France. J. Clin. Microbiol. 2004, 42, 3359–3362. [Google Scholar] [CrossRef] [Green Version]
- Miriagou, V.; Tassios, P.T.; Legakis, N.J.; Tzouvelekis, L.S. Expanded-spectrum cephalosporin resistance in non-typhoid Salmonella. Int. J. Antimicrob. Agents 2004, 23, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Ngoi, S.T.; Teh, C.S.; Chai, L.C.; Thong, K.L. Overview of Molecular Typing Tools for The Characterization of Salmonella enterica in Malaysia. Biomed. Environ. Sci. 2015, 28, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Tate, H.; Folster, J.P.; Hsu, C.H.; Chen, J.; Hoffmann, M.; Li, C.; Morales, C.; Tyson, G.H.; Mukherjee, S.; Brown, A.C.; et al. Comparative Analysis of Extended-Spectrum-beta-Lactamase CTX-M-65-Producing Salmonella enterica Serovar Infantis Isolates from Humans, Food Animals, and Retail Chickens in the United States. Antimicrob. Agents Chemother. 2017, 61, e00488-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hindermann, D.; Gopinath, G.; Chase, H.; Negrete, F.; Althaus, D.; Zurfluh, K.; Tall, B.D.; Stephan, R.; Nuesch-Inderbinen, M. Salmonella enterica serovar Infantis from Food and Human Infections, Switzerland, 2010–2015: Poultry-Related Multidrug Resistant Clones and an Emerging ESBL Producing Clonal Lineage. Front. Microbiol. 2017, 8, 1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogomazova, A.N.; Gordeeva, V.D.; Krylova, E.V.; Soltynskaya, I.V.; Davydova, E.E.; Ivanova, O.E.; Komarov, A.A. Mega-plasmid found worldwide confers multiple antimicrobial resistance in Salmonella Infantis of broiler origin in Russia. Int. J. Food Microbiol. 2020, 319, 108497. [Google Scholar] [CrossRef]
- Egorova, A.; Mikhaylova, Y.; Saenko, S.; Tyumentseva, M.; Tyumentsev, A.; Karbyshev, K.; Chernyshkov, A.; Manzeniuk, I.; Akimkin, V.; Shelenkov, A. Comparative Whole-Genome Analysis of Russian Foodborne Multidrug-Resistant Salmonella Infantis Isolates. Microorganisms 2021, 10, 89. [Google Scholar] [CrossRef]
- Vazquez, X.; Fernandez, J.; Rodriguez-Lozano, J.; Calvo, J.; Rodicio, R.; Rodicio, M.R. Genomic Analysis of Two MDR Isolates of Salmonella enterica Serovar Infantis from a Spanish Hospital Bearing the bla(CTX-M-65) Gene with or without fosA3 in pESI-like Plasmids. Antibiotics 2022, 11, 786. [Google Scholar] [CrossRef]
- Yan, S.; Zhang, W.; Li, C.; Liu, X.; Zhu, L.; Chen, L.; Yang, B. Serotyping, MLST, and Core Genome MLST Analysis of Salmonella enterica From Different Sources in China During 2004–2019. Front. Microbiol. 2021, 12, 688614. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelenkov, A.; Mikhaylova, Y.; Yanushevich, Y.; Samoilov, A.; Petrova, L.; Fomina, V.; Gusarov, V.; Zamyatin, M.; Shagin, D.; Akimkin, V. Molecular Typing, Characterization of Antimicrobial Resistance, Virulence Profiling and Analysis of Whole-Genome Sequence of Clinical Klebsiella pneumoniae Isolates. Antibiotics 2020, 9, 261. [Google Scholar] [CrossRef]
- Tyumentseva, M.; Mikhaylova, Y.; Prelovskaya, A.; Tyumentsev, A.; Petrova, L.; Fomina, V.; Zamyatin, M.; Shelenkov, A.; Akimkin, V. Genomic and Phenotypic Analysis of Multidrug-Resistant Acinetobacter baumannii Clinical Isolates Carrying Different Types of CRISPR/Cas Systems. Pathogens 2021, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Feijao, P.; Yao, H.T.; Fornika, D.; Gardy, J.; Hsiao, W.; Chauve, C.; Chindelevitch, L. MentaLiST—A fast MLST caller for large MLST schemes. Microb. Genom. 2018, 4, e000146. [Google Scholar] [CrossRef]
- Nascimento, M.; Sousa, A.; Ramirez, M.; Francisco, A.P.; Carrico, J.A.; Vaz, C. PHYLOViZ 2.0: Providing scalable data integration and visualization for multiple phylogenetic inference methods. Bioinformatics 2017, 33, 128–129. [Google Scholar] [CrossRef] [Green Version]
- Ling, T.K.; Tam, P.C.; Liu, Z.K.; Cheng, A.F. Evaluation of VITEK 2 rapid identification and susceptibility testing system against gram-negative clinical isolates. J. Clin. Microbiol. 2001, 39, 2964–2966. [Google Scholar] [CrossRef] [Green Version]
- Langridge, G.C.; Fookes, M.; Connor, T.R.; Feltwell, T.; Feasey, N.; Parsons, B.N.; Seth-Smith, H.M.; Barquist, L.; Stedman, A.; Humphrey, T.; et al. Patterns of genome evolution that have accompanied host adaptation in Salmonella. Proc. Natl. Acad. Sci. USA 2015, 112, 863–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Olsen, R.H.; Xiao, J.; Meng, H.; Peng, S.; Shi, L. Genetic context of bla (CTX-M-55) and qnrS1 genes in a foodborne Salmonella enterica serotype Saintpaul isolate from China. Front. Microbiol. 2022, 13, 899062. [Google Scholar] [CrossRef] [PubMed]
- Phan, M.D.; Kidgell, C.; Nair, S.; Holt, K.E.; Turner, A.K.; Hinds, J.; Butcher, P.; Cooke, F.J.; Thomson, N.R.; Titball, R.; et al. Variation in Salmonella enterica serovar typhi IncHI1 plasmids during the global spread of resistant typhoid fever. Antimicrob. Agents Chemother. 2009, 53, 716–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Hassena, A.; Haendiges, J.; Zormati, S.; Guermazi, S.; Gdoura, R.; Gonzalez-Escalona, N.; Siala, M. Virulence and resistance genes profiles and clonal relationships of non-typhoidal food-borne Salmonella strains isolated in Tunisia by whole genome sequencing. Int. J. Food Microbiol. 2021, 337, 108941. [Google Scholar] [CrossRef]
- Lartigue, M.F.; Leflon-Guibout, V.; Poirel, L.; Nordmann, P.; Nicolas-Chanoine, M.H. Promoters P3, Pa/Pb, P4, and P5 upstream from bla(TEM) genes and their relationship to beta-lactam resistance. Antimicrob. Agents Chemother. 2002, 46, 4035–4037. [Google Scholar] [CrossRef] [Green Version]
- Devanga Ragupathi, N.K.; Muthuirulandi Sethuvel, D.P.; Shankar, B.A.; Munusamy, E.; Anandan, S.; Veeraraghavan, B. Draft genome sequence of bla(TEM-1)-mediated cephalosporin-resistant Salmonella enterica serovar Typhi from bloodstream infection. J. Glob. Antimicrob. Resist. 2016, 7, 11–12. [Google Scholar] [CrossRef]
- Foley, S.L.; Kaldhone, P.R.; Ricke, S.C.; Han, J. Incompatibility Group I1 (IncI1) Plasmids: Their Genetics, Biology, and Public Health Relevance. Microbiol. Mol. Biol. Rev. 2021, 85. [Google Scholar] [CrossRef]
- Thomas, C.M. Plasmid Incompatibility. In Molecular Life Sciences: An Encyclopedic Reference; Bell, E., Ed.; Springer: New York, NY, USA, 2021; pp. 1–3. [Google Scholar]
- Orlek, A.; Phan, H.; Sheppard, A.E.; Doumith, M.; Ellington, M.; Peto, T.; Crook, D.; Walker, A.S.; Woodford, N.; Anjum, M.F.; et al. Ordering the mob: Insights into replicon and MOB typing schemes from analysis of a curated dataset of publicly available plasmids. Plasmid 2017, 91, 42–52. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Sakano, C.; Kuroda, M.; Sekizuka, T.; Ishioka, T.; Morita, Y.; Ryo, A.; Tsukagoshi, H.; Kawai, Y.; Inoue, N.; Takada, H.; et al. Genetic analysis of non-hydrogen sulfide-producing Salmonella enterica serovar typhimurium and S. enterica serovar infantis isolates in Japan. J. Clin. Microbiol. 2013, 51, 328–330. [Google Scholar] [CrossRef] [Green Version]
- Elkenany, R.; Elsayed, M.M.; Zakaria, A.I.; El-Sayed, S.A.; Rizk, M.A. Antimicrobial resistance profiles and virulence genotyping of Salmonella enterica serovars recovered from broiler chickens and chicken carcasses in Egypt. BMC Vet. Res. 2019, 15, 124. [Google Scholar] [CrossRef] [Green Version]
- Campos, J.; Mourao, J.; Silveira, L.; Saraiva, M.; Correia, C.B.; Macas, A.P.; Peixe, L.; Antunes, P. Imported poultry meat as a source of extended-spectrum cephalosporin-resistant CMY-2-producing Salmonella Heidelberg and Salmonella Minnesota in the European Union, 2014–2015. Int. J. Antimicrob. Agents 2018, 51, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Silveira, L.; Nunes, A.; Pista, A.; Isidro, J.; Belo Correia, C.; Saraiva, M.; Batista, R.; Castanheira, I.; Machado, J.; Gomes, J.P. Characterization of Multidrug-Resistant Isolates of Salmonella enterica Serovars Heidelberg and Minnesota from Fresh Poultry Meat Imported to Portugal. Microb. Drug Resist. 2021, 27, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Monte, D.F.; Lincopan, N.; Berman, H.; Cerdeira, L.; Keelara, S.; Thakur, S.; Fedorka-Cray, P.J.; Landgraf, M. Genomic Features of High-Priority Salmonella enterica Serovars Circulating in the Food Production Chain, Brazil, 2000–2016. Sci. Rep. 2019, 9, 11058. [Google Scholar] [CrossRef] [Green Version]
- Habib, I.; Elbediwi, M.; Ghazawi, A.; Mohamed, M.I.; Lakshmi, G.B.; Khan, M. First report from supermarket chicken meat and genomic characterization of colistin resistance mediated by mcr-1.1 in ESBL-producing, multidrug-resistant Salmonella Minnesota. Int. J. Food Microbiol. 2022, 379, 109835. [Google Scholar] [CrossRef]
- Bertrand, S.; Weill, F.X.; Cloeckaert, A.; Vrints, M.; Mairiaux, E.; Praud, K.; Dierick, K.; Wildemauve, C.; Godard, C.; Butaye, P.; et al. Clonal emergence of extended-spectrum beta-lactamase (CTX-M-2)-producing Salmonella enterica serovar Virchow isolates with reduced susceptibilities to ciprofloxacin among poultry and humans in Belgium and France (2000 to 2003). J. Clin. Microbiol. 2006, 44, 2897–2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weill, F.X.; Lailler, R.; Praud, K.; Kerouanton, A.; Fabre, L.; Brisabois, A.; Grimont, P.A.; Cloeckaert, A. Emergence of extended-spectrum-beta-lactamase (CTX-M-9)-producing multiresistant strains of Salmonella enterica serotype Virchow in poultry and humans in France. J. Clin. Microbiol. 2004, 42, 5767–5773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kameyama, M.; Chuma, T.; Yokoi, T.; Yabata, J.; Tominaga, K.; Miyasako, D.; Iwata, H.; Okamoto, K. Emergence of Salmonella enterica serovar infantis harboring IncI1 plasmid with bla(CTX-M-14) in a broiler farm in Japan. J. Vet. Med. Sci. 2012, 74, 1213–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penha Filho, R.A.C.; Ferreira, J.C.; Kanashiro, A.M.I.; Berchieri Junior, A.; Darini, A. Emergent multidrug-resistant nontyphoidal Salmonella serovars isolated from poultry in Brazil coharboring bla(CTX-M-2) and qnrB or bla(CMY-2) in large plasmids. Diagn. Microbiol. Infect. Dis. 2019, 95, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Ye, Y.; Lan, P.; Han, X.; Quan, J.; Zhou, M.; Yu, Y.; Jiang, Y. Prevalence and Characteristics of Ceftriaxone-Resistant Salmonella in Children’s Hospital in Hangzhou, China. Front. Microbiol. 2021, 12, 764787. [Google Scholar] [CrossRef]
- Rodrigues, I.B.B.E.; Ferreira, K.F.S.; Silva, R.L.; Machado, S.A.; Nascimento, E.R.; Rodrigues, D.P.; Aquino, M.H.C.; Pereira, V.L.A. Amoxicillin/Clavulanic Acid and Cefotaxime Resistance in Salmonella Minnesota and Salmonella Heidelberg from Broiler Chickens. Poult. Sci. J. 2017, 5, 123–129. [Google Scholar] [CrossRef]
- McDermott, P.F.; Zhao, S.; Tate, H. Antimicrobial Resistance in Nontyphoidal Salmonella. Microbiol. Spectr. 2018, 6, 261–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlan, J.P.R.; Lopes, R.; Stehling, E.G. Multidrug resistance IncC plasmid carrying bla(CMY-97) in Shiga toxin-producing Escherichia coli ST215-H54 of ovine origin. Infect. Genet. Evol. 2021, 93, 104989. [Google Scholar] [CrossRef] [PubMed]
- Nordkvist, E.; Zuidema, T.; Herbes, R.G.; Berendsen, B.J. Occurrence of chloramphenicol in cereal straw in north-western Europe. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 798–803. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, B.; Stolker, L.; de Jong, J.; Nielen, M.; Tserendorj, E.; Sodnomdarjaa, R.; Cannavan, A.; Elliott, C. Evidence of natural occurrence of the banned antibiotic chloramphenicol in herbs and grass. Anal. Bioanal. Chem. 2010, 397, 1955–1963. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, J.K.; Mathur, A.D.; Arora, M.M. Reemergence of Chloramphenicol Sensitivity in Enteric Fever. Med. J. Armed Forces India 2007, 63, 212–214. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, F.; Ruiz, J.; Marco, F.; Towner, K.J.; Vila, J. Increase in incidence of resistance to ampicillin, chloramphenicol and trimethoprim in clinical isolates of Salmonella serotype Typhimurium with investigation of molecular epidemiology and mechanisms of resistance. J. Med. Microbiol. 1999, 48, 367–374. [Google Scholar] [CrossRef]
- Sin, M.; Yoon, S.; Kim, Y.B.; Noh, E.B.; Seo, K.W.; Lee, Y.J. Molecular characteristics of antimicrobial resistance determinants and integrons in Salmonella isolated from chicken meat in Korea. J. Appl. Poult. Res. 2020, 29, 502–514. [Google Scholar] [CrossRef]
- Toro, C.S.; Lobos, S.R.; Calderon, I.; Rodriguez, M.; Mora, G.C. Clinical isolate of a porinless Salmonella typhi resistant to high levels of chloramphenicol. Antimicrob. Agents Chemother. 1990, 34, 1715–1719. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, R.; Qi, C.; Gao, H.; Wei, Q.; Tan, L.; Sun, F. Mechanisms of polymyxin resistance induced by Salmonella typhimurium in vitro. Vet. Microbiol. 2021, 257, 109063. [Google Scholar] [CrossRef]
- Patil, N.; Mule, P. Sensitivity Pattern of Salmonella typhi and Paratyphi A Isolates to Chloramphenicol and other Anti-Typhoid Drugs: An In Vitro Study. Infect. Drug Resist. 2019, 12, 3217–3225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.; Kaur, J.; Kaistha, N. Re-emerging chloramphenicol sensitivity and emerging low level ciprofloxacin resistance among Salmonella enterica serotype typhi isolates in North India. Trop. Doct. 2009, 39, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Lerminiaux, N.A.; MacKenzie, K.D.; Cameron, A.D.S. Salmonella Pathogenicity Island 1 (SPI-1): The Evolution and Stabilization of a Core Genomic Type Three Secretion System. Microorganisms 2020, 8, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurekci, C.; Sahin, S.; Iwan, E.; Kwit, R.; Bomba, A.; Wasyl, D. Whole-genome sequence analysis of Salmonella Infantis isolated from raw chicken meat samples and insights into pESI-like megaplasmid. Int. J. Food Microbiol. 2021, 337, 108956. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Soto, S.; Abdel-Glil, M.Y.; Tomaso, H.; Linde, J.; Methner, U. Emergence of Multidrug-Resistant Salmonella enterica Subspecies enterica Serovar Infantis of Multilocus Sequence Type 2283 in German Broiler Farms. Front. Microbiol. 2020, 11, 1741. [Google Scholar] [CrossRef] [PubMed]
- Shaidullina, E.; Shelenkov, A.; Yanushevich, Y.; Mikhaylova, Y.; Shagin, D.; Alexandrova, I.; Ershova, O.; Akimkin, V.; Kozlov, R.; Edelstein, M. Antimicrobial Resistance and Genomic Characterization of OXA-48- and CTX-M-15-Co-Producing Hypervirulent Klebsiella pneumoniae ST23 Recovered from Nosocomial Outbreak. Antibiotics 2020, 9, 862. [Google Scholar] [CrossRef] [PubMed]
- Andesfha, E.; Indrawati, A.; Mayasari, N.; Rahayuningtyas, I.; Jusa, I. Detection of Salmonella pathogenicity island and Salmonella plasmid virulence genes in Salmonella Enteritidis originated from layer and broiler farms in Java Island. J. Adv. Vet. Anim. Res. 2019, 6, 384–393. [Google Scholar] [CrossRef]
- Mambu, J.; Virlogeux-Payant, I.; Holbert, S.; Grepinet, O.; Velge, P.; Wiedemann, A. An Updated View on the Rck Invasin of Salmonella: Still Much to Discover. Front. Cell. Infect. Microbiol. 2017, 7, 500. [Google Scholar] [CrossRef]
- Karacan Sever, N.; Akan, M. Molecular analysis of virulence genes of Salmonella Infantis isolated from chickens and turkeys. Microb. Pathog. 2019, 126, 199–204. [Google Scholar] [CrossRef]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Plasmids in Gram negatives: Molecular typing of resistance plasmids. Int. J. Med. Microbiol. 2011, 301, 654–658. [Google Scholar] [CrossRef]
- Gilmour, M.W.; Thomson, N.R.; Sanders, M.; Parkhill, J.; Taylor, D.E. The complete nucleotide sequence of the resistance plasmid R478: Defining the backbone components of incompatibility group H conjugative plasmids through comparative genomics. Plasmid 2004, 52, 182–202. [Google Scholar] [CrossRef]
- Holt, K.E.; Thomson, N.R.; Wain, J.; Phan, M.D.; Nair, S.; Hasan, R.; Bhutta, Z.A.; Quail, M.A.; Norbertczak, H.; Walker, D.; et al. Multidrug-resistant Salmonella enterica serovar paratyphi A harbors IncHI1 plasmids similar to those found in serovar typhi. J. Bacteriol. 2007, 189, 4257–4264. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Fang, T.; Zhou, X.; Zhang, D.; Shi, X.; Shi, C. IncHI2 Plasmids Are Predominant in Antibiotic-Resistant Salmonella Isolates. Front. Microbiol. 2016, 7, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillan, E.A.; Jackson, C.R.; Frye, J.G. Transferable Plasmids of Salmonella enterica Associated with Antibiotic Resistance Genes. Front. Microbiol. 2020, 11, 562181. [Google Scholar] [CrossRef] [PubMed]
- Harmer, C.J.; Hall, R.M. The A to Z of A/C plasmids. Plasmid 2015, 80, 63–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Lynne, A.M.; David, D.E.; Tang, H.; Xu, J.; Nayak, R.; Kaldhone, P.; Logue, C.M.; Foley, S.L. DNA sequence analysis of plasmids from multidrug resistant Salmonella enterica serotype Heidelberg isolates. PLoS ONE 2012, 7, e51160. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MLST | Isolates | Total Number of Genes Found | Virulence Genes | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| allB | cdtB | entA | fepC | fyuA/psn; irp1,2; ybt | lpfA-E | mig-5 | pefA-D | pipB2 | pltAB | ratB | rck | shdA | sodCI | spvBCD | sseI/srfH | sseK1 | sseK2 | sspH2 | tae4 | tlde1 | tssM | |||
| ST11 | Crie-F1247 | 140 | + | − | + | − | − | + | + | + | + | − | + | + | − | + | + | + | + | − | + | − | + | + |
| ST142 | Crie-F1249 | 130 | + | − | + | + | − | + | − | − | + | − | + | − | − | + | − | + | + | + | + | − | − | − |
| ST32 | Crie-F1021 | 162 | + | − | − | + | + | + | − | − | + | − | + | − | + | − | − | − | + | + | + | + | + | − |
| ST32 | Crie-F1025 | 162 | + | − | − | + | + | + | − | − | + | − | + | − | + | − | − | − | + | + | + | + | + | − |
| ST32 | Crie-F1089 | 162 | + | − | − | + | + | + | − | − | + | − | + | − | + | − | − | − | + | + | + | + | + | − |
| ST32 | Crie-F1104 | 162 | + | − | − | + | + | + | − | − | + | − | + | − | + | − | − | − | + | + | + | + | + | − |
| ST32 | Crie-F1110 | 162 | + | − | − | + | + | + | − | − | + | − | + | − | + | − | − | − | + | + | + | + | + | − |
| ST32 | Crie-F1235 | 162 | + | − | − | + | + | + | − | − | + | − | + | − | + | − | − | − | + | + | + | + | + | − |
| ST469 | Crie-F1048 | 149 | + | − | + | + | − | + | − | − | + | − | − | − | − | − | − | − | + | + | − | + | + | − |
| ST548 | Crie-F1017 | 156 | − | + | − | + | + | − | − | − | − | + | + | − | − | − | − | − | − | + | − | + | + | − |
| ST548 | Crie-F1252 | 156 | − | + | − | + | + | − | − | − | − | + | + | − | − | − | − | − | − | + | − | + | + | − |
| MLST | Isolates | Plasmid Replicons | Predicted Mobility | |||
|---|---|---|---|---|---|---|
| IncF | IncC | Col | Others | |||
| ST11 | Crie-F1247 | IncFIB, IncFII | − | − | IncI1-I | |
| ST142 | Crie-F1249 * | − | − | Col(pHAD28) | IncHI2, IncHI2A | conjugative |
| ST32 | Crie-F1021 | IncFIB | − | − | − | mobilizable |
| ST32 | Crie-F1025 | IncFIB | − | − | − | mobilizable |
| ST32 | Crie-F1089 | IncFIB | − | − | − | non-mobilizable |
| ST32 | Crie-F1104 | IncFIB | − | − | − | |
| ST32 | Crie-F1110 * | IncFIB | − | − | − | conjugative |
| ST32 | Crie-F1235 | IncFIB | − | − | − | conjugative |
| ST469 | Crie-F1048 | − | − | Col(pHAD28) | IncI1-I | non-mobilizable |
| ST548 | Crie-F1017 * | − | IncC | Col(pHAD28) | − | conjugative |
| ST548 | Crie-F1252 * | − | IncC | − | − | conjugative |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egorova, A.; Shelenkov, A.; Kuleshov, K.; Kulikova, N.; Chernyshkov, A.; Manzeniuk, I.; Mikhaylova, Y.; Akimkin, V. Plasmid Composition, Antimicrobial Resistance and Virulence Genes Profiles of Ciprofloxacin- and Third-Generation Cephalosporin-Resistant Foodborne Salmonella enterica Isolates from Russia. Microorganisms 2023, 11, 347. https://doi.org/10.3390/microorganisms11020347
Egorova A, Shelenkov A, Kuleshov K, Kulikova N, Chernyshkov A, Manzeniuk I, Mikhaylova Y, Akimkin V. Plasmid Composition, Antimicrobial Resistance and Virulence Genes Profiles of Ciprofloxacin- and Third-Generation Cephalosporin-Resistant Foodborne Salmonella enterica Isolates from Russia. Microorganisms. 2023; 11(2):347. https://doi.org/10.3390/microorganisms11020347
Chicago/Turabian StyleEgorova, Anna, Andrey Shelenkov, Konstantin Kuleshov, Nina Kulikova, Aleksey Chernyshkov, Igor Manzeniuk, Yulia Mikhaylova, and Vasiliy Akimkin. 2023. "Plasmid Composition, Antimicrobial Resistance and Virulence Genes Profiles of Ciprofloxacin- and Third-Generation Cephalosporin-Resistant Foodborne Salmonella enterica Isolates from Russia" Microorganisms 11, no. 2: 347. https://doi.org/10.3390/microorganisms11020347
APA StyleEgorova, A., Shelenkov, A., Kuleshov, K., Kulikova, N., Chernyshkov, A., Manzeniuk, I., Mikhaylova, Y., & Akimkin, V. (2023). Plasmid Composition, Antimicrobial Resistance and Virulence Genes Profiles of Ciprofloxacin- and Third-Generation Cephalosporin-Resistant Foodborne Salmonella enterica Isolates from Russia. Microorganisms, 11(2), 347. https://doi.org/10.3390/microorganisms11020347