
Whole-Genome Sequencing-Based Resistome Analysis of Nosocomial Multidrug-Resistant Non-Fermenting Gram-Negative Pathogens from the Balkans


Abstract
:1. Introduction
2. Antimicrobial Resistance in NFGNBs in the Balkan States
3. Whole-Genome Sequencing of Bacterial Pathogens
4. P. aeruginosa
4.1. Mechanisms of Carbapenem Resistance in P. aeruginosa
4.2. WGS of Clinical P. aeruginosa Isolates in Albania
4.3. WGS of Clinical P. aeruginosa Isolates in Bulgaria
4.4. WGS of Clinical P. aeruginosa Isolates in Greece
4.5. WGS of Clinical P. aeruginosa Isolates in Romania
4.6. WGS of Clinical P. aeruginosa Isolates in Serbia
4.7. WGS of Clinical P. aeruginosa Isolates in Turkey
4.8. WGS of Clinical P. aeruginosa Isolates from the Balkans in International Projects
5. A. baumannii
5.1. Mechanisms of Carbapenem Resistance in A. baumannii
5.2. Mechanisms of Colistin Resistance in A. baumannii
5.3. WGS of Clinical A. baumannii Isolates in Albania
5.4. WGS of Clinical A. baumannii Isolates in Croatia
5.5. WGS of Clinical A. baumannii Isolates in Greece
5.6. WGS of Clinical A. baumannii Isolates in Romania
5.7. WGS of Clinical A. baumannii Isolates in Serbia
5.8. WGS of Clinical A. baumannii Isolates in Turkey
5.9. WGS of Clinical A. baumannii Isolates from the Balkans in International Projects
6. S. maltophilia
6.1. Mechanisms of Antibiotic Resistance in S. maltophilia
6.2. WGS of Clinical S. maltophilia Isolates in Bulgaria
7. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rezaei, N. Encyclopedia of Infection and Immunity; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Wisplinghoff, H. Pseudomonas spp., Acinetobacter spp. and Miscellaneous Gram-Negative Bacilli; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1579–1599.e2. [Google Scholar] [CrossRef]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial resistance in ESKAPE pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/en/news-room/detail/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 19 January 2023).
- Asokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO Global Priority Pathogens List: A Bibliometric Analysis of Medline-PubMed for Knowledge Mobilization to Infection Prevention and Control Practices in Bahrain. Oman Med. J. 2019, 34, 184. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States. 2019. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html (accessed on 10 January 2023).
- Kadri, S.S. Key Takeaways from the U.S. CDC’s 2019 Antibiotic Resistance Threats Report for Frontline Providers. Crit. Care Med. 2020, 48, 939–945. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet. Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Chumbita, M.; Monzo-Gallo, P.; Lopera-Mármol, C.; Aiello, T.F.; Puerta-Alcalde, P.; Garcia-Vidal, C. New treatments for multidrug-resistant non-fermenting Gram-negative bacilli Infections. Rev. Española Quimioter. 2022, 35, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Soriano, M.C.; Montufar, J.; Blandino-Ortiz, A. Cefiderocol. Rev. Española Quimioter. 2022, 35 (Suppl. S1), 31. [Google Scholar] [CrossRef]
- Syed, Y.Y. Cefiderocol: A Review in Serious Gram-Negative Bacterial Infections. Drugs 2021, 81, 1559–1571. [Google Scholar] [CrossRef]
- El-Sayed Ahmed, M.A.E.G.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and its role in the Era of antibiotic resistance: An extended review(2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [Green Version]
- Paterson, D.L.; Harris, P.N.A. Colistin resistance: A major breach in our last line of defence. Lancet Infect. Dis. 2016, 16, 132–133. [Google Scholar] [CrossRef]
- Hasannejad-Bibalan, M.; Jafari, A.; Sabati, H.; Goswami, R.; Jafaryparvar, Z.; Sedaghat, F.; Sedigh Ebrahim-Saraie, H. Risk of type III secretion systems in burn patients with Pseudomonas aeruginosa wound infection: A systematic review and meta-analysis. Burns 2021, 47, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Mitov, I.; Strateva, T.; Markova, B. Prevalence of Virulence Genes Among Bulgarian Nosocomial and Cystic Fibrosis Isolates of Pseudomonas aeruginosa. Braz. J. Microbiol. 2010, 41, 588–595. [Google Scholar] [CrossRef]
- Yakout, M.A.; Abdelwahab, I.A. Diabetic Foot Ulcer Infections and Pseudomonas aeruginosa Biofilm Production during the Covid-19 Pandemic. J. Pure Appl. Microbiol. 2022, 16, 138–146. [Google Scholar] [CrossRef]
- Bhagirath, A.Y.; Li, Y.; Somayajula, D.; Dadashi, M.; Badr, S.; Duan, K. Cystic fibrosis lung environment and Pseudomonas aeruginosa infection. BMC Pulm. Med. 2016, 16, 174. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Nadal, G.; Puerta-Alcalde, P.; Gudiol, C.; Cardozo, C.; Albasanz-Puig, A.; Marco, F.; Laporte-Amargós, J.; Moreno-García, E.; Domingo-Doménech, E.; Chumbita, M.; et al. Inappropriate Empirical Antibiotic Treatment in High-risk Neutropenic Patients With Bacteremia in the Era of Multidrug Resistance. Clin. Infect. Dis. 2020, 70, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Mestrovic, T.; Robles Aguilar, G.; Swetschinski, L.R.; Ikuta, K.S.; Gray, A.P.; Davis Weaver, N.; Han, C.; Wool, E.E.; Gershberg Hayoon, A.; Hay, S.I.; et al. The burden of bacterial antimicrobial resistance in the WHO European region in 2019: A cross-country systematic analysis. Lancet Public Health 2022, 7, e897–e913. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moubareck, C.A.; Halat, D.H. Insights into Acinetobacter baumannii: A Review of Microbiological, Virulence, and Resistance Traits in a Threatening Nosocomial Pathogen. Antibiotics 2020, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.N.; Burns, T.C.; Hayda, R.A.; Hospenthal, D.R.; Murray, C.K. Infectious complications of open type III tibial fractures among combat casualties. Clin. Infect. Dis. 2007, 45, 409–415. [Google Scholar] [CrossRef]
- Keen, E.F.; Murray, C.K.; Robinson, B.J.; Hospenthal, D.R.; Aldous, W.K. Changes in the incidences of multidrug-resistant and extensively drug-resistant organisms isolated in a military medical center. Infect. Control. Hosp. Epidemiol. 2010, 31, 728–732. [Google Scholar] [CrossRef]
- Looney, W.J.; Narita, M.; Mühlemann, K. Stenotrophomonas maltophilia: An emerging opportunist human pathogen. Lancet Infect. Dis. 2009, 9, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Trifonova, A.; Strateva, T. Stenotrophomonas maltophilia—A low-grade pathogen with numerous virulence factors. Infect. Dis. 2019, 51, 168–178. [Google Scholar] [CrossRef]
- Chawla, K.; Vishwanath, S.; Munim, F.C. Nonfermenting Gram-negative Bacilli other than Pseudomonas aeruginosa and Acinetobacter Spp. Causing Respiratory Tract Infections in a Tertiary Care Center. J. Glob. Infect. Dis. 2013, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- Abbott, I.J.; Slavin, M.A.; Turnidge, J.D.; Thursky, K.A.; Worth, L.J. Stenotrophomonas maltophilia: Emerging disease patterns and challenges for treatment. Expert Rev. Anti-Infect. Ther. 2014, 9, 471–488. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Hua, M.; Liu, X.; Du, C.; Pu, L.; Xiang, P.; Wang, L.; Liu, J. Bacterial and fungal co-infections among COVID-19 patients in intensive care unit. Microbes Infect. 2021, 23, 104806. [Google Scholar] [CrossRef]
- COVID-19 Deaths per Capita by Country|Statista. Available online: https://www.statista.com/statistics/1104709/coronavirus-deaths-worldwide-per-million-inhabitants/ (accessed on 20 January 2023).
- Sánchez, M.B. Antibiotic resistance in the opportunistic pathogen Stenotrophomonas maltophilia. Front. Microbiol. 2015, 6, 658. [Google Scholar] [CrossRef] [Green Version]
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2016, 3, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.T.; Lin, C.Y.; Chen, Y.H.; Hsueh, P.R. Update on infections caused by Stenotrophomonas maltophilia with particular attention to resistance mechanisms and therapeutic options. Front. Microbiol. 2015, 6, 893. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, C.H.; Nastro, M.; Calvo, J.L.; Fariña, M.E.; Dabos, L.; Famiglietti, A. In vitro activity of colistin against Stenotrophomonas maltophilia. J. Glob. Antimicrob. Resist. 2014, 2, 316–317. [Google Scholar] [CrossRef]
- Lee, D.H.; Cha, J.H.; Kim, D.W.; Lee, K.; Kim, Y.S.; Oh, H.Y.; Cho, Y.H.; Cha, C.J. Colistin-degrading proteases confer collective resistance to microbial communities during polymicrobial infections. Microbiome 2022, 10, 129. [Google Scholar] [CrossRef]
- Britannica. Balkans|Definition, Map, Countries, & Facts|Britannica. Available online: https://www.britannica.com/place/Balkans (accessed on 20 January 2023).
- European Centre for Disease Prevention and Control. Antimicrobial resistance surveillance in Europe 2020. Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net); ECDC: Stockholm, Sweden, 2020. [Google Scholar]
- GLASS Whole-Genome Sequencing for Surveillance of Antimicrobial Resistance. Available online: https://www.who.int/publications/i/item/9789240011007 (accessed on 20 January 2023).
- Beg, A.Z.; Khan, A.U. Exploring bacterial resistome and resistance dessemination: An approach of whole genome sequencing. Future Med. Chem. 2019, 11, 247–260. [Google Scholar] [CrossRef]
- Fleischmann, R.D.; Adams, M.D.; White, O.; Clayton, R.A.; Kirkness, E.F.; Kerlavage, A.R.; Bult, C.J.; Tomb, J.F.; Dougherty, B.A.; Merrick, J.M.; et al. Whole-Genome Random Sequencing and Assembly of Haemophilus influenzae Rd. Science 1995, 269, 496–512. [Google Scholar] [CrossRef] [Green Version]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.L.; Hufnagle, W.O.; Kowallk, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossman, L.C.; Gould, V.C.; Dow, J.M.; Vernikos, G.S.; Okazaki, A.; Sebaihia, M.; Saunders, D.; Arrowsmith, C.; Carver, T.; Peters, N.; et al. The complete genome, comparative and functional analysis of Stenotrophomonas maltophilia reveals an organism heavily shielded by drug resistance determinants. Genome Biol. 2008, 9, R74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, P.E.; Vallenet, D.; Barbe, V.; Audic, S.; Ogata, H.; Poirel, L.; Richet, H.; Robert, C.; Mangenot, S.; Abergel, C.; et al. Comparative Genomics of Multidrug Resistance in Acinetobacter baumannii. PLoS Genet. 2006, 2, e7. [Google Scholar] [CrossRef] [Green Version]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenblum, B.B.; Lee, L.G.; Spurgeon, S.L.; Khan, S.H.; Menchen, S.M.; Heiner, C.R.; Chen, S.M. New dye-labeled terminators for improved DNA sequencing patterns. Nucleic Acids Res. 1997, 25, 4500–4504. [Google Scholar] [CrossRef] [Green Version]
- Behjati, S.; Tarpey, P.S. What is next generation sequencing? Arch. Dis. Child.-Educ. Pract. 2013, 98, 236–238. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.G.; Gianoulis, T.A.; Pukatzki, S.; Mekalanos, J.J.; Ornston, L.N.; Gerstein, M.; Snyder, M. New insights into Acinetobacter baumannii pathogenesis revealed by high-density pyrosequencing and transposon mutagenesis. Genes Dev. 2007, 21, 601–614. [Google Scholar] [CrossRef] [Green Version]
- Nadon, C.; van Walle, I.; Gerner-Smidt, P.; Campos, J.; Chinen, I.; Concepcion-Acevedo, J.; Gilpin, B.; Smith, A.M.; Kam, K.M.; Perez, E.; et al. PulseNet International: Vision for the implementation of whole genome sequencing (WGS) for global food-borne disease surveillance. Eurosurveillance 2017, 22, 30544. [Google Scholar] [CrossRef] [Green Version]
- Didelot, X.; Bowden, R.; Wilson, D.J.; Peto, T.E.A.; Crook, D.W. Transforming clinical microbiology with bacterial genome sequencing. Nat. Rev. Genet. 2012, 13, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, M.; D’Andrea, M.M.; Pelegrin, A.C.; Mirande, C.; Brkic, S.; Cirkovic, I.; Goossens, H.; Rossolini, G.M.; van Belkum, A. Genomic Epidemiology of Carbapenem- and Colistin-Resistant Klebsiella pneumoniae Isolates from Serbia: Predominance of ST101 Strains Carrying a Novel OXA-48 Plasmid. Front. Microbiol. 2020, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Strateva, T.; Peykov, S.; Sirakov, I.; Savov, E.; Dimov, S.; Mitov, I. First detection and characterisation of a VanA-type Enterococcus faecalis clinical isolate from Bulgaria. J. Glob. Antimicrob. Resist. 2019, 18, 260–262. [Google Scholar] [CrossRef] [PubMed]
- Peykov, S.; Stratev, A.; Kirov, B.; Gergova, R.; Strateva, T. First detection of a colistin-resistant Klebsiella aerogenes isolate from a critically ill patient with septic shock in Bulgaria. Acta Microbiol. Immunol. Hung. 2022, 69, 209–214. [Google Scholar] [CrossRef]
- Surleac, M.; Barbu, I.C.; Paraschiv, S.; Popa, L.I.; Gheorghe, I.; Marutescu, L.; Popa, M.; Sarbu, I.; Talapan, D.; Nita, M.; et al. Whole genome sequencing snapshot of multi-drug resistant Klebsiella pneumoniae strains from hospitals and receiving wastewater treatment plants in Southern Romania. PLoS ONE 2020, 15, e0228079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turumtay, H.; Allam, M.; Sandalli, A.; Turumtay, E.A.; Genç, H.; Sandalli, C. Characteristics in the whole-genome sequence of Klebsiella pneumoniae ST147 from Turkey. Acta Microbiol. Immunol. Hung. 2022, 69, 144–149. [Google Scholar] [CrossRef]
- Meletis, G.; Chatzopoulou, F.; Chatzidimitriou, D.; Tsingerlioti, F.; Botziori, C.; Tzimagiorgis, G.; Skoura, L. Whole Genome Sequencing of NDM-1-Producing ST11 Klebsiella pneumoniae Isolated in a Private Laboratory in Greece. Microb. Drug Resist. 2019, 25, 80–86. [Google Scholar] [CrossRef]
- Karampatakis, T.; Papadopoulos, P.; Tsergouli, K.; Angelidis, A.S.; Sergelidis, D.; Papa, A. Genetic characterization of two methicillin-resistant Staphylococcus aureus spa type t127 strains isolated from workers in the dairy production chain in Greece. Acta Microbiol. Immunol. Hung. 2021, 68, 189–194. [Google Scholar] [CrossRef]
- Slatko, B.E.; Gardner, A.F.; Ausubel, F.M. Overview of Next Generation Sequencing Technologies. Curr. Protoc. Mol. Biol. 2018, 122, e59. [Google Scholar] [CrossRef]
- Mardis, E.R. Next-generation DNA sequencing methods. Annu. Rev. Genomics Hum. Genet. 2008, 9, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Fuller, C.W.; Middendorf, L.R.; Benner, S.A.; Church, G.M.; Harris, T.; Huang, X.; Jovanovich, S.B.; Nelson, J.R.; Schloss, J.A.; Schwartz, D.C.; et al. The challenges of sequencing by synthesis. Nat. Biotechnol. 2009, 27, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Satola, S.W.; Read, T.D. Genome-Based Prediction of Bacterial Antibiotic Resistance. J. Clin. Microbiol. 2019, 57, e01405-18. [Google Scholar] [CrossRef] [Green Version]
- Help for Assembly. Available online: https://www.ncbi.nlm.nih.gov/assembly/help/ (accessed on 16 January 2023).
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple Alignment of Conserved Genomic Sequence With Rearrangements. Genome Res. 2004, 14, 1394. [Google Scholar] [CrossRef] [Green Version]
- Kolmogorov, M.; Raney, B.; Paten, B.; Pham, S. Ragout-a reference-assisted assembly tool for bacterial genomes. Bioinformatics 2014, 30, i302–i309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolmogorov, M.; Armstrong, J.; Raney, B.J.; Streeter, I.; Dunn, M.; Yang, F.; Odom, D.; Flicek, P.; Keane, T.M.; Thybert, D.; et al. Chromosome assembly of large and complex genomes using multiple references. Genome Res. 2018, 28, 1720–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.C.; Ju, Y.R.; Lu, C.L. Multi-CSAR: A web server for scaffolding contigs using multiple reference genomes. Nucleic Acids Res. 2022, 50, W500–W509. [Google Scholar] [CrossRef] [PubMed]
- Bosi, E.; Donati, B.; Galardini, M.; Brunetti, S.; Sagot, M.F.; Lió, P.; Crescenzi, P.; Fani, R.; Fondi, M. MeDuSa: A multi-draft based scaffolder. Bioinformatics 2015, 31, 2443–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyre, D.W.; Silva, D.; de Cole, K.; Peters, J.; Cole, M.J.; Grad, Y.H.; Demczuk, W.; Martin, I.; Mulvey, M.R.; Crook, D.W.; et al. WGS to predict antibiotic MICs for Neisseria gonorrhoeae. J. Antimicrob. Chemother. 2017, 72, 1937–1947. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, A.; Arnold, C.; Woodford, N. Rapid detection and estimation by pyrosequencing of 23S rRNA genes with a single nucleotide polymorphism conferring linezolid resistance in Enterococci. Antimicrob. Agents Chemother. 2003, 47, 3620–3622. [Google Scholar] [CrossRef] [Green Version]
- Ellington, M.J.; Ekelund, O.; Aarestrup, F.M.; Canton, R.; Doumith, M.; Giske, C.; Grundman, H.; Hasman, H.; Holden, M.T.G.; Hopkins, K.L.; et al. The role of whole genome sequencing in antimicrobial susceptibility testing of bacteria: Report from the EUCAST Subcommittee. Clin. Microbiol. Infect. 2017, 23, 2–22. [Google Scholar] [CrossRef] [Green Version]
- Lauretti, L.; Riccio, M.L.; Mazzariol, A.; Cornaglia, G.; Amicosante, G.; Fontana, R.; Rossolini, G.M. Cloning and Characterization of blaVIM, a New Integron-Borne Metallo-β-Lactamase Gene from a Pseudomonas aeruginosa Clinical Isolate. Antimicrob. Agents Chemother. 1999, 43, 1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strateva, T.; Yordanov, D. Pseudomonas aeruginosa—A phenomenon of bacterial resistance. J. Med. Microbiol. 2009, 58, 1133–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakata, Y.; Yamaguchi, T.; Nakano, M.; Izumikawa, K.; Mine, M.; Aoki, S.; Kondoh, A.; Matsuda, J.; Hirayama, M.; Yanagihara, K.; et al. Clinical and Bacteriological Characteristics of IMP-Type Metallo-β-Lactamase-Producing Pseudomonas aeruginosa. Clin. Infect. Dis. 2003, 37, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, H.; Arakawa, Y.; Ohsuka, S.; Wacharotayankun, R.; Kato, N.; Ohta, M. Plasmid-mediated dissemination of the metallo-beta-lactamase gene blaIMP among clinically isolated strains of Serratia marcescens. Antimicrob. Agents Chemother. 1995, 39, 824–829. [Google Scholar] [CrossRef] [Green Version]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.F.; Chen, G.S.; Kong, Q.X.; Gao, L.P.; Chen, X.; Ye, Y.; Li, J. Bin Increase in the Prevalence of Resistance Determinants to Trimethoprim/Sulfamethoxazole in Clinical Stenotrophomonas maltophilia Isolates in China. PLoS ONE 2016, 11, e0157693. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.H.; Kuo, S.C.; Lee, Y.T.; Chang, I.C.Y.; Yang, S.P.; Chen, T.L.; Fung, C.P. Amino acid substitutions of quinolone resistance determining regions in GyrA and ParC associated with quinolone resistance in Acinetobacter baumannii and Acinetobacter genomic species 13TU. J. Microbiol. Immunol. Infect. 2012, 45, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Zaki, M.E.S.; ElKheir, N.A.; Mofreh, M. Molecular Study of Quinolone Resistance Determining Regions of gyrA Gene and parC Genes in Clinical Isolates of Acintobacter baumannii Resistant to Fluoroquinolone. Open Microbiol. J. 2018, 12, 116. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.T.; Lee, M.F.; Peng, C.F. Mutations in the quinolone resistance-determining regions associated with ciprofloxacin resistance in Pseudomonas aeruginosa isolates from Southern Taiwan. Biomark. Genom. Med. 2014, 6, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, L.A.; Herrera, C.M.; Fernandez, L.; Hankins, J.V.; Trent, M.S.; Hancock, R.E.W. The pmrCAB Operon Mediates Polymyxin Resistance in Acinetobacter baumannii ATCC 17978 and Clinical Isolates through Phosphoethanolamine Modification of Lipid A. Antimicrob. Agents Chemother. 2011, 55, 3743. [Google Scholar] [CrossRef] [Green Version]
- Fernández, L.; Álvarez-Ortega, C.; Wiegand, I.; Olivares, J.; Kocíncová, D.; Lam, J.S.; Martínez, J.L.; Hancock, R.E.W. Characterization of the Polymyxin B Resistome of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 57, 110. [Google Scholar] [CrossRef] [Green Version]
- Gutu, A.D.; Sgambati, N.; Strasbourger, P.; Brannon, M.K.; Jacobs, M.A.; Haugen, E.; Kaul, R.K.; Johansen, H.K.; Høiby, N.; Moskowitz, S.M. Polymyxin resistance of Pseudomonas aeruginosa phoQ mutants is dependent on additional two-component regulatory systems. Antimicrob. Agents Chemother. 2013, 57, 2204–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buermans, H.P.J.; den Dunnen, J.T. Next generation sequencing technology: Advances and applications. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 1932–1941. [Google Scholar] [CrossRef] [Green Version]
- Drmanac, R.; Sparks, A.B.; Callow, M.J.; Halpern, A.L.; Burns, N.L.; Kermani, B.G.; Carnevali, P.; Nazarenko, I.; Nilsen, G.B.; Yeung, G.; et al. Human genome sequencing using unchained base reads on self-assembling DNA nanoarrays. Science 2010, 327, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Korostin, D.; Kulemin, N.; Naumov, V.; Belova, V.; Kwon, D.; Gorbachev, A. Comparative analysis of novel MGISEQ-2000 sequencing platform vs Illumina HiSeq 2500 for whole-genome sequencing. PLoS ONE 2020, 15, e0230301. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Erickson, D.L.; Meng, J. Benchmarking hybrid assembly approaches for genomic analyses of bacterial pathogens using Illumina and Oxford Nanopore sequencing. BMC Genom. 2020, 21, 631. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Jeong, S.H. Mobile Carbapenemase Genes in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 614058. [Google Scholar] [CrossRef]
- Mosquera-Rendón, J.; Rada-Bravo, A.M.; Cárdenas-Brito, S.; Corredor, M.; Restrepo-Pineda, E.; Benítez-Páez, A. Pangenome-wide and molecular evolution analyses of the Pseudomonas aeruginosa species. BMC Genomics 2016, 17, 45. [Google Scholar] [CrossRef] [Green Version]
- Drusano, G.L.; Standiford, H.C. Emergence of resistance to carbapenem antibiotics in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 1989, 24 (Suppl. A), 161–167. [Google Scholar] [CrossRef] [Green Version]
- Köhler, T.; Michea-Hamzehpour, M.; Epp, S.F.; Pechere, J.C. Carbapenem activities against Pseudomonas aeruginosa: Respective contributions of OprD and efflux systems. Antimicrob. Agents Chemother. 1999, 43, 424–427. [Google Scholar] [CrossRef] [Green Version]
- Farra, A.; Islam, S.; Strålfors, A.; Sörberg, M.; Wretlind, B. Role of outer membrane protein OprD and penicillin-binding proteins in resistance of Pseudomonas aeruginosa to imipenem and meropenem. Int. J. Antimicrob. Agents 2008, 31, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate specificities of MexAB-OprM, MexCD-OprJ, and MexXY-oprM efflux pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Martínez, J.M.; Poirel, L.; Nordmann, P. Extended-spectrum cephalosporinases in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2009, 53, 1766–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalhoub, H.; Sáenz, Y.; Rodriguez-Villalobos, H.; Denis, O.; Kahl, B.C.; Tulkens, P.M.; Van Bambeke, F. High-level resistance to meropenem in clinical isolates of Pseudomonas aeruginosa in the absence of carbapenemases: Role of active efflux and porin alterations. Int. J. Antimicrob. Agents 2016, 48, 740–743. [Google Scholar] [CrossRef] [PubMed]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile beta-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matagne, A.; Dubus, A.; Galleni, M.; Frère, J.M. The beta-lactamase cycle: A tale of selective pressure and bacterial ingenuity. Nat. Prod. Rep. 1999, 16, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Frase, H.; Shi, Q.; Testero, S.A.; Mobashery, S.; Vakulenko, S.B. Mechanistic basis for the emergence of catalytic competence against carbapenem antibiotics by the GES family of beta-lactamases. J. Biol. Chem. 2009, 284, 29509–29513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vourli, S.; Giakkoupi, P.; Miriagou, V.; Tzelepi, E.; Vatopoulos, A.C.; Tzouvelekis, L.S. Novel GES/IBC extended-spectrum beta-lactamase variants with carbapenemase activity in clinical enterobacteria. FEMS Microbiol. Lett. 2004, 234, 209–213. [Google Scholar] [CrossRef]
- Bonnin, R.A.; Nordmann, P.; Potron, A.; Lecuyer, H.; Zahar, J.R.; Poirel, L. Carbapenem-Hydrolyzing GES-Type Extended-Spectrum β-Lactamase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Bogaerts, P.; Naas, T.; El Garch, F.; Cuzon, G.; Deplano, A.; Delaire, T.; Huang, T.D.; Lissoir, B.; Nordmann, P.; Glupczynski, Y. GES Extended-Spectrum β-Lactamases in Acinetobacter baumannii Isolates in Belgium. Antimicrob. Agents Chemother. 2010, 54, 4872–4878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, I.K.; Lee, Y.N.; Jeong, S.H.; Hong, S.G.; Lee, J.H.; Lee, S.H.; Kim, H.J.; Youn, H. Genetic and biochemical characterization of GES-5, an extended-spectrum class A beta-lactamase from Klebsiella pneumoniae. Diagn. Microbiol. Infect. Dis. 2007, 58, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Wachino, J.I.; Doi, Y.; Yamane, K.; Shibata, N.; Yagi, T.; Kubota, T.; Arakawa, Y. Molecular characterization of a cephamycin-hydrolyzing and inhibitor-resistant class A beta-lactamase, GES-4, possessing a single G170S substitution in the omega-loop. Antimicrob. Agents Chemother. 2004, 48, 2905–2910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hishinuma, T.; Tada, T.; Kuwahara-Arai, K.; Yamamoto, N.; Shimojima, M.; Kirikae, T. Spread of GES-5 carbapenemase-producing Pseudomonas aeruginosa clinical isolates in Japan due to clonal expansion of ST235. PLoS ONE 2018, 13, e0207134. [Google Scholar] [CrossRef]
- Malkoçoǧlu, G.; Aktaş, E.; Bayraktar, B.; Otlu, B.; Bulut, M.E. VIM-1, VIM-2, and GES-5 Carbapenemases among Pseudomonas aeruginosa Isolates at a Tertiary Hospital in Istanbul, Turkey. Microb. Drug Resist. 2017, 23, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Kostyanev, T.; Nguyen, M.N.; Markovska, R.; Stankova, P.; Xavier, B.B.; Lammens, C.; Marteva-Proevska, Y.; Velinov, T.; Cantón, R.; Goossens, H.; et al. Emergence of ST654 Pseudomonas aeruginosa co-harbouring blaNDM-1 and blaGES-5 in novel class I integron In1884 from Bulgaria. J. Glob. Antimicrob. Resist. 2020, 22, 672–673. [Google Scholar] [CrossRef]
- Villegas, M.V.; Lolans, K.; Correa, A.; Kattan, J.N.; Lopez, J.A.; Quinn, J.P. First identification of Pseudomonas aeruginosa isolates producing a KPC-type carbapenem-hydrolyzing beta-lactamase. Antimicrob. Agents Chemother. 2007, 51, 1553–1555. [Google Scholar] [CrossRef] [Green Version]
- De Araújo Jácome, P.R.L.; Rodrigues Alves, Ĺ.; Borges Cabral, A.; Lopes, A.C.S.; Vieira Maciel, M.A. First report of KPC-producing Pseudomonas aeruginosa in Brazil. Antimicrob. Agents Chemother. 2012, 56, 4990. [Google Scholar] [CrossRef] [Green Version]
- Ge, C.; Wei, Z.; Jiang, Y.; Shen, P.; Yu, Y.; Li, L. Identification of KPC-2-producing Pseudomonas aeruginosa isolates in China. J. Antimicrob. Chemother. 2011, 66, 1184–1186. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Nordmann, P.; Lagrutta, E.; Cleary, T.; Munoz-Price, L.S. Emergence of KPC-producing Pseudomonas aeruginosa in the United States. Antimicrob. Agents Chemother. 2010, 54, 3072. [Google Scholar] [CrossRef] [Green Version]
- Hagemann, J.B.; Pfennigwerth, N.; Gatermann, S.G.; von Baum, H.; Essig, A. KPC-2 carbapenemase-producing Pseudomonas aeruginosa reaching Germany. J. Antimicrob. Chemother. 2018, 73, 1812–1814. [Google Scholar] [CrossRef]
- Palzkill, T. Metallo-β-lactamase structure and function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104. [Google Scholar] [CrossRef]
- Watanabe, M.; Iyobe, S.; Inoue, M.; Mitsuhashi, S. Transferable imipenem resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1991, 35, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Naas, T.; Nicolas, D.; Collet, L.; Bellais, S.; Cavallo, J.D.; Nordmann, P. Characterization of VIM-2, a Carbapenem-Hydrolyzing Metallo-β-Lactamase and Its Plasmid- and Integron-Borne Gene from a Pseudomonas aeruginosa Clinical Isolate in France. Antimicrob. Agents Chemother. 2000, 44, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Liakopoulos, A.; Mavroidi, A.; Katsifas, E.A.; Theodosiou, A.; Karagouni, A.D.; Miriagou, V.; Petinaki, E. Carbapenemase-producing Pseudomonas aeruginosa from central Greece: Molecular epidemiology and genetic analysis of class I integrons. BMC Infect. Dis. 2013, 13, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahar, G.; Mazzariol, A.; Koncan, R.; Mert, A.; Fontana, R.; Rossolini, G.M.; Cornaglia, G. Detection of VIM-5 metallo-beta-lactamase in a Pseudomonas aeruginosa clinical isolate from Turkey. J. Antimicrob. Chemother. 2004, 54, 282–283. [Google Scholar] [CrossRef]
- Schneider, I.; Keuleyan, E.; Rasshofer, R.; Markovska, R.; Queenan, A.M.; Bauernfeind, A. VIM-15 and VIM-16, two new VIM-2-like metallo-beta-lactamases in Pseudomonas aeruginosa isolates from Bulgaria and Germany. Antimicrob. Agents Chemother. 2008, 52, 2977–2979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bośnjak, Z.; Bedenić, B.; Mazzariol, A.; Jarža-Davila, N.; Šuto, S.; Kalenić, S. VIM-2 beta-lactamase in Pseudomonas aeruginosa isolates from Zagreb, Croatia. Scand. J. Infect. Dis. 2010, 42, 193–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dortet, L.; Flonta, M.; Boudehen, Y.M.; Creton, E.; Bernabeu, S.; Vogel, A.; Naas, T. Dissemination of carbapenemase-producing Enterobacteriaceae and Pseudomonas aeruginosa in Romania. Antimicrob. Agents Chemother. 2015, 59, 7100–7103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazmierczak, K.M.; Rabine, S.; Hackel, M.; McLaughlin, R.E.; Biedenbach, D.J.; Bouchillon, S.K.; Sahm, D.F.; Bradford, P.A. Multiyear, Multinational Survey of the Incidence and Global Distribution of Metallo-β-Lactamase-Producing Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2015, 60, 1067–1078. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [Green Version]
- Struelens, M.J.; Monnet, D.L.; Magiorakos, A.P.; O’Connor, F.S.; Giesecke, J.; Grisold, A.; Zarfel, G.; Jans, B.; Velinov, T.; Kantardjiev, T.; et al. New Delhi metallo-beta-lactamase 1-producing Enterobacteriaceae: Emergence and response in Europe. Euro Surveill. 2010, 15, 19716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovcic, B.; Lepsanovic, Z.; Suljagic, V.; Rackov, G.; Begovic, J.; Topisirovic, L.; Kojic, M. Emergence of NDM-1 Metallo-β-Lactamase in Pseudomonas aeruginosa Clinical Isolates from Serbia. Antimicrob. Agents Chemother. 2011, 55, 3929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cayci, Y.T.; Biyik, İ.; Birinci, A. VIM, NDM, IMP, GES, SPM, GIM, SIM Metallobetalactamases in Carbapenem-Resistant Pseudomonas aeruginosa Isolates from a Turkish University Hospital. J. Arch. Mil. Med. 2022, 10, 118712. [Google Scholar] [CrossRef]
- Golemi, D.; Maveyraud, L.; Vakulenko, S.; Samama, J.P.; Mobashery, S. Critical involvement of a carbamylated lysine in catalytic function of class D beta-lactamases. Proc. Natl. Acad. Sci. USA 2001, 98, 14280–14285. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606. [Google Scholar] [CrossRef] [Green Version]
- del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A. Pseudomonas aeruginosa epidemic high-risk clones and their association with horizontally-acquired β-lactamases: 2020 update. Int. J. Antimicrob. Agents 2020, 56, 106196. [Google Scholar] [CrossRef]
- Tafaj, S.; Gona, F.; Rodrigues, C.F.; Kapisyzi, P.; Caushi, F.; Rossen, J.W.; Cirillo, D.M. Whole-Genome Sequences of Two NDM-1-Producing Pseudomonas aeruginosa Strains Isolated in a Clinical Setting in Albania in 2018. Microbiol. Resour. Announc. 2020, 9, e01291-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strateva, T.; Setchanova, L.; Peykov, S. Characterization of a Bulgarian VIM-2 metallo-β-lactamase-producing Pseudomonas aeruginosa clinical isolate belonging to the high-risk sequence type 111. Infect. Dis. 2021, 53, 883–887. [Google Scholar] [CrossRef]
- Papagiannitsis, C.C.; Verra, A.; Galani, V.; Xitsas, S.; Bitar, I.; Hrabak, J.; Petinaki, E. Unravelling the Features of Success of VIM-Producing ST111 and ST235 Pseudomonas aeruginosa in a Greek Hospital. Microorganisms 2020, 8, 1884. [Google Scholar] [CrossRef]
- Gheorghe-Barbu, I.; Barbu, I.C.; Popa, L.I.; Pîrcălăbioru, G.G.; Popa, M.; Măruțescu, L.; Niță-Lazar, M.; Banciu, A.; Stoica, C.; Gheorghe, Ș.; et al. Temporo-spatial variations in resistance determinants and clonality of Acinetobacter baumannii and Pseudomonas aeruginosa strains from Romanian hospitals and wastewaters. Antimicrob. Antimicrob. Resist. Infect. Control. 2022, 11, 115. [Google Scholar] [CrossRef] [PubMed]
- Kabic, J.; Fortunato, G.; Vaz-Moreira, I.; Kekic, D.; Jovicevic, M.; Pesovic, J.; Ranin, L.; Opavski, N.; Manaia, C.M.; Gajic, I. Dissemination of Metallo-β-Lactamase-Producing Pseudomonas aeruginosa in Serbian Hospital Settings: Expansion of ST235 and ST654 Clones. Int. J. Mol. Sci. 2023, 24, 1519. [Google Scholar] [CrossRef] [PubMed]
- Çekin, Z.K.; Dabos, L.; Malkoçoğlu, G.; Fortineau, N.; Bayraktar, B.; Iorga, B.I.; Naas, T.; Aktaş, E. Carbapenemase -producing Pseudomonas aeruginosa isolates from Turkey: First report of P. aeruginosa high-risk clones with VIM-5- and IMP-7-type carbapenemases in a tertiary hospital. Diagn. Microbiol. Infect. Dis. 2021, 99, 115174. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Molina-Mora, J.A.; Campos-Sánchez, R.; Rodríguez, C.; Shi, L.; García, F. High quality 3C de novo assembly and annotation of a multidrug resistant ST-111 Pseudomonas aeruginosa genome: Benchmark of hybrid and non-hybrid assemblers. Sci. Reports 2020, 10, 1392. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Lim, Y.S.; Yong, D.; Yum, J.H.; Chong, Y. Evaluation of the Hodge test and the imipenem-EDTA double-disk synergy test for differentiating metallo-β-lactamase-producing isolates of Pseudomonas spp. and Acinetobacter spp. J. Clin. Microbiol. 2003, 41, 4623–4629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, P.; Poirel, L.; Dortet, L. Rapid detection of carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2012, 18, 1503–1507. [Google Scholar] [CrossRef] [Green Version]
- Van Der Zwaluw, K.; De Haan, A.; Pluister, G.N.; Bootsma, H.J.; De Neeling, A.J.; Schouls, L.M. The carbapenem inactivation method (CIM), a simple and low-cost alternative for the Carba NP test to assess phenotypic carbapenemase activity in gram-negative rods. PLoS ONE 2015, 10, e0123690. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Jacobo, V.M.; Hernández-Ramírez, K.C.; Romo-Rodríguez, P.; Pérez-Gallardo, R.V.; Campos-García, J.; Félix Gutiérrez-Corona, J.; García-Merinos, J.P.; Meza-Carmen, V.; Silva-Sánchez, J.; Ramírez-Díaz, M.I. CrpP Is a Novel Ciprofloxacin-Modifying Enzyme Encoded by the Pseudomonas aeruginosa pUM505 Plasmid. Antimicrob. Agents Chemother. 2018, 62, e02629-17. [Google Scholar] [CrossRef] [Green Version]
- Khaledi, A.; Weimann, A.; Schniederjans, M.; Asgari, E.; Kuo, T.; Oliver, A.; Cabot, G.; Kola, A.; Gastmeier, P.; Hogardt, M.; et al. Predicting antimicrobial resistance in Pseudomonas aeruginosa with machine learning-enabled molecular diagnostics. EMBO Mol. Med. 2020, 12, e10264. [Google Scholar] [CrossRef]
- Treepong, P.; Kos, V.N.; Guyeux, C.; Blanc, D.S.; Bertrand, X.; Valot, B.; Hocquet, D. Global emergence of the widespread Pseudomonas aeruginosa ST235 clone. Clin. Microbiol. Infect. 2018, 24, 258–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaillard, M.; van Belkum, A.; Cady, K.C.; Creely, D.; Shortridge, D.; Blanc, B.; Barbu, E.M.; Dunne, W.M.; Zambardi, G.; Enright, M.; et al. Correlation between phenotypic antibiotic susceptibility and the resistome in Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2017, 50, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Kos, V.N.; Déraspe, M.; McLaughlin, R.E.; Whiteaker, J.D.; Roy, P.H.; Alm, R.A.; Corbeil, J.; Gardner, H. The resistome of Pseudomonas aeruginosa in relationship to phenotypic susceptibility. Antimicrob. Agents Chemother. 2015, 59, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakkala, H.; Samantarrai, D.; Gribskov, M.; Siddavattam, D. Comparative genome analysis reveals niche-specific genome expansion in Acinetobacter baumannii strains. PLoS ONE 2019, 14, e0218204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.-F.; Lan, C.-Y. Antimicrobial resistance in Acinetobacter baumannii: From bench to bedside. World J. Clin. Cases WJCC 2014, 2, 787. [Google Scholar] [CrossRef]
- Doi, Y.; Murray, G.L.; Peleg, A.Y. Acinetobacter baumannii: Evolution of antimicrobial resistance-treatment options. Semin. Respir. Crit. Care Med. 2015, 36, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Nordmann, P. Carbapenem resistance in Acinetobacter baumannii: Mechanisms and epidemiology. Clin. Microbiol. Infect. 2006, 12, 826–836. [Google Scholar] [CrossRef] [Green Version]
- Traglia, G.M.; Chua, K.; Centron, D.; Tolmasky, M.E.; Ramírez, M.S. Whole-Genome Sequence Analysis of the Naturally Competent Acinetobacter baumannii Clinical Isolate A118. Genome Biol. Evol. 2014, 6, 2235–2239. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Don, M.; Merkier, A.K.; Bistué, A.J.S.; Zorreguieta, A.; Centrón, D.; Tolmasky, M.E. Naturally competent Acinetobacter baumannii clinical isolate as a convenient model for genetic studies. J. Clin. Microbiol. 2010, 48, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Traglia, G.M.; Quinn, B.; Schramm, S.T.J.; Soler-Bistue, A.; Ramirez, M.S. Serum Albumin and Ca2+ Are Natural Competence Inducers in the Human Pathogen Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 4920–4929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moubareck, C.; Brémont, S.; Conroy, M.C.; Courvalin, P.; Lambert, T. GES-11, a novel integron-associated GES variant in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 3579–3581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robledo, I.E.; Aquino, E.E.; Santé, M.I.; Santana, J.L.; Otero, D.M.; León, C.F.; Vázquez, G.J. Detection of KPC in Acinetobacter spp. in Puerto Rico. Antimicrob. Agents Chemother. 2010, 54, 1354. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, P.C.S.; Monteiro, A.S.; Marques, S.G.; Monteiro, S.G.; Monteiro-Neto, V.; Coqueiro, M.M.M.; Marques, A.C.G.; de Jesus Gomes Turri, R.; Santos, S.G.; Bomfim, M.R.Q. Phenotypic and molecular detection of the blaKPC gene in clinical isolates from inpatients at hospitals in São Luis, MA, Brazil. BMC Infect. Dis. 2016, 16, 737. [Google Scholar] [CrossRef] [Green Version]
- Lima, W.G.; Silva Alves, G.C.; Sanches, C.; Antunes Fernandes, S.O.; de Paiva, M.C. Carbapenem-resistant Acinetobacter baumannii in patients with burn injury: A systematic review and meta-analysis. Burns 2019, 45, 1495–1508. [Google Scholar] [CrossRef] [PubMed]
- Tognim, M.C.B.; Gales, A.C.; Penteado, A.P.; Silbert, S.; Sader, H.S. Dissemination of IMP-1 metallo- beta -lactamase-producing Acinetobacter species in a Brazilian teaching hospital. Infect. Control. Hosp. Epidemiol. 2006, 27, 742–747. [Google Scholar] [CrossRef]
- Riccio, M.L.; Franceschini, N.; Boschi, L.; Caravelli, B.; Cornaglia, G.; Fontana, R.; Amicosante, G.; Rossolini, G.M. Characterization of the metallo-beta-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of bla(IMP) allelic variants carried by gene cassettes of different phylogeny. Antimicrob. Agents Chemother. 2000, 44, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.W.; Afzal-Shah, M.; Houang, E.T.S.; Palepou, M.F.I.; Lyon, D.J.; Woodford, N.; Livermore, D.M. IMP-4, a novel metallo-beta-lactamase from nosocomial Acinetobacter spp. collected in Hong Kong between 1994 and 1998. Antimicrob. Agents Chemother. 2001, 45, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.J.; Correia, M.; Vital, C.; Ribeiro, G.; Sousa, J.C.; Leitão, R.; Peixe, L.; Duarte, A. Molecular characterization of bla(IMP-5), a new integron-borne metallo-beta-lactamase gene from an Acinetobacter baumannii nosocomial isolate in Portugal. FEMS Microbiol. Lett. 2002, 215, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Gales, A.C.; Tognim, M.C.B.; Reis, A.O.; Jones, R.N.; Sader, H.S. Emergence of an IMP-like metallo-enzyme in an Acinetobacter baumannii clinical strain from a Brazilian teaching hospital. Diagn. Microbiol. Infect. Dis. 2003, 45, 77–79. [Google Scholar] [CrossRef]
- Lee, M.F.; Peng, C.F.; Hsu, H.J.; Chen, Y.H. Molecular characterisation of the metallo-beta-lactamase genes in imipenem-resistant Gram-negative bacteria from a university hospital in southern Taiwan. Int. J. Antimicrob. Agents 2008, 32, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Nagao, M.; Matsumura, Y.; Matsushima, A.; Ito, Y.; Takakura, S.; Ichiyama, S. Interspecies dissemination of a novel class 1 integron carrying blaIMP-19 among Acinetobacter species in Japan. J. Antimicrob. Chemother. 2011, 66, 2480–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakibaie, M.R.; Azizi, O.; Shahcheraghi, F. Insight into stereochemistry of a new IMP allelic variant (IMP-55) metallo-β-lactamase identified in a clinical strain of Acinetobacter baumannii. Infect. Genet. Evol. 2017, 51, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, Z.; Jiang, Y.; Yu, Y. Emergence of NDM-1-producing Acinetobacter baumannii in China. J. Antimicrob. Chemother. 2011, 66, 1255–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girija, S.A.S.; Priyadharsini, J.V.; Arumugam, P. Prevalence of VIM- and GIM-producing Acinetobacter baumannii from patients with severe urinary tract infection. Acta Microbiol. Immunol. Hung. 2018, 65, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Espinal, P.; Fugazza, G.; López, Y.; Kasma, M.; Lerman, Y.; Malhotra-Kumar, S.; Goossens, H.; Carmeli, Y.; Vila, J. Dissemination of an NDM-2-producing Acinetobacter baumannii clone in an Israeli rehabilitation center. Antimicrob. Agents Chemother. 2011, 55, 5396–5398. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M. Identification of a Novel NDM Variant, blaNDM-3, from a Multidrug-Resistant Acinetobacter baumannii. Infect. Control. Hosp. Epidemiol. 2016, 37, 747–748. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Yum, J.H.; Yong, D.; Lee, H.M.; Kim, H.D.; Docquier, J.D.; Rossolini, G.M.; Chong, Y. Novel acquired metallo-beta-lactamase gene, bla(SIM-1), in a class 1 integron from Acinetobacter baumannii clinical isolates from Korea. Antimicrob. Agents Chemother. 2005, 49, 4485–4491. [Google Scholar] [CrossRef] [Green Version]
- Tsakris, A.; Ikonomidis, A.; Pournaras, S.; Tzouvelekis, L.S.; Sofianou, D.; Legakis, N.J.; Maniatis, A.N. VIM-1 metallo-beta-lactamase in Acinetobacter baumannii. Emerg. Infect. Dis. 2006, 12, 981–983. [Google Scholar] [CrossRef] [Green Version]
- Yum, J.H.; Yi, K.; Lee, H.; Yong, D.; Lee, K.; Kim, J.M.; Rossolini, G.M.; Chong, Y. Molecular characterization of metallo-beta-lactamase-producing Acinetobacter baumannii and Acinetobacter genomospecies 3 from Korea: Identification of two new integrons carrying the bla(VIM-2) gene cassettes. J. Antimicrob. Chemother. 2002, 49, 837–840. [Google Scholar] [CrossRef] [Green Version]
- Tsakris, A.; Ikonomidis, A.; Poulou, A.; Spanakis, N.; Vrizas, D.; Diomidous, M.; Pournaras, S.; Markou, F. Clusters of imipenem-resistant Acinetobacter baumannii clones producing different carbapenemases in an intensive care unit. Clin. Microbiol. Infect. 2008, 14, 588–594. [Google Scholar] [CrossRef] [Green Version]
- Walther-Rasmussen, J.; Høiby, N. OXA-type carbapenemases. J. Antimicrob. Chemother. 2006, 57, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarrilli, R.; Giannouli, M.; Tomasone, F.; Triassi, M.; Tsakris, A. Carbapenem resistance in Acinetobacter baumannii: The molecular epidemic features of an emerging problem in health care facilities. J. Infect. Dev. Ctries. 2009, 3, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Mugnier, P.D.; Poirel, L.; Naas, T.; Nordmann, P. Worldwide dissemination of the blaOXA-23 Carbapenemase gene of Acinetobacter baumannii. Emerg. Infect. Dis. 2010, 16, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, R.A.; Poirel, L.; Licker, M.; Nordmann, P. Genetic diversity of carbapenem-hydrolysing β-lactamases in Acinetobacter baumannii from Romanian hospitals. Clin. Microbiol. Infect. 2011, 17, 1524–1528. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Naas, T.; Nordmann, P. Diversity, epidemiology, and genetics of class D beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 24–38. [Google Scholar] [CrossRef] [Green Version]
- Goic-Barisic, I.; Towner, K.J.; Kovacic, A.; Sisko-Kraljevic, K.; Tonkic, M.; Novak, A.; Punda-Polic, V. Outbreak in Croatia caused by a new carbapenem-resistant clone of Acinetobacter baumannii producing OXA-72 carbapenemase. J. Hosp. Infect. 2011, 77, 368–369. [Google Scholar] [CrossRef]
- Héritier, C.; Poirel, L.; Fournier, P.E.; Claverie, J.M.; Raoult, D.; Nordmann, P. Characterization of the naturally occurring oxacillinase of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 4174–4179. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.L.; Lee, Y.T.; Kuo, S.C.; Hsueh, P.R.; Chang, F.Y.; Siu, L.K.; Ko, W.C.; Fung, C.P. Emergence and Distribution of Plasmids Bearing the blaOXA-51-like gene with an upstream ISAba1 in carbapenem-resistant Acinetobacter baumannii isolates in Taiwan. Antimicrob. Agents Chemother. 2010, 54, 4575–4581. [Google Scholar] [CrossRef] [Green Version]
- Pournaras, S.; Markogiannakis, A.; Ikonomidis, A.; Kondyli, L.; Bethimouti, K.; Maniatis, A.N.; Legakis, N.J.; Tsakris, A. Outbreak of multiple clones of imipenem-resistant Acinetobacter baumannii isolates expressing OXA-58 carbapenemase in an intensive care unit. J. Antimicrob. Chemother. 2006, 57, 557–561. [Google Scholar] [CrossRef]
- Vahaboglu, H.; Budak, F.; Kasap, M.; Gacar, G.; Torol, S.; Karadenizli, A.; Kolayli, F.; Eroglu, C. High prevalence of OXA-51-type class D β-lactamases among ceftazidime-resistant clinical isolates of Acinetobacter spp.: Co-existence with OXA-58 in multiple centres. J. Antimicrob. Chemother. 2006, 58, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.L.; Wu, R.C.C.; Shaio, M.F.; Fung, C.P.; Cho, W.L. Acquisition of a plasmid-borne blaOXA-58 gene with an upstream IS1008 insertion conferring a high level of carbapenem resistance to Acinetobacter baumannii. Antimicrob. Agents Chemother. 2008, 52, 2573–2580. [Google Scholar] [CrossRef] [Green Version]
- Ravasi, P.; Limansky, A.S.; Rodriguez, R.E.; Viale, A.M.; Mussi, M.A. ISAba825, a functional insertion sequence modulating genomic plasticity and bla(OXA-58) expression in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 917–920. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Pérez-Llarena, F.J.; Zander, E.; Fernández, A.; Bou, G.; Seifert, H. OXA-235, a novel class D β-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2013, 57, 2121–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.K.; Lee, Y.; Lee, H.; Woo, G.J.; Song, W.; Kim, M.N.; Lee, W.G.; Jeong, S.H.; Lee, K.; Chong, Y. Prevalence and diversity of carbapenemases among imipenem-nonsusceptible Acinetobacter isolates in Korea: Emergence of a novel OXA-182. Diagn. Microbiol. Infect. Dis. 2010, 68, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.G.; Poirel, L.; Lehmann, M.; Nordmann, P.; Seifert, H. OXA-143, a novel carbapenem-hydrolyzing class D β-lactamase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 5035–5038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, Q.; Zou, M.; Li, J.; Wang, H.; Hu, Y.; Liu, W. AdeABC efflux pump and resistance of Acinetobacter baumannii against carbapenem. J. Cent. South Univ. Med. Sci. 2017, 42, 426–433. [Google Scholar] [CrossRef]
- Su, X.Z.; Chen, J.; Mizushima, T.; Kuroda, T.; Tsuchiya, T. AbeM, an H+-coupled Acinetobacter baumannii multidrug efflux pump belonging to the MATE family of transporters. Antimicrob. Agents Chemother. 2005, 49, 4362–4364. [Google Scholar] [CrossRef] [Green Version]
- Benmahmod, A.B.; Said, H.S.; Ibrahim, R.H. Prevalence and Mechanisms of Carbapenem Resistance Among Acinetobacter baumannii Clinical Isolates in Egypt. Microb. Drug Resist. 2019, 25, 480–488. [Google Scholar] [CrossRef]
- Bou, G.; Cervero, G.; Dominguez, M.A.; Quereda, C.; Martinez-Beltran, J. Characterization of a nosocomial outbreak caused by a multiresistant Acinetobacter baumannii strain with a carbapenem-hydrolyzing enzyme: High-level carbapenem resistance in A. baumannii is not due solely to the presence of beta-lactamases. J. Clin. Microbiol. 2000, 38, 3299–3305. [Google Scholar] [CrossRef] [Green Version]
- Tomás, M.D.M.; Beceiro, A.; Pérez, A.; Velasco, D.; Moure, R.; Villanueva, R.; Martínez-Beltrán, J.; Bou, G. Cloning and functional analysis of the gene encoding the 33- to 36-kilodalton outer membrane protein associated with carbapenem resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 5172–5175. [Google Scholar] [CrossRef] [Green Version]
- Quale, J.; Bratu, S.; Landman, D.; Heddurshetti, R. Molecular epidemiology and mechanisms of carbapenem resistance in Acinetobacter baumannii endemic in New York City. Clin. Infect. Dis. 2003, 37, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Gehrlein, M.; Leying, H.; Cullmann, W.; Wendt, S.; Opferkuch, W. Imipenem resistance in Acinetobacter baumanii is due to altered penicillin-binding proteins. Chemotherapy 1991, 37, 405–412. [Google Scholar] [CrossRef]
- Hussein, N.H.; AL-Kadmy, I.M.S.; Taha, B.M.; Hussein, J.D. Mobilized colistin resistance (mcr) genes from 1 to 10: A comprehensive review. Mol. Biol. Rep. 2021, 48, 2897–2907. [Google Scholar] [CrossRef]
- Ling, Z.; Yin, W.; Shen, Z.; Wang, Y.; Shen, J.; Walsh, T.R. Epidemiology of mobile colistin resistance genes mcr-1 to mcr-9. J. Antimicrob. Chemother. 2020, 75, 3087–3095. [Google Scholar] [CrossRef]
- Maciuca, I.E.; Cummins, M.L.; Cozma, A.P.; Rimbu, C.M.; Guguianu, E.; Panzaru, C.; Licker, M.; Szekely, E.; Flonta, M.; Djordjevic, S.P.; et al. Genetic Features of mcr-1 Mediated Colistin Resistance in CMY-2-Producing Escherichia coli from Romanian Poultry. Front. Microbiol. 2019, 10, 2267. [Google Scholar] [CrossRef] [Green Version]
- Mišić, D.; Kiskaroly, F.; Szostak, M.P.; Cabal, A.; Ruppitsch, W.; Bernreiter-Hofer, T.; Milovanovic, V.; Feßler, A.T.; Allerberger, F.; Spergser, J.; et al. The First Report of mcr-1-Carrying Escherichia coli Originating from Animals in Serbia. Antibiotics 2021, 10, 1063. [Google Scholar] [CrossRef]
- Kurekci, C.; Aydin, M.; Nalbantoglu, O.U.; Gundogdu, A. First report of Escherichia coli carrying the mobile colistin resistance gene mcr-1 in Turkey. J. Glob. Antimicrob. Resist. 2018, 15, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Protonotariou, E.; Meletis, G.; Malousi, A.; Kotzamanidis, C.; Tychala, A.; Mantzana, P.; Theodoridou, K.; Ioannidou, M.; Hatzipantelis, E.; Tsakris, A.; et al. First detection of mcr-1-producing Escherichia coli in Greece. J. Glob. Antimicrob. Resist. 2022, 31, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Jovcic, B.; Novovic, K.; Dekic, S.; Hrenovic, J. Colistin Resistance in Environmental Isolates of Acinetobacter baumannii. Microb. Drug Resist. 2021, 27, 328–336. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, V.; Conzemius, R.; Varda-Brkić, D.; Bogdan, M.; Grisold, A.; Gyssens, I.C.; Bedenić, B.; Barišić, I. Epidemiology of colistin-resistant, carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Croatia. Infect. Genet. Evol. 2020, 81, 104263. [Google Scholar] [CrossRef]
- Hamel, M.; Rolain, J.M.; Baron, S.A. The History of Colistin Resistance Mechanisms in Bacteria: Progress and Challenges. Microorganisms 2021, 9, 442. [Google Scholar] [CrossRef]
- Jieling, Z. Mechanism of Colistin Resistance to Acinetobacter baumannii and its Progress: A Review Article. Biomed. J. Sci. Tech. Res. 2020, 29, 22183–22188. [Google Scholar] [CrossRef]
- Cai, Y.; Chai, D.; Wang, R.; Liang, B.; Bai, N. Colistin resistance of Acinetobacter baumannii: Clinical reports, mechanisms and antimicrobial strategies. J. Antimicrob. Chemother. 2012, 67, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Gerson, S.; Betts, J.W.; Lucaßen, K.; Nodari, C.S.; Wille, J.; Josten, M.; Göttig, S.; Nowak, J.; Stefanik, D.; Roca, I.; et al. Investigation of Novel pmrB and eptA Mutations in Isogenic Acinetobacter baumannii Isolates Associated with Colistin Resistance and Increased Virulence In Vivo. Antimicrob. Agents Chemother. 2019, 63, e01586-18. [Google Scholar] [CrossRef] [Green Version]
- Abdelbary, M.M.H.; Prod’hom, G.; Greub, G.; Senn, L.; Blanc, D.S. Draft Genome Sequences of Two Carbapenemase-Producing Acinetobacter baumannii Clinical Strains Isolated from Albanian and Togolese Patients. Genome Announc. 2017, 5, e00115-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamidian, M.; Wick, R.R.; Judd, L.M.; Holt, K.E.; Hall, R.M. Complete Genome Sequence of A388, an Antibiotic-Resistant Acinetobacter baumannii Global Clone 1 Isolate from Greece. Microbiol. Resour. Announc. 2019, 8, e00971-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, M.; D’Andrea, M.M.; Pelegrin, A.C.; Perrot, N.; Mirande, C.; Blanc, B.; Legakis, N.; Goossens, H.; Rossolini, G.M.; van Belkum, A. Abundance of Colistin-Resistant, OXA-23- and ArmA-Producing Acinetobacter baumannii Belonging to International Clone 2 in Greece. Front. Microbiol. 2020, 11, 668. [Google Scholar] [CrossRef] [Green Version]
- Gheorghe, I.; Barbu, I.C.; Surleac, M.; Sârbu, I.; Popa, L.I.; Paraschiv, S.; Feng, Y.; Lazăr, V.; Chifiriuc, M.C.; Oţelea, D.; et al. Subtypes, resistance and virulence platforms in extended-drug resistant Acinetobacter baumannii Romanian isolates. Sci. Rep. 2021, 11, 13288. [Google Scholar] [CrossRef]
- Dortet, L.; Bonnin, R.A.; Girlich, D.; Imanci, D.; Bernabeu, S.; Fortineau, N.; Naas, T. Whole-Genome Sequence of a European Clone II and OXA-72-Producing Acinetobacter baumannii Strain from Serbia. Genome Announc. 2015, 3, 1390–1405. [Google Scholar] [CrossRef] [Green Version]
- Kabic, J.; Novovic, K.; Kekic, D.; Trudic, A.; Opavski, N.; Dimkic, I.; Jovcic, B.; Gajic, I. Comparative genomics and molecular epidemiology of colistin-resistant Acinetobacter baumannii. Comput. Struct. Biotechnol. J. 2023, 21, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Gülbüz, M.; Saral Sariyer, A. Combined in silico approach and whole genome sequencing: Acinetobacter baumannii ST218 isolate harboring ADC-73 β-lactamase which has a similar C-loop with ADC-56 and ADC-68 β-lactamase. J. Mol. Graph. Model. 2022, 114, 108195. [Google Scholar] [CrossRef] [PubMed]
- Vidaillac, C.; Benichou, L.; Duval, R.E. In vitro synergy of colistin combinations against colistin-resistant Acinetobacter baumannii, Pseudomonas aeruginosa, and Klebsiella pneumoniae isolates. Antimicrob. Agents Chemother. 2012, 56, 4856–4861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavroidi, A.; Likousi, S.; Palla, E.; Katsiari, M.; Roussou, Z.; Maguina, A.; Platsouka, E.D. Molecular identification of tigecycline- and colistinresistant carbapenemase-producing Acinetobacter baumannii from a Greek hospital from 2011 to 2013. J. Med. Microbiol. 2015, 64, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, O.; Sarrou, S.; Papagiannitsis, C.C.; Georgiadou, S.; Mantzarlis, K.; Zakynthinos, E.; Dalekos, G.N.; Petinaki, E. Rapid dissemination of colistin and carbapenem resistant Acinetobacter baumannii in Central Greece: Mechanisms of resistance, molecular identification and epidemiological data. BMC Infect. Dis. 2015, 15, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostyanev, T.; Xavier, B.B.; García-Castillo, M.; Lammens, C.; Bravo-Ferrer Acosta, J.; Rodríguez-Baño, J.; Cantón, R.; Glupczynski, Y.; Goossens, H. Phenotypic and molecular characterizations of carbapenem-resistant Acinetobacter baumannii isolates collected within the EURECA study. Int. J. Antimicrob. Agents 2021, 57, 106345. [Google Scholar] [CrossRef]
- Carrara, J.B.A.; Barroso, C.D.N.; Tuon, F.F.; Faoro, H.; Lorusso, A.B.; Carrara, J.A.; Deuttner, C.; Barroso, N.; Tuon, F.F.; Faoro, H. Role of Efflux Pumps on Antimicrobial Resistance in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2022, 23, 15779. [Google Scholar] [CrossRef]
- Poole, K. Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 2007, 39, 162–176. [Google Scholar] [CrossRef]
- Gil-Gil, T.; Martínez, J.L.; Blanco, P. Mechanisms of antimicrobial resistance in Stenotrophomonas maltophilia: A review of current knowledge. Expert Rev. Anti. Infect. Ther. 2020, 18, 335–347. [Google Scholar] [CrossRef]
- García-León, G.; Hernández, A.; Hernando-Amado, S.; Alavi, P.; Berg, G.; Martínez, J.L. A Function of SmeDEF, the Major Quinolone Resistance Determinant of Stenotrophomonas maltophilia, Is the Colonization of Plant Roots. Appl. Environ. Microbiol. 2014, 80, 4559. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.; Martinez, J.L. Cloning and Characterization of SmeDEF, a Novel Multidrug Efflux Pump from Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2000, 44, 3079. [Google Scholar] [CrossRef] [Green Version]
- Ibn Saied, W.; Merceron, S.; Schwebel, C.; Le Monnier, A.; Oziel, J.; Garrouste-Orgeas, M.; Marcotte, G.; Ruckly, S.; Souweine, B.; Darmon, M.; et al. Ventilator-associated pneumonia due to Stenotrophomonas maltophilia: Risk factors and outcome. J. Infect. 2020, 80, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Lin, J.C.; Chang, F.Y.; Yu, C.M.; Lin, W.S.; Yeh, K.M. Risk factors for hospital acquisition of trimethoprim-sulfamethoxazole resistant Stenotrophomonas maltophilia in adults: A matched case-control study. J. Microbiol. Immunol. Infect. 2017, 50, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Crowder, M.W.; Walsh, T.R.; Banovic, L.; Pettit, M.; Spencer, J. Overexpression, purification, and characterization of the cloned metallo-beta-lactamase L1 from Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 1998, 42, 921–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, T.R.; MacGowan, A.P.; Bennett, P.M. Sequence analysis and enzyme kinetics of the L2 serine beta-lactamase from Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 1997, 41, 1460–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.W.; Huang, Y.W.; Hu, R.M.; Chiang, K.H.; Yang, T.C. The role of AmpR in regulation of L1 and L2 beta-lactamases in Stenotrophomonas maltophilia. Res. Microbiol. 2009, 160, 152–158. [Google Scholar] [CrossRef]
- Yang, T.C.; Huang, Y.W.; Hu, R.M.; Huang, S.C.; Lin, Y.T. AmpDI is involved in expression of the chromosomal L1 and L2 beta-lactamases of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2009, 53, 2902–2907. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.W.; Lin, C.W.; Hu, R.M.; Lin, Y.T.; Chung, T.C.; Yang, T.C. AmpN-AmpG operon is essential for expression of L1 and L2 beta-lactamases in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2010, 54, 2583–2589. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Zhang, L.; McKay, G.A.; Poole, K. Role of the acetyltransferase AAC(6′)-Iz modifying enzyme in aminoglycoside resistance in Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 2003, 51, 803–811. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, A.; Avison, M.B. Aph(3′)-IIc, an aminoglycoside resistance determinant from Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2007, 51, 359–360. [Google Scholar] [CrossRef] [Green Version]
- Tada, T.; Miyoshi-Akiyama, T.; Dahal, R.K.; Mishra, S.K.; Shimada, K.; Ohara, H.; Kirikae, T.; Pokhrelc, B.M. Identification of a novel 6′-N-aminoglycoside acetyltransferase, AAC(6′)-Iak, from a multidrug-resistant clinical isolate of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2014, 58, 6324–6327. [Google Scholar] [CrossRef] [Green Version]
- Al-Hamad, A.; Upton, M.; Burnie, J. Molecular cloning and characterization of SmrA, a novel ABC multidrug efflux pump from Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 2009, 64, 731–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.T.; Huang, Y.W.; Liou, R.S.; Chang, Y.C.; Yang, T.C. MacABCsm, an ABC-type tripartite efflux pump of Stenotrophomonas maltophilia involved in drug resistance, oxidative and envelope stress tolerances and biofilm formation. J. Antimicrob. Chemother. 2014, 69, 3221–3226. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.W.; Hu, R.M.; Chu, F.Y.; Lin, H.R.; Yang, T.C. Characterization of a major facilitator superfamily (MFS) tripartite efflux pump EmrCABsm from Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 2013, 68, 2498–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, R.M.; Liao, S.T.; Huang, C.C.; Huang, Y.W.; Yang, T.C. An Inducible Fusaric Acid Tripartite Efflux Pump Contributes to the Fusaric Acid Resistance in Stenotrophomonas maltophilia. PLoS ONE 2012, 7, e51053. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.B.; Martínez, J.L. Overexpression of the Efflux Pumps SmeVWX and SmeDEF Is a Major Cause of Resistance to Co-trimoxazole in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2018, 62, e00301-18. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.J.; Lu, H.F.; Lin, Y.T.; Zhang, M.S.; Li, L.H.; Yang, T.C. Substantial Contribution of SmeDEF, SmeVWX, SmQnr, and Heat Shock Response to Fluoroquinolone Resistance in Clinical Isolates of Stenotrophomonas maltophilia. Front. Microbiol. 2019, 10, 822. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, K.; Kikuchi, K.; Sasaki, T.; Takahashi, N.; Ohtsuka, M.; Ono, Y.; Hiramatsu, K. Smqnr, a New Chromosome-Carried Quinolone Resistance Gene in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2008, 52, 3823. [Google Scholar] [CrossRef] [Green Version]
- Gordon, N.C.; Wareham, D.W. Novel variants of the Smqnr family of quinolone resistance genes in clinical isolates of Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 2010, 65, 483–489. [Google Scholar] [CrossRef]
- Adegoke, A.A.; Stenström, T.A.; Okoh, A.I. Stenotrophomonas maltophilia as an emerging ubiquitous pathogen: Looking beyond contemporary antibiotic therapy. Front. Microbiol. 2017, 8, 2276. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.F.; Chang, X.; Ye, Y.; Wang, Z.X.; Shao, Y.B.; Shi, W.; Li, X.; Li, J. Bin Stenotrophomonas maltophilia resistance to trimethoprim/sulfamethoxazole mediated by acquisition of sul and dfrA genes in a plasmid-mediated class 1 integron. Int. J. Antimicrob. Agents 2011, 37, 230–234. [Google Scholar] [CrossRef]
- Barbolla, R.; Catalano, M.; Orman, B.E.; Famiglietti, A.; Vay, C.; Smayevsky, J.; Centrón, D.; Piñeiro, S.A. Class 1 Integrons Increase Trimethoprim-Sulfamethoxazole MICs against Epidemiologically Unrelated Stenotrophomonas maltophilia Isolates. Antimicrob. Agents Chemother. 2004, 48, 666–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toleman, M.A.; Bennett, P.M.; Bennett, D.M.C.; Jones, R.N.; Walsh, T.R. Global emergence of trimethoprim/sulfamethoxazole resistance in Stenotrophomonas maltophilia mediated by acquisition of sul genes. Emerg. Infect. Dis. 2007, 13, 559–565. [Google Scholar] [CrossRef]
- Strateva, T.; Trifonova, A.; Savov, E.; Mitov, I.; Peykov, S. Characterization of an extensively drug-resistant Stenotrophomonas maltophilia clinical isolate with strong biofilm formation ability from Bulgaria. Infect. Dis. 2020, 52, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Libisch, B.; Poirel, L.; Lepsanovic, Z.; Mirovic, V.; Balogh, B.; Pászti, J.; Hunyadi, Z.; Dobák, A.; Füzi, M.; Nordmann, P. Identification of PER-1 extended-spectrum beta-lactamase producing Pseudomonas aeruginosa clinical isolates of the international clonal complex CC11 from Hungary and Serbia. FEMS Immunol. Med. Microbiol. 2008, 54, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strateva, T.; Trifonova, A.; Sirakov, I.; Borisova, D.; Stancheva, M.; Keuleyan, E.; Setchanova, L.; Peykov, S. Analysis of biofilm formation in nosocomial Stenotrophomonas maltophilia isolates collected in Bulgaria: An 11-year study (2011–2022). Acta Microbiol. Immunol. Hung. 2023, 70, 11–21. [Google Scholar] [CrossRef]
- Ciofu, O.; Rojo-Molinero, E.; Macià, M.D.; Oliver, A. Antibiotic treatment of biofilm infections. APMIS 2017, 125, 304–319. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]




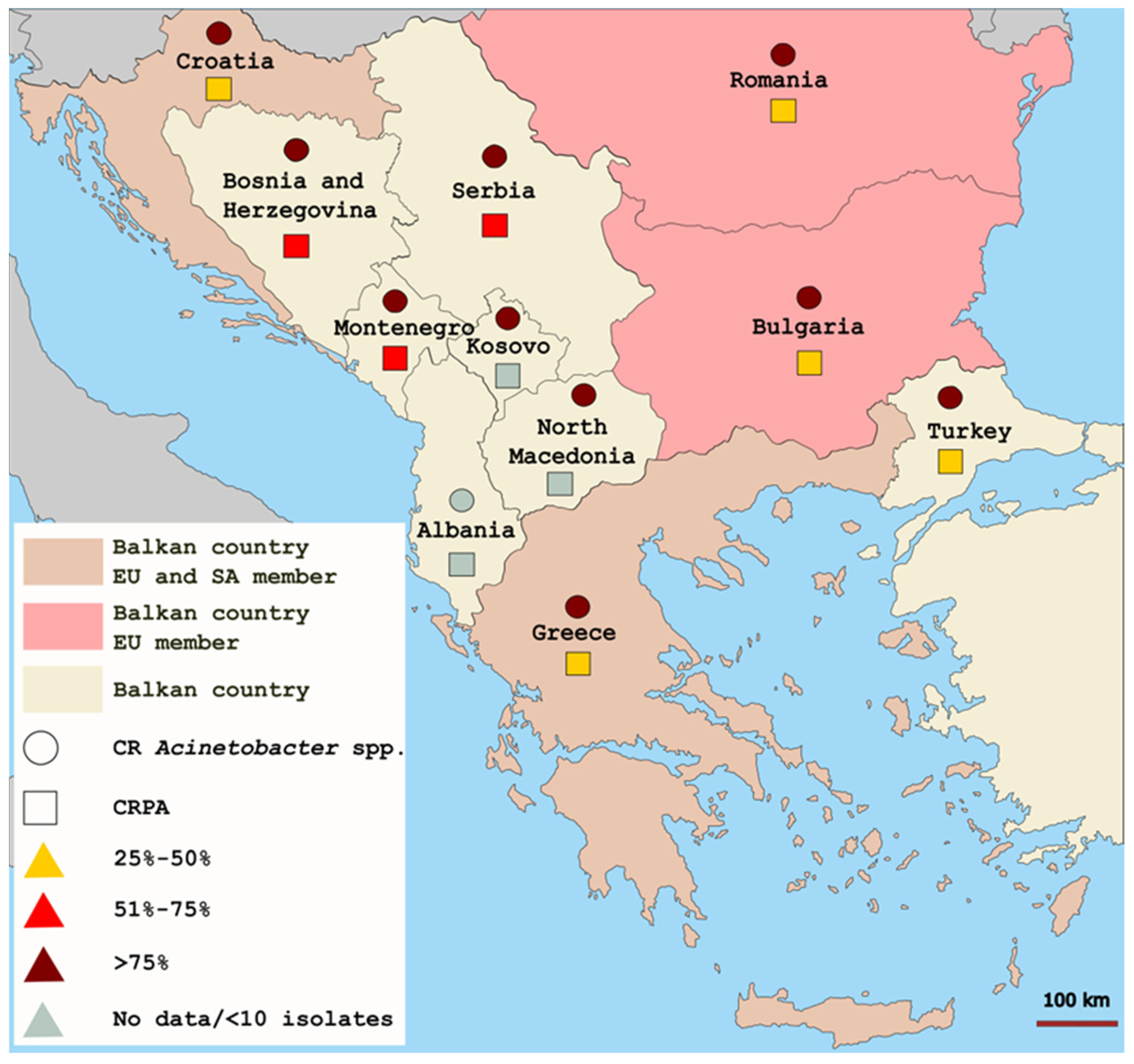{kind=link}
{kind=link}
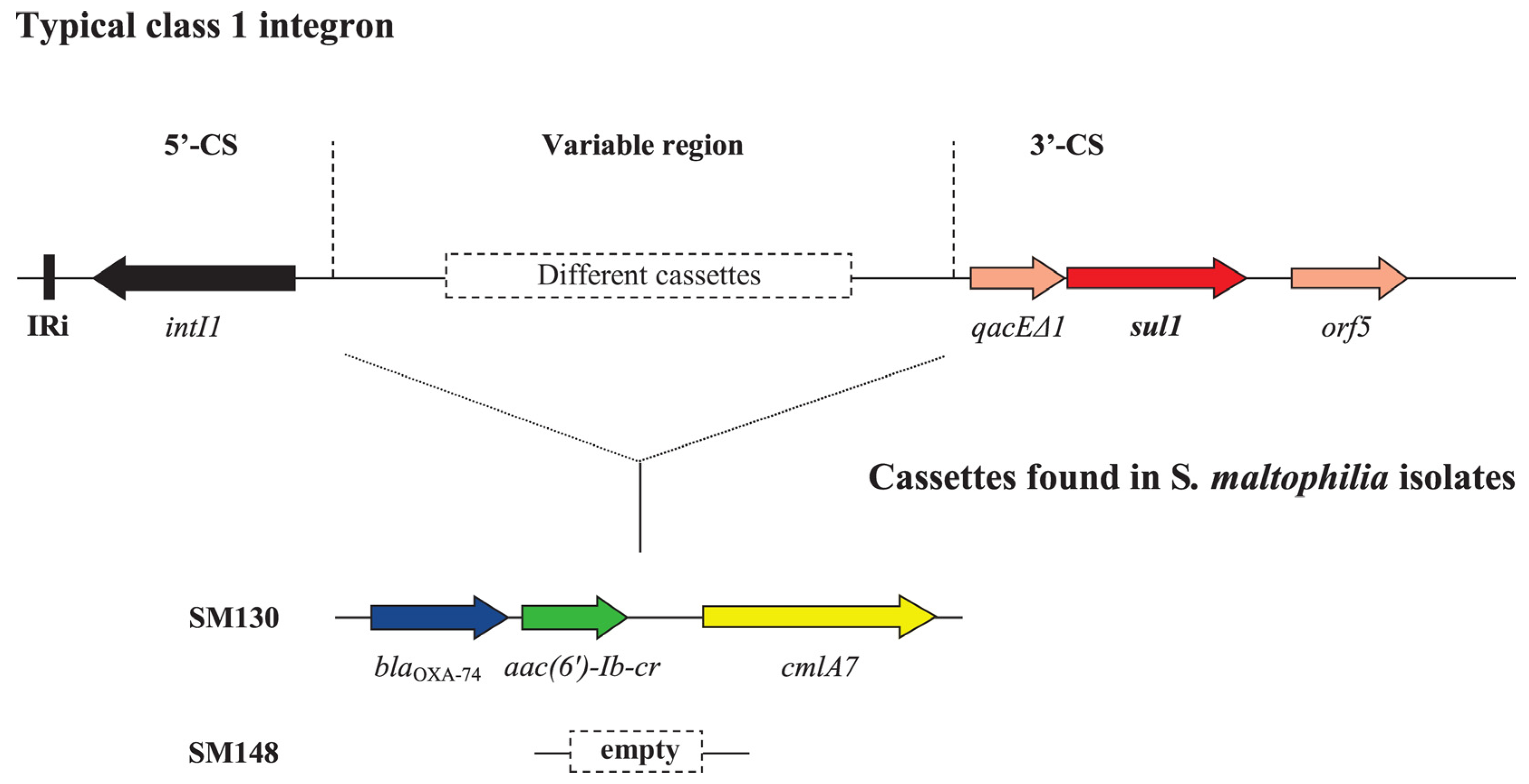{kind=link}
| Country | Isolates Analyzed | Year | β-Lactam Resistance | Aminoglycoside Resistance | Fluoroquinolone Resistance | Other AMR Determinants | Source |
|---|---|---|---|---|---|---|---|
| Albania | 2 isolates, CRPA, ST235 | 2018 | blaOXA-488, blaNDM-1, blaPDC-2 | aac(6′)-Il, ant(2″)-Ia, aph(3′)-IIb | cpxR, pmpM, gyrA mutations | bcr1, catB7, emrE, fosA, sul1 | [127] |
| Bulgaria | 5 isolates, XDR, ST654 | 2017–2018 | blaNDM-1, blaGES-1, blaGES-5 | strA, strB, aph(3′)-Via, aadB | sul1, sul3, tetA, tetR | [105] | |
| 1 isolate, XDR, ST111 | 2019 | blaVIM-2, blaPAO, blaOXA-395 | aac(6′)-29a, aph(30)-IIb, ant(3″)-Ia | crpP, gyrA and parC mutations | catB7, cmlB1, fosA, sul1 | [128] | |
| Greece | 15 isolates, CRPA, ST111 (x2), ST235 (x6), ST162 (x5), ST395 (x2) | 2018 | blaVIM-2, blaVIM-4, blaPAO, blaOXA-35, blaOXA-50, blaOXA-395, blaOXA-488, blaOXA-494, blaPER-1 | aph(3′)-IIb, ant(2″)-Ia, aadA6, aph(3′)-Via, aacA4, aadA2, aacA29, strA, strB | catB7, sul1 | [129] | |
| Romania | 10 isolates, ST357, ST395, ST621 | 2018 | blaIMP-13 | aph(3′)-IIb, ant(2″)-I | bcr1, catB7, fosA, sul1 | [130] | |
| Serbia | 4 isolates, CRPA, ST235 (x3), ST654 (x1) | 2018–2021 | blaNDM-1 | aac(6′)Ii, aph(3′)-IIb, aph(6′)Ib, aph(6′)Id, aphA6, aadA6 | sul1 | [131] | |
| Turkey | 2 isolates, MDR, ST308 (x1), ST357 (x1) | 2015–2016 | blaVIM-5, blaIMP-7, blaPAO, blaOXA-2, blaOXA-50, blaOXA-488 | aac(6′)-1Ib-cr, aph(3′)-IIb, aac(6′)-Ib3, aph(3″)-Ib, aph(6)-Id, aac(6′)-II, aadA1 | crpP, crpP-2 | catB7, fosA, sul1 | [132] |
| Country | Isolates Analyzed | Year | β-Lactam Resistance | Aminoglycoside Resistance | Colistin Resistance | Other AMR Determinants | Source |
|---|---|---|---|---|---|---|---|
| Albania | 1 isolate, CRAB, ST2/ST436 | 2015 | ampC, blaOXA-23, blaMBL, blaOXA-51, blaTEM-1 | armA, aph(3′)-Ia, aphA6, strA, and strB | sul2, tetB | [205] | |
| Croatia | 3 isolates, PDR | 2018 | blaOXA-23, blaADC-25, blaOXA-66 | aac(3)-Ia, aph(3′)-Via, aph(3″)-Ib, aph(3′)-Via, aph(6)-Id, armA, aadA1 | pmrB mutations: S14L, A138T, S183F, T269P | catA1, sul1, tet(B) | [200] |
| Greece | 1 isolate, CRAB, ST1/ST439 | 2002 | blaOXA-58 | aphA6, aacA4, aacC1, aphA1 | sul1, tetA | [206] | |
| 42 isolates, (40 x ColR-CRAB, 2 x CRAB) | 2015–2017 | blaADC, blaOXA–51, * blaOXA–23 | Several chromosomal mutations in genes potentially involved in colistin resistance | QRDR mutations: GyrA S83L and ParC S80L | [207] | ||
| Romania | 7 isolates, XDR, ST3636/- (x1), ST492/- (x2), ST1/- (x1), ST636/- (x1), ST2/- (x2) | 2017 | blaOXA-24, blaOXA-23, * blaOXA-23, blaOXA-51,* blaOXA-51, blaOXA-72, blaADC-11, blaADC-25, blaADC-30, blaADC-74, blaTEM-12, blaTEM-84, blaPER-1 | aac(3)-Ia, aph(6)-Id, ant(3″)-IIa, aph(3″)-Ib, aadA1, aph(3′)-Ia, aadA2, armA, aph(3′)-VIa, aph(3′)VIb, armA | catA1, dfrA12, msr(E), mph(E), sul1, sul2, tet(A), tet(B), tetR QRDR mutations: GyrA S83L and ParC S84L, S467G | [208] | |
| 21 isolates, no antimicrobial susceptibility and ST affiliation data | 2018–2019 | blaOXA-23, blaOXA-24, blaTEM, blaVIM, blaVEB | aph(6)-Id, aph(3′)-Via, ant(2″)-Ia, ant(3″)-IIa, armA, aadA1 | msr(E), mph(E), sul1, sul2, tet(B) | [130] | ||
| Serbia | 1 isolate, CRAB | blaOXA-72 | [209] | ||||
| 30 isolates, ColR-CRAB | 2018–2021 | blaNDM-1, blaOXA-23, blaOXA-24, blaADC-30, blaADC-73, blaADC-74, blaADC-217 | aadA2, aph(3′)-VI, aac(3)-Ia, aadA, aph(3″)-Ib, aph(3′)-Ia, armA, ant(3″)-IIc, aph(3′)-Via, aph(6)-Id | Various mutations | catI, dfrA1, dfrA12, msr(E), mph(E), sul1, sul2, tet(B), tetR QRDR mutations: GyrA S84L, V104I, D105E and ParC S81L | [210] | |
| Turkey | 1 isolate, MDR, ST218 | blaADC-73 | [211] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peykov, S.; Strateva, T. Whole-Genome Sequencing-Based Resistome Analysis of Nosocomial Multidrug-Resistant Non-Fermenting Gram-Negative Pathogens from the Balkans. Microorganisms 2023, 11, 651. https://doi.org/10.3390/microorganisms11030651
Peykov S, Strateva T. Whole-Genome Sequencing-Based Resistome Analysis of Nosocomial Multidrug-Resistant Non-Fermenting Gram-Negative Pathogens from the Balkans. Microorganisms. 2023; 11(3):651. https://doi.org/10.3390/microorganisms11030651
Chicago/Turabian StylePeykov, Slavil, and Tanya Strateva. 2023. "Whole-Genome Sequencing-Based Resistome Analysis of Nosocomial Multidrug-Resistant Non-Fermenting Gram-Negative Pathogens from the Balkans" Microorganisms 11, no. 3: 651. https://doi.org/10.3390/microorganisms11030651
APA StylePeykov, S., & Strateva, T. (2023). Whole-Genome Sequencing-Based Resistome Analysis of Nosocomial Multidrug-Resistant Non-Fermenting Gram-Negative Pathogens from the Balkans. Microorganisms, 11(3), 651. https://doi.org/10.3390/microorganisms11030651