
16S-23S rRNA Internal Transcribed Spacer Region (ITS) Sequencing: A Potential Molecular Diagnostic Tool for Differentiating Lactococcus garvieae and Lactococcus petauri

 ,
,  ,
,  ,
,  , , , , , , , , ,
, , , , , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Polymerase Chain Reaction (PCR) of ITS 16S-23S Region
2.3. Phylogenetic Analysis
2.4. Genome Sequencing
2.5. Genetic Identification
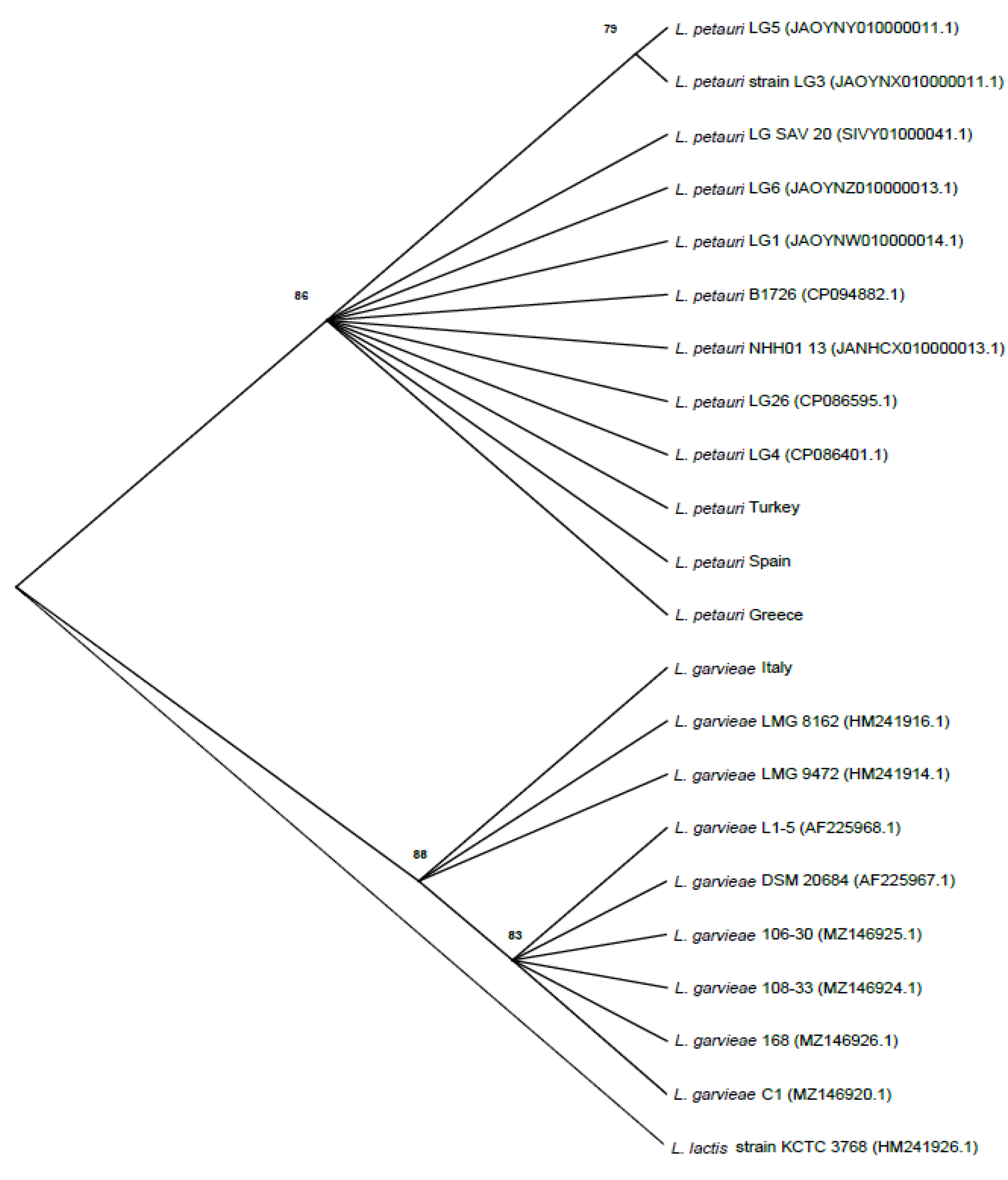
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vendrell, D.; Balcázar, J.L.; Ruiz-Zarzuela, I.; De Blas, I.; Gironés, O.; Múzquiz, J.L. Lactococcus garvieae in fish: A review. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Algöet, M.; Roberts, E.G.; Feist, S.W.; Wheeler, R.W.; Verner-Jeffreys, D.W. Susceptibility of selected freshwater fish species to a UK Lactococcus garvieae isolate. Dis. Aquat. Org. 2009, 36, 227–231. [Google Scholar] [CrossRef]
- Castro, R.; Reguera-Brito, M.; López-Campos, G.H.; Blanco, M.M.; Aguado-Urda, M.; Fernández-Garayzábal, J.F.; Gibello, A. How does temperature influence the development of lactococcosis? Transcriptomic and immunoproteomic in vitro approaches. J. Fish Dis. 2017, 40, 1285–1297. [Google Scholar] [CrossRef]
- Goodman, L.B.; Lawton, M.R.; Franklin-Guild, R.J.; Anderson, R.R.; Schaan, L.; Thachil, A.J.; Wiedmann, M.; Miller, C.B.; Alcaine, S.D.; Kovac, J. Lactococcus petauri sp. nov., isolated from an abscess of a sugar glider. Int. J. Syst. Evol. Microbiol. 2017, 67, 4397–4404. [Google Scholar] [CrossRef]
- Savvidis, K.; Anatoliotis, C.; Kanaki, Z.; Vafeas, G. Epizootic outbreaks of lactococcosis disease in rainbow trout, Oncorhynchus mykiss (Walbaum), culture in Greece. Bull. Eur. Assoc. Fish Pathol. 2007, 27, 223–228. [Google Scholar]
- Shahin, K.; Mukkatira, K.; Yazdi, Z.; Richey, C.; Kwak, K.; Heckman, T.I.; Mohammed, H.H.; Ortega, C.; Avendaño-Herrera, R.; Keleher, B.; et al. Development of a quantitative polymerase chain reaction assay for detection of the aetiological agents of piscine lactococcosis. J. Fish. Dis. 2021, 45, 847–859. [Google Scholar] [CrossRef]
- Egger, R.C.; Câmara Rosa, J.C.; Lara Resende, L.F.; de Pádua, S.B.; de Oliveira Barbosa, F.; Zerbini, M.T.; Tavares, G.C.; Pereira Figueiredo, H.C. Emerging fish pathogens Lactococcus petauri and L. garvieae in Nile tilapia (Oreochromis niloticus) farmed in Brazil. Aquaculture 2023, 565, 739093. [Google Scholar] [CrossRef]
- Assis, G.B.N.; Pereira, F.L.; Zegarra, A.U.; Tavares, G.C.; Leal, C.A.; Figueiredo, H.C.P. Use of MALDI-TOF Mass Spectrometry for the fast identification of Gram-positive fish pathogens. Microbiology 2017, 8, 1492. [Google Scholar] [CrossRef] [PubMed]
- Sciuto, S.; Colli, L.; Fabris, A.; Pastorino, P.; Stoppani, N.; Esposito, G.; Prearo, M.; Esposito, G.; Ajmone-Marsan, P.; Acutis, P.L.; et al. What can genetics do for the control of infectious diseases in aquaculture? Animals 2022, 12, 2176. [Google Scholar] [CrossRef]
- Clarridge, J.E. Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef]
- Zlotkin, A.; Eldar, A.; Ghittino, C.; Bercovier, H. Identification of Lactococcus garvieae by PCR. J. Clin. Microbiol. 1998, 36, 983–985. [Google Scholar] [CrossRef]
- Altinok, I.; Ozturk, R.C.; Ture, M. NGS analysis revealed that Lactococcus garvieae Lg-Per was Lactococcus petauri in Turkey. J. Fish. Dis. 2022, 45, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.J.; Ren, Q.Q.; Fang, S.T.; Wu, J.G.; Jiang, Y.X.; Chen, Y.R.; Zhong, Y.; Wang, D.D.; Zhang, G.X. Complete genome insights into Lactococcus petauri CF11 isolated from a healthy human gut using second- and third-generation sequencing. Front. Genet. 2020, 26, 11–119. [Google Scholar] [CrossRef]
- Kotzamanidis, C.; Malousi, A.; Bitchava, K.; Vafeas, G.; Chatzidinitriou, D.; Skoura, L.; Papadimitriou, E.; Chatzopoulou, F.; Zdragas, A. First report of isolation and genome sequence of L. petauri strain from a rainbow trout lactococcosis outbreak. Curr. Microbiol. 2020, 77, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Quinoo, S.; Coolen, J.P.M.; Van Hijum, S.A.F.T.; Huynen, M.A.; Melches, W.J.G.; van Schaik, W.; Wertheim, H.F.L. Whole-Genome sequencing of bacterial pathogens: The future of nosocomial outbreak analysis. Clin. Microbiol. Rev. 2017, 30, 1015–1063. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; de Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Martinovic, A.; Cabal, A.; Nisic, A.; Sucher, J.; Stöger, A.; Allerberger, F.; Ruppitsch, W. Genome sequences of Lactococcus garvieae and Lactococcus petauri strains isolated from traditional Montenegrin brine cheeses. Microbiol. Resour. Announc. 2021, 10, e0054621. [Google Scholar] [CrossRef]
- Dang, H.T.; Park, H.K.; Myung, S.C.; Kim, W. Development of a novel PCR assay based on the 16S-23S rRNA internal transcribed spacer region for the detection of Lactococcus garvieae. J. Fish. Dis. 2012, 35, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Thanh, H.D.; Park, H.K.; Kim, W.; Shin, H.S. Development of a 16S-23S rRNA intergenic spacer-based quantitative PCR assay for improved detection and enumeration of Lactococcus garvieae. FEMS Microbiol. Lett. 2013, 339, 10–16. [Google Scholar] [CrossRef]
- Pastorino, P.; Colussi, S.; Pizzul, E.; Varello, K.; Menconi, V.; Mugetti, D.; Tomasoni, M.; Esposito, G.; Bertoli, M.; Bozzetta, E.; et al. The unusual isolation of carnobacteria in eyes of healthy salmonids in high-mountain lakes. Sci. Rep. 2021, 11, 2314. [Google Scholar] [CrossRef] [PubMed]
- Kabadjova, P.; Dousset, X.; Le Cam, V.; Prevost, H. Differentiation of closely related Carnobacterium food isolates based on 16S-23S ribosomal DNA intergenic spacer region polymorphism. Appl. Environ. Microbiol. 2002, 68, 5358–5366. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbour-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef]
- Peeters, K.; Willems, A. The gyrB gene is a useful phylogenetic marker for exploring the diversity of Flavobacterium strains isolated from terrestrial and aquatic habitats in Antarctica. FEMS Microbiol. Lett. 2011, 321, 130–140. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Italy | Spain | Greece | Turkey | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain ID | Geographical Origin | Date of Isolation | Strain ID | Geographical Origin | Date of Isolation | Strain ID | Geographical Origin | Date of Isolation | Strain ID | Geographical Origin | Date of Isolation |
| 1-IT | Quinto | 2016 | 5239-VISAVET | Granada | 2019 | LG1 | Macedonia | 2016 | Y-LG1 | Trabzon | 2016 |
| 2-IT | Quinto | 2016 | 5424-VISAVET | Granada | 2017 | 2-ELGO | Macedonia | 2010 | Y2-KTU | Gumushane | 2016 |
| 3-IT | Cassolnovo | 2016 | 5664-VISAVET | Granada | 2017 | LG3 | Macedonia | 2010 | Y3-KTU | Trabzon | 2016 |
| 4-IT | Cassolnovo | 2016 | 5787-VISAVET | Granada | 2016 | 4-ELGO | Macedonia | 2010 | Y6-KTU | Trabzon | 2016 |
| 5-IT | Cerano | 2017 | 02/6071-VISAVET | Granada | 2002 | LG5 | Ipiros | 2009 | Y7-KTU | Trabzon | 2017 |
| 6-IT | Cassolnovo | 2017 | 8666-VISAVET | Granada | 2016 | LG6 | Macedonia | 2008 | K2-KTU | Trabzon | 2017 |
| 7-IT | Cassolnovo | 2018 | 8059-VISAVET | Lérida | 2016 | 7-ELGO | Ipiros | 2009 | K3-KTU | Gumushane | 2016 |
| 8-IT | Cassolnovo | 2018 | 8495-VISAVET | Madrid | 2016 | 8-ELGO | Macedonia | 2009 | K7-KTU | Gumushane | 2018 |
| 9-IT | Cassolnovo | 2018 | 8516-VISAVET | La Coruña | 2016 | 9-ELGO | Ipiros | 2008 | K8-KTU | Gumushane | 2018 |
| 10-IT | Quinto | 2019 | 03/8568-VISAVET | Asturias | 2003 | 10-ELGO | Macedonia | 2009 | Kürtün-KTU | Gumushane | 2020 |
| 11-IT | Quinto | 2019 | 818-VISAVET | Guadalajara | 2016 | 11-ELGO | Macedonia | 2007 | Rize-KTU | Rize | 2020 |
| 12-IT | Cerano | 2019 | 820-VISAVET | Guadalajara | 2016 | 12-ELGO | Macedonia | 2010 | Antalya-KTU | Antalya | 2019 |
| 13-IT | Cassolnovo | 2019 | 393-VISAVET | Granada | 2019 | 13-ELGO | Ipiros | 2010 | Vakfıkebir-KTU | Trabzon | 2020 |
| 14-IT | Quinto | 2020 | 195-VISAVET | La Coruña | 2019 | 14-ELGO | Ipiros | 2010 | LG10 | Mugla | 2017 |
| 15-IT | Preore | 2020 | ICM16/00935 | Granada | 2016 | 15-ELGO | Ipiros | 2008 | 20-KTU | Mugla | 2016 |
| 16-IT | Ormelle | 2017 | 307-VISAVET | Granada | 2016 | 16-ELGO | Ipiros | 2006 | 9-KTU | Kayseri | 2018 |
| 17-IT | Cassolnovo | 2017 | 1008-VISAVET | La Coruña | 2017 | 17-ELGO | Ipiros | 2006 | 13-KTU | Izmir | 2016 |
| 18-IT | Quinto | 2019 | 8831-VISAVET | Lérida | 2017 | 18-ELGO | Macedonia | 2007 | 123-KTU | Izmir | 2017 |
| 19-IT | Cassolnovo | 2019 | P04/8864-VISAVET | Granada | 2004 | 19-ELGO | Macedonia | 2008 | 140-KTU | Elazig | 2016 |
| 20-IT | Quinto | 2019 | 8943-VISAVET | Granada | 2016 | 20-ELGO | Ipiros | 2007 | 107B-KTU | Rize | 2019 |
| 1683 | San Daniele Friuli | 1997 | |||||||||
| 1691-2 | Porcia | 1997 | |||||||||
| 16S-23S Region Position a | |||||||
|---|---|---|---|---|---|---|---|
| 219 | 358 | 440 | 442 | 443 | 469 | 478 | |
| Strain type | |||||||
| L. garvieae DSM20684T | - | G | T | T | T | C | G |
| L. petauri 159469T | A | T | G | A | - | A | T |
| Control strains | |||||||
| L. garvieae Lg.Granada | - | G | T | T | T | C | G |
| L. garvieae Lg 9 | - | G | T | T | T | C | G |
| L. garvieae FDAARGOS_929 | - | G | T | T | T | C | G |
| L. petauri 1001095IJ_161003_C7 | A | T | G | A | - | A | T |
| L. petauri 8831 | A | T | G | A | - | A | T |
| L. petauri D375 | A | T | G | A | - | A | T |
| L. petauri S39 | A | T | G | A | - | A | T |
| Reference strains | |||||||
| L. garvieae LMG 9472 | - | G | T | T | T | C | G |
| L. garvieae LMG 8162 | - | G | T | T | T | C | G |
| L. garvieae C1 | - | G | T | T | T | C | G |
| L. garvieae 108-33 | - | G | T | T | T | C | G |
| L. garvieae 106-30 | - | G | T | T | T | C | G |
| L. garvieae 168 | - | G | T | T | T | C | G |
| L. garvieae 20684 | - | G | T | T | T | C | G |
| L. garvieae L1-5 | - | G | T | T | T | C | G |
| L. petauri LG4 | A | T | G | A | - | A | T |
| L. petauri LG26 | A | T | G | A | - | A | T |
| L. petauri B1726 | A | T | G | A | - | A | T |
| L. petauri NHH01_13 | A | T | G | A | - | A | T |
| L. petauri LG_SAV_20 | A | T | G | A | - | A | T |
| Clinical field strains | |||||||
| Italian (n = 22) b | - | G | T | T | T | C | G |
| Spanish (n = 20) c | A | T | G | A | - | A | T |
| Greek (n = 20) c | A | T | G | A | - | A | T |
| Turkish (n = 20) c | A | T | G | A | - | A | T |
| Isolate | L. petauri 159469T (NZ_MUIZ0000000) | L. garvieae DSM 20684T (JXJV01000001) | Accession Number | ||
|---|---|---|---|---|---|
| %ANI | %DDH | %ANI | %DDH | ||
| Control strains | |||||
| 1001095IJ_161003_C7 | 98.8 | 89.5 | 93.1 | 50.9 | NZ_JADPGL010000010 |
| 8831 | 98.5 | 85.5 | 93.0 | 50.5 | NZ_AFCD01000000 |
| D375 | 98.4 | 85.4 | 93.1 | 50.7 | JAOSHM010000001 |
| S39 | 98.3 | 85.5 | 93.0 | 50.8 | JALDMQ010000000 |
| Lg. Granada | 93.2 | 51.7 | 98.3 | 85.1 | CP084377 |
| Lg 9 | 93.2 | 51.2 | 98.3 | 85.3 | NZ_AGQY00000000 |
| FDAARGOS_929 | 92.9 | 50.8 | 99.9 | 99.9 | CP065637 |
| Field clinical strains | |||||
| ICM16/00935 a | 98.4 | 85.7 | 92.0 | 50.5 | JARHWC000000000 |
| 1683 b | 93.3 | 51.1 | 98.4 | 85.4 | JARHWV000000000 |
| 1691-2 b | 93.3 | 51.1 | 98.4 | 85.4 | JARHWU000000000 |
| Y-LG1 c | 98.4 | 85.6 | 93.0 | 50.8 | JAQPON000000000 |
| LG10 c | 98.4 | 85.5 | 93.1 | 50.8 | JAQPOM000000000 |
| LG6 d | 98.5 | 85.4 | 93.1 | 50.4 | JAOYNZ010000001 |
| LG5 d | 98.4 | 85.4 | 92.4 | 50.4 | JAOYNY010000001 |
| LG3 d | 98.4 | 85.4 | 92.5 | 50.4 | JAOYNX010000001 |
| LG1 d | 98.4 | 85.3 | 92.9 | 50.5 | JAOYNW010000001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoppani, N.; Colussi, S.; Pastorino, P.; Prearo, M.; Sciuto, S.; Altinok, I.; Öztürk, R.Ç.; Ture, M.; Vela, A.I.; Blanco, M.d.M.; et al. 16S-23S rRNA Internal Transcribed Spacer Region (ITS) Sequencing: A Potential Molecular Diagnostic Tool for Differentiating Lactococcus garvieae and Lactococcus petauri. Microorganisms 2023, 11, 1320. https://doi.org/10.3390/microorganisms11051320
Stoppani N, Colussi S, Pastorino P, Prearo M, Sciuto S, Altinok I, Öztürk RÇ, Ture M, Vela AI, Blanco MdM, et al. 16S-23S rRNA Internal Transcribed Spacer Region (ITS) Sequencing: A Potential Molecular Diagnostic Tool for Differentiating Lactococcus garvieae and Lactococcus petauri. Microorganisms. 2023; 11(5):1320. https://doi.org/10.3390/microorganisms11051320
Chicago/Turabian StyleStoppani, Nadia, Silvia Colussi, Paolo Pastorino, Marino Prearo, Simona Sciuto, Ilhan Altinok, Rafet Çağrı Öztürk, Mustafa Ture, Ana Isabel Vela, Maria del Mar Blanco, and et al. 2023. "16S-23S rRNA Internal Transcribed Spacer Region (ITS) Sequencing: A Potential Molecular Diagnostic Tool for Differentiating Lactococcus garvieae and Lactococcus petauri" Microorganisms 11, no. 5: 1320. https://doi.org/10.3390/microorganisms11051320
APA StyleStoppani, N., Colussi, S., Pastorino, P., Prearo, M., Sciuto, S., Altinok, I., Öztürk, R. Ç., Ture, M., Vela, A. I., Blanco, M. d. M., Kotzamanidis, C., Bitchava, K., Malousi, A., Fariano, L., Volpatti, D., Acutis, P. L., & Fernández-Garayzábal, J. F. (2023). 16S-23S rRNA Internal Transcribed Spacer Region (ITS) Sequencing: A Potential Molecular Diagnostic Tool for Differentiating Lactococcus garvieae and Lactococcus petauri. Microorganisms, 11(5), 1320. https://doi.org/10.3390/microorganisms11051320