



Study of Antimicrobial Resistance, Biofilm Formation, and Motility of Pseudomonas aeruginosa Derived from Urine Samples
 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Bacterial Isolates
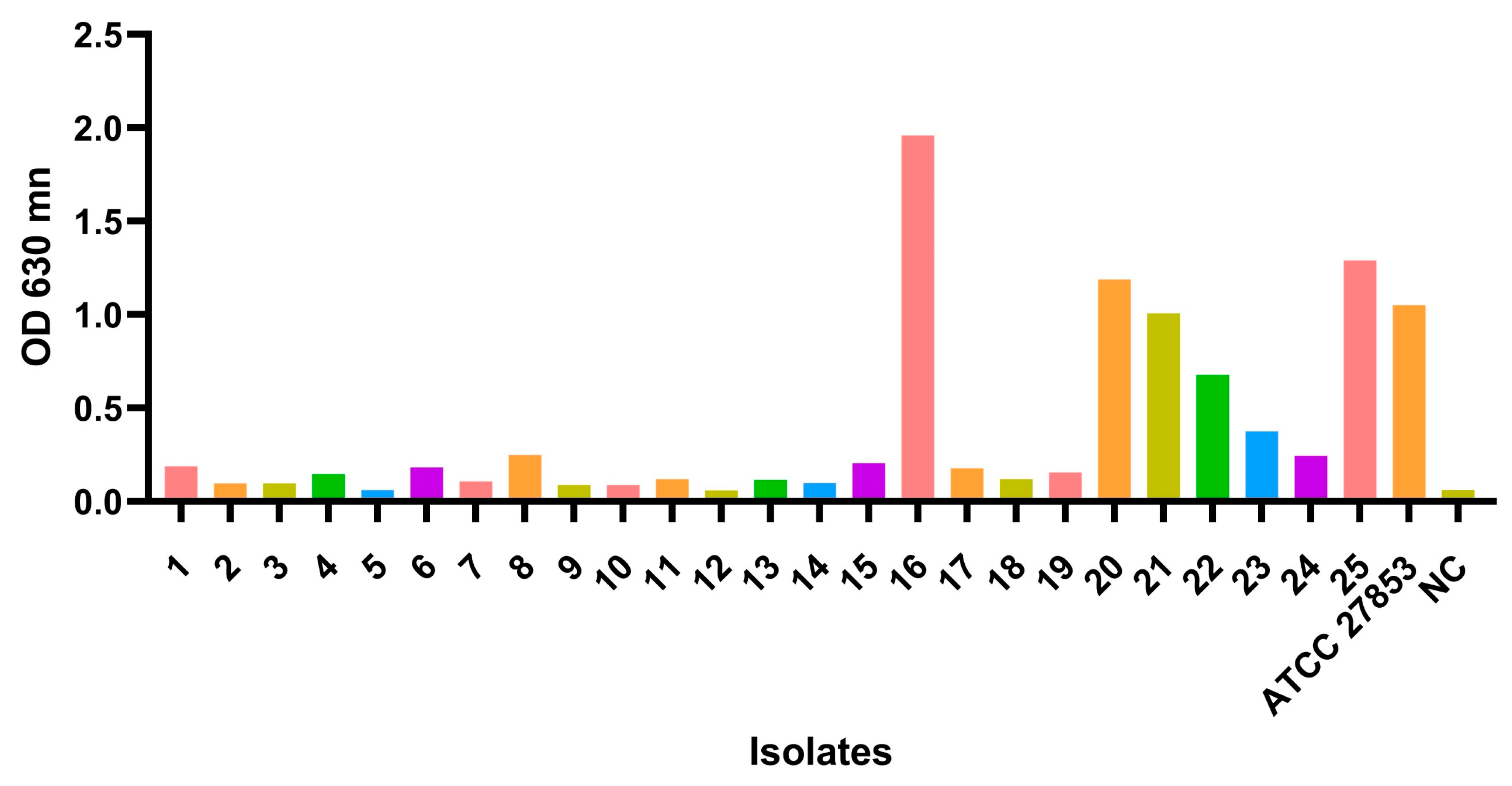
2.2. Biofilm Formation and Biomass Quantification
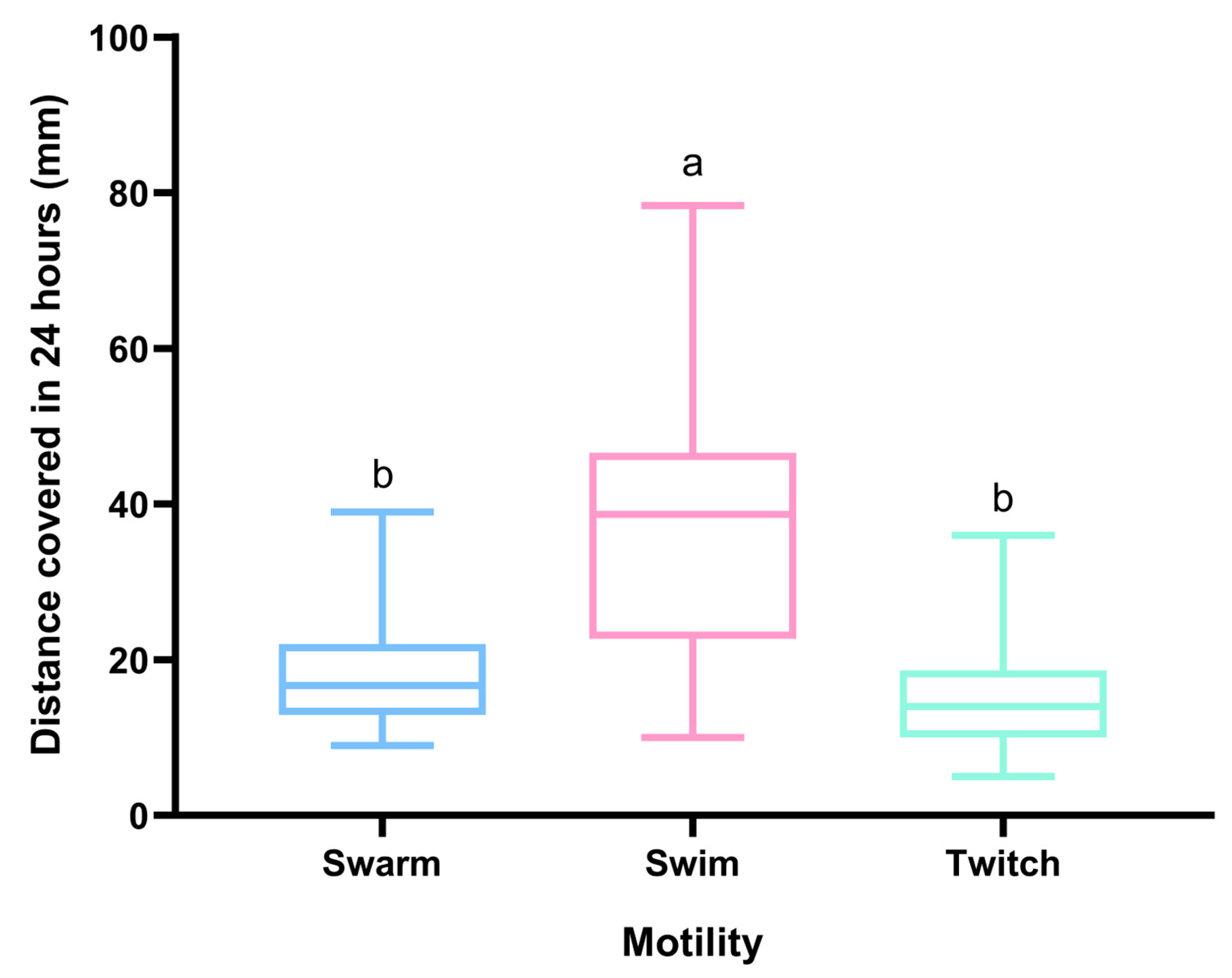
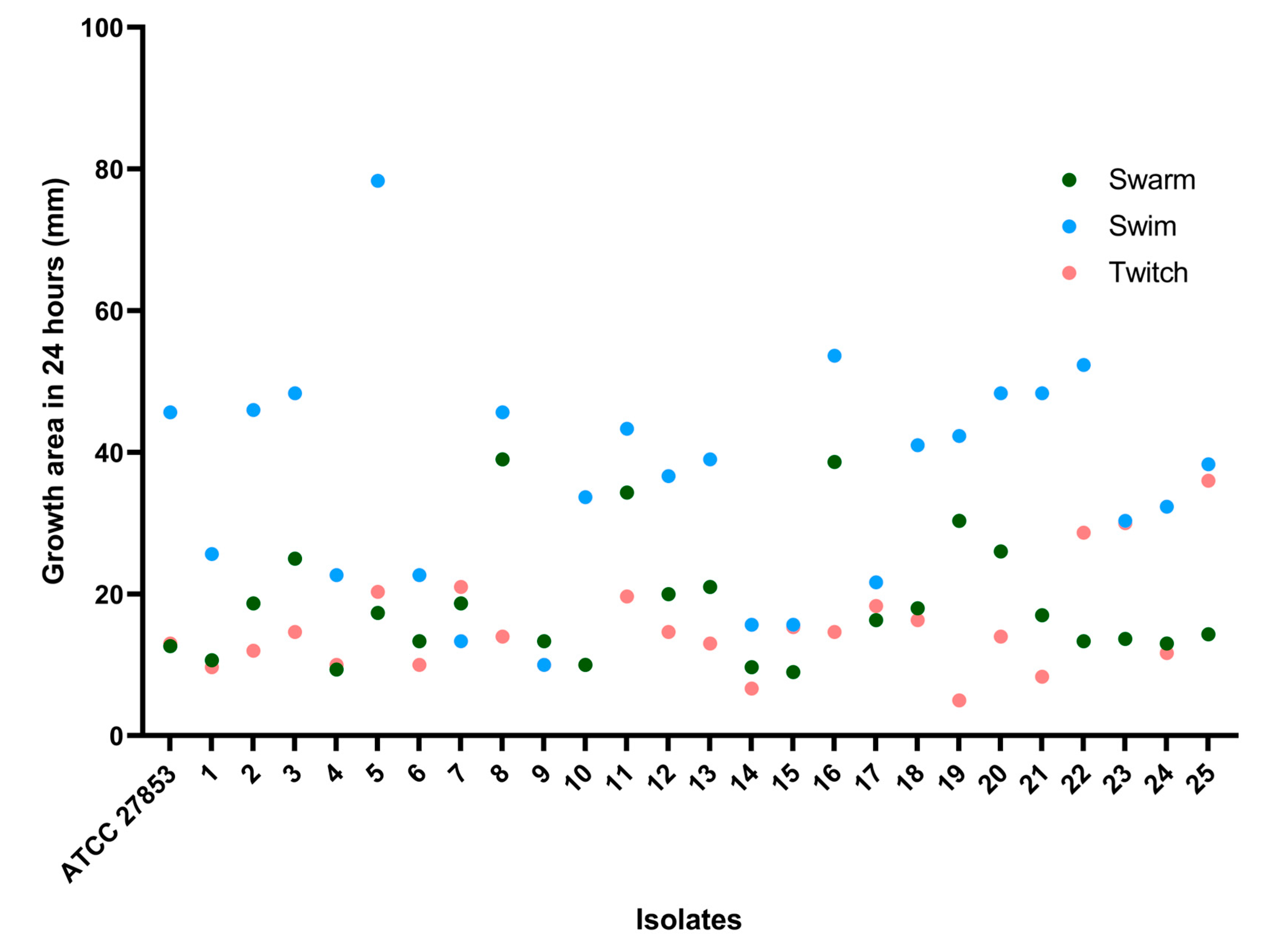
2.3. Motility Assays
2.3.1. Swimming Motility
2.3.2. Swarming Motility
2.3.3. Twitching Motility
2.4. Interpretation of Motility Assays
2.5. Antimicrobial Susceptibility Testing
2.6. DNA Extraction
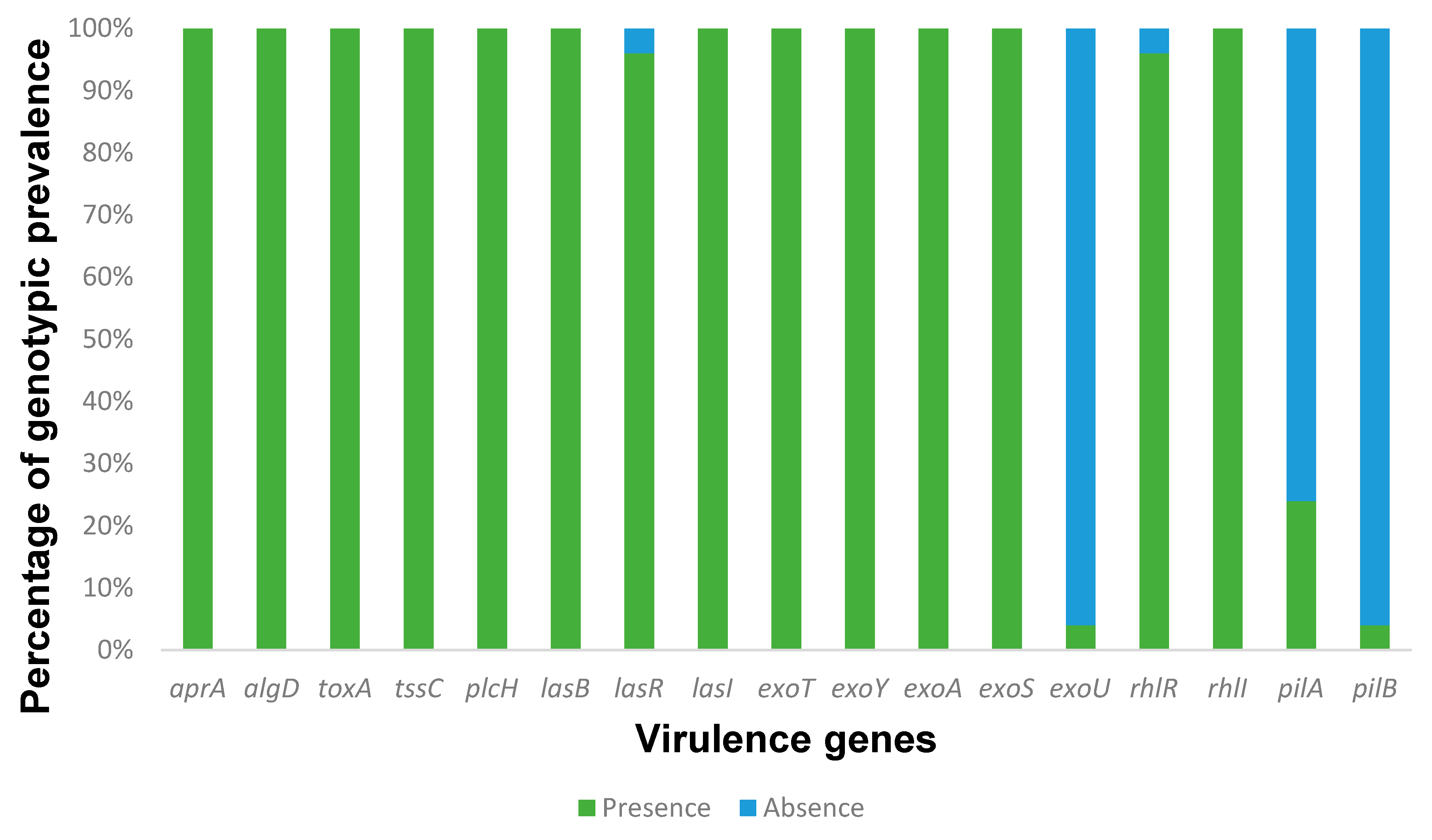
2.7. Antimicrobial Resistance and Virulence Genes
2.8. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anju, V.T.; Busi, S.; Ranganathan, S.; Ampasala, D.R.; Kumar, S.; Suchiang, K.; Kumavath, R.; Dyavaiah, M. Sesamin and sesamolin rescues Caenorhabditis elegans from Pseudomonas aeruginosa infection through the attenuation of quorum sensing regulated virulence factors. Microb. Pathog. 2021, 155, 104912. [Google Scholar] [CrossRef] [PubMed]
- Cullen, L.; McClean, S. Bacterial Adaptation during Chronic Respiratory Infections. Pathogens 2015, 4, 66–89. [Google Scholar] [CrossRef] [PubMed]
- Saeki, E.K.; Yamada, A.Y.; de Araujo, L.A.; Anversa, L.; Garcia, D.O.; de Souza, R.L.B.; Martins, H.M.; Kobayashi, R.K.T.; Nakazato, G. Subinhibitory Concentrations of Biogenic Silver Nanoparticles Affect Motility and Biofilm Formation in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2021, 11, 656984. [Google Scholar] [CrossRef]
- Kipnis, E.; Sawa, T.; Wiener-Kronish, J. Targeting mechanisms of Pseudomonas aeruginosa pathogenesis. Med. Mal. Infect. 2006, 36, 78–91. [Google Scholar] [CrossRef]
- Sawa, T.; Shimizu, M.; Moriyama, K.; Wiener-Kronish, J.P. Association between Pseudomonas aeruginosa type III secretion, antibiotic resistance, and clinical outcome: A review. Crit. Care 2014, 18, 668. [Google Scholar] [CrossRef]
- de Sousa, T.; Hebraud, M.; Dapkevicius, M.; Maltez, L.; Pereira, J.E.; Capita, R.; Alonso-Calleja, C.; Igrejas, G.; Poeta, P. Genomic and Metabolic Characteristics of the Pathogenicity in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 12892. [Google Scholar] [CrossRef] [PubMed]
- Korotkov, K.V.; Sandkvist, M.; Hol, W.G. The type II secretion system: Biogenesis, molecular architecture and mechanism. Nat. Rev. Microbiol. 2012, 10, 336–351. [Google Scholar] [CrossRef]
- Casilag, F.; Lorenz, A.; Krueger, J.; Klawonn, F.; Weiss, S.; Haussler, S. The LasB Elastase of Pseudomonas aeruginosa Acts in Concert with Alkaline Protease AprA To Prevent Flagellin-Mediated Immune Recognition. Infect. Immun. 2016, 84, 162–171. [Google Scholar] [CrossRef]
- Gallique, M.; Bouteiller, M.; Merieau, A. The type VI secretion system: A dynamic system for bacterial communication? Front. Microbiol. 2017, 8, 1454. [Google Scholar] [CrossRef]
- Wood, T.E.; Aksoy, E.; Hachani, A. From welfare to warfare: The arbitration of host-microbiota interplay by the type VI secretion system. Front. Cell. Infect. Microbiol. 2020, 10, 587948. [Google Scholar] [CrossRef]
- Venturi, V. Regulation of quorum sensing in Pseudomonas. FEMS Microbiol. Rev. 2006, 30, 274–291. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wu, G. Can biofilm be reversed through quorum sensing in Pseudomonas aeruginosa? Front. Microbiol. 2019, 10, 1582. [Google Scholar] [CrossRef] [PubMed]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Kohler, T.; Curty, L.K.; Barja, F.; van Delden, C.; Pechere, J.C. Swarming of Pseudomonas aeruginosa is dependent on cell-to-cell signaling and requires flagella and pili. J. Bacteriol. 2000, 182, 5990–5996. [Google Scholar] [CrossRef] [PubMed]
- O’May, C.; Tufenkji, N. The swarming motility of Pseudomonas aeruginosa is blocked by cranberry proanthocyanidins and other tannin-containing materials. Appl. Environ. Microbiol. 2011, 77, 3061–3067. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Behrens, A.-J.; Kaever, V.; Kazmierczak, B.I. Type IV pilus assembly in Pseudomonas aeruginosa over a broad range of cyclic di-GMP concentrations. J. Bacteriol. 2012, 194, 4285–4294. [Google Scholar] [CrossRef] [PubMed]
- Burrows, L.L. Pseudomonas aeruginosa twitching motility: Type IV pili in action. Annu. Rev. Microbiol. 2012, 66, 493–520. [Google Scholar] [CrossRef]
- Okuda, J.; Hayashi, N.; Arakawa, M.; Minagawa, S.; Gotoh, N. Type IV pilus protein PilA of Pseudomonas aeruginosa modulates calcium signaling through binding the calcium-modulating cyclophilin ligand. J. Infect. Chemother. 2013, 19, 653–664. [Google Scholar] [CrossRef]
- Nunn, D.; Bergman, S.; Lory, S. Products of three accessory genes, pilB, pilC, and pilD, are required for biogenesis of Pseudomonas aeruginosa pili. J. Bacteriol. 1990, 172, 2911–2919. [Google Scholar] [CrossRef]
- Olivares, E.; Badel-Berchoux, S.; Provot, C.; Prevost, G.; Bernardi, T.; Jehl, F. Clinical Impact of Antibiotics for the Treatment of Pseudomonas aeruginosa Biofilm Infections. Front. Microbiol. 2019, 10, 2894. [Google Scholar] [CrossRef]
- Strempel, N.; Neidig, A.; Nusser, M.; Geffers, R.; Vieillard, J.; Lesouhaitier, O.; Brenner-Weiss, G.; Overhage, J. Human host defense peptide LL-37 stimulates virulence factor production and adaptive resistance in Pseudomonas aeruginosa. PLoS ONE 2013, 8, e82240. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.D.; Starkey, M.; Kremer, S.; Parsek, M.R.; Wozniak, D.J. Identification of psl, a locus encoding a potential exopolysaccharide that is essential for Pseudomonas aeruginosa PAO1 biofilm formation. J. Bacteriol. 2004, 186, 4466–4475. [Google Scholar] [CrossRef] [PubMed]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, J.; Ju, Z.; Chang, W.; Sun, S. Molecular characterization of antimicrobial resistance in Escherichia coli from rabbit farms in Tai’an, China. BioMed Res. Int. 2018, 2018, 8607647. [Google Scholar] [CrossRef]
- Van De Klundert, J.; Vliegenthart, J. Nomenclature of aminoglycoside resistance genes: A comment. Antimicrob. Agents Chemother. 1993, 37, 927–928. [Google Scholar] [CrossRef]
- Schäfer, E.; Malecki, M.; Tellez-Castillo, C.J.; Pfennigwerth, N.; Marlinghaus, L.; Higgins, P.G.; Mattner, F.; Wendel, A.F. Molecular surveillance of carbapenemase-producing Pseudomonas aeruginosa at three medical centres in Cologne, Germany. Antimicrob. Resist. Infect. Control 2019, 8, 208. [Google Scholar] [CrossRef]
- Capatina, D.; Feier, B.; Hosu, O.; Tertis, M.; Cristea, C. Analytical methods for the characterization and diagnosis of infection with Pseudomonas aeruginosa: A critical review. Anal. Chim. Acta 2022, 1204, 339696. [Google Scholar] [CrossRef]
- Horino, T.; Chiba, A.; Kawano, S.; Kato, T.; Sato, F.; Maruyama, Y.; Nakazawa, Y.; Yoshikawa, K.; Yoshida, M.; Hori, S. Clinical characteristics and risk factors for mortality in patients with bacteremia caused by Pseudomonas aeruginosa. Intern. Med. 2012, 51, 59–64. [Google Scholar] [CrossRef]
- Djordjevic, Z.; Folic, M.M.; Zivic, Z.; Markovic, V.; Jankovic, S.M. Nosocomial urinary tract infections caused by Pseudomonas aeruginosa and Acinetobacter species: Sensitivity to antibiotics and risk factors. Am. J. Infect. Control 2013, 41, 1182–1187. [Google Scholar] [CrossRef]
- Bouza, E.; San Juan, R.; Munoz, P.; Voss, A.; Kluytmans, J.; Infections, C. A European perspective on nosocomial urinary tract infections I. Report on the microbiology workload, etiology and antimicrobial susceptibility (ESGNI–003 study). Clin. Microbiol. Infect. 2001, 7, 523–531. [Google Scholar] [CrossRef]
- Gastmeier, P. Nosocomial urinary tract infections: Many unresolved questions. Clin. Microbiol. Infect. 2001, 7, 521–522. [Google Scholar] [CrossRef] [PubMed]
- Fothergill, J.; Floyd, R.; Newman, J. Invasion and diversity in Pseudomonas aeruginosa urinary tract infections. J. Med. Microbiol. 2022, 71, 001458. [Google Scholar]
- Mittal, R.; Aggarwal, S.; Sharma, S.; Chhibber, S.; Harjai, K. Urinary tract infections caused by Pseudomonas aeruginosa: A minireview. J. Infect. Public Health 2009, 2, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.J.; Records, A.R.; Orr, M.W.; Linden, S.B.; Lee, V.T. Catheter-associated urinary tract infection by Pseudomonas aeruginosa is mediated by exopolysaccharide-independent biofilms. Infect. Immun. 2014, 82, 2048–2058. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.W. Catheter-associated urinary tract infections. Infect. Dis. Clin. N. Am. 1997, 11, 609–622. [Google Scholar] [CrossRef]
- Newman, J.W.; Floyd, R.V.; Fothergill, J.L. The contribution of Pseudomonas aeruginosa virulence factors and host factors in the establishment of urinary tract infections. FEMS Microbiol. Lett. 2017, 364, 15. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A. Microtiter dish biofilm formation assay. JoVE 2011, 47, e2437. [Google Scholar]
- Deziel, E.; Lepine, F.; Milot, S.; Villemur, R. rhlA is required for the production of a novel biosurfactant promoting swarming motility in Pseudomonas aeruginosa: 3-(3-hydroxyalkanoyloxy) alkanoic acids (HAAs), the precursors of rhamnolipids. Microbiology 2003, 149, 2005–2013. [Google Scholar] [CrossRef]
- Tremblay, J.; Richardson, A.P.; Lépine, F.; Déziel, E. Self-produced extracellular stimuli modulate the Pseudomonas aeruginosa swarming motility behaviour. Environ. Microbiol. 2007, 9, 2622–2630. [Google Scholar] [CrossRef]
- Alm, R.A.; Mattick, J.S. Identification of a gene, pilV, required for type 4 fimbrial biogenesis in Pseudomonas aeruginosa, whose product possesses a pre-pilin-like leader sequence. Mol. Microbiol. 1995, 16, 485–496. [Google Scholar] [CrossRef]
- Lee, B.; Haagensen, J.A.; Ciofu, O.; Andersen, J.B.; Høiby, N.; Molin, S. Heterogeneity of biofilms formed by nonmucoid Pseudomonas aeruginosa isolates from patients with cystic fibrosis. J. Clin. Microbiol. 2005, 43, 5247–5255. [Google Scholar] [CrossRef] [PubMed]
- Murray, T.S.; Ledizet, M.; Kazmierczak, B.I. Swarming motility, secretion of type 3 effectors and biofilm formation phenotypes exhibited within a large cohort of Pseudomonas aeruginosa clinical isolates. J. Med. Microbiol. 2010, 59 Pt 5, 511. [Google Scholar] [CrossRef] [PubMed]
- Dashti, A.A.; Jadaon, M.M.; Abdulsamad, A.M.; Dashti, H.M. Heat treatment of bacteria: A simple method of DNA extraction for molecular techniques. Kuwait Med. J. 2009, 41, 117–122. [Google Scholar]
- Amoon, R.H.; Abdallha, A.H.; Sharif, A.O.; Moglad, E.H.; Altyb, H.N.; Elzaki, S.G.; Salih, M.A. Molecular characterization of Pseudomonas aeruginosa isolates from Sudanese patients: A cross-sectional study. F1000Research 2018, 7, 1135. [Google Scholar] [CrossRef]
- Belaaouaj, A.; Lapoumeroulie, C.; Caniça, M.M.; Vedel, G.; Névot, P.; Krishnamoorthy, R.; Paul, G. Nucleotide sequences of the genes coding for the TEM-like β-lactamases IRT-1 and IRT-2 (formerly called TRI-1 and TRI-2). FEMS Microbiol. Lett. 1994, 120, 75–80. [Google Scholar] [PubMed]
- Steward, C.D.; Rasheed, J.K.; Hubert, S.K.; Biddle, J.W.; Raney, P.M.; Anderson, G.J.; Williams, P.P.; Brittain, K.L.; Oliver, A.; McGowan, J.E., Jr. Characterization of clinical isolates of Klebsiella pneumoniae from 19 laboratories using the National Committee for Clinical Laboratory Standards extended-spectrum β-lactamase detection methods. J. Clin. Microbiol. 2001, 39, 2864–2872. [Google Scholar] [CrossRef] [PubMed]
- Pagani, L.; Dell’Amico, E.; Migliavacca, R.; D’Andrea, M.M.; Giacobone, E.; Amicosante, G.; Romero, E.; Rossolini, G.M. Multiple CTX-M-type extended-spectrum β-lactamases in nosocomial isolates of Enterobacteriaceae from a hospital in northern Italy. J. Clin. Microbiol. 2003, 41, 4264–4269. [Google Scholar] [CrossRef]
- Weldhagen, G.F.; Poirel, L.; Nordmann, P. Ambler class A extended-spectrum β-lactamases in Pseudomonas aeruginosa: Novel developments and clinical impact. Antimicrob. Agents Chemother. 2003, 47, 2385–2392. [Google Scholar] [CrossRef]
- Hong, S.S.; Kim, K.; Huh, J.Y.; Jung, B.; Kang, M.S.; Hong, S.G. Multiplex PCR for rapid detection of genes encoding class A carbapenemases. Ann. Lab. Med. 2012, 32, 359–361. [Google Scholar] [CrossRef]
- Neyestanaki, D.K.; Mirsalehian, A.; Rezagholizadeh, F.; Jabalameli, F.; Taherikalani, M.; Emaneini, M. Determination of extended spectrum beta-lactamases, metallo-beta-lactamases and AmpC-beta-lactamases among carbapenem resistant Pseudomonas aeruginosa isolated from burn patients. Burns 2014, 40, 1556–1561. [Google Scholar] [CrossRef]
- Ellington, M.J.; Kistler, J.; Livermore, D.M.; Woodford, N. Multiplex PCR for rapid detection of genes encoding acquired metallo-β-lactamases. J. Antimicrob. Chemother. 2007, 59, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Amudhan, M.S.; Sekar, U.; Kamalanathan, A.; Balaraman, S. blaIMP and blaVIM mediated carbapenem resistance in Pseudomonas and Acinetobacter species in India. J. Infect. Dev. Ctries. 2012, 6, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.R.G.; Caiaffa-Filho, H.H.; Burattini, M.N.; Rossi, F. Metallo-beta-lactamases among imipenem-resistant Pseudomonas aeruginosa in a Brazilian university hospital. Clinics 2010, 65, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Terzi, H.A.; Kulah, C.; Atasoy, A.R.; Ciftci, I.H. Investigation of OprD porin protein levels in carbapenem-resistant Pseudomonas aeruginosa isolates. Jundishapur J. Microbiol. 2015, 8, e25952. [Google Scholar]
- Fazeli, N.; Momtaz, H. Virulence gene profiles of multidrug-resistant Pseudomonas aeruginosa isolated from Iranian hospital infections. Iran. Red Crescent Med. J. 2014, 16, e15722. [Google Scholar] [CrossRef] [PubMed]
- Petit, S.M.-C.; Lavenir, R.; Colinon-Dupuich, C.; Boukerb, A.M.; Cholley, P.; Bertrand, X.; Freney, J.; Doléans-Jordheim, A.; Nazaret, S.; Laurent, F.; et al. Lagooning of wastewaters favors dissemination of clinically relevant Pseudomonas aeruginosa. Res. Microbiol. 2013, 164, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Khattab, M.; Nour, M.; ElSheshtawy, N. Genetic identification of Pseudomonas aeruginosa virulence genes among different isolates. J. Microb. Biochem. Technol. 2015, 7, 274–277. [Google Scholar]
- Zhang, L.; Hinz, A.J.; Nadeau, J.-P.; Mah, T.-F. Pseudomonas aeruginosa tssC1 links type VI secretion and biofilm-specific antibiotic resistance. J. Bacteriol. 2011, 193, 5510–5513. [Google Scholar] [CrossRef]
- Galdino, A.C.M.; Viganor, L.; Ziccardi, M.; Nunes, A.P.F.; Dos Santos, K.R.; Branquinha, M.H.; Santos, A.L. Heterogeneous production of proteases from Brazilian clinical isolates of Pseudomonas aeruginosa. Enferm. Infecc. Y Microbiol. Clin. 2017, 35, 630–637. [Google Scholar] [CrossRef]
- Schaber, J.A.; Carty, N.L.; McDonald, N.A.; Graham, E.D.; Cheluvappa, R.; Griswold, J.A.; Hamood, A.N. Analysis of quorum sensing-deficient clinical isolates of Pseudomonas aeruginosa. J. Med. Microbiol. 2004, 53, 841–853. [Google Scholar] [CrossRef]
- Faraji, F.; Mahzounieh, M.; Ebrahimi, A.; Fallah, F.; Teymournejad, O.; Lajevardi, B. Molecular detection of virulence genes in Pseudomonas aeruginosa isolated from children with Cystic Fibrosis and burn wounds in Iran. Microb. Pathog. 2016, 99, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Ajayi, T.; Allmond, L.R.; Sawa, T.; Wiener-Kronish, J.P. Single-nucleotide-polymorphism mapping of the Pseudomonas aeruginosa type III secretion toxins for development of a diagnostic multiplex PCR system. J. Clin. Microbiol. 2003, 41, 3526–3531. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Bandara, R.; Conibear, T.C.; Thuruthyil, S.J.; Rice, S.A.; Kjelleberg, S.; Givskov, M.; Willcox, M.D. Pseudomonas aeruginosa with lasI quorum-sensing deficiency during corneal infection. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1897–1903. [Google Scholar] [CrossRef] [PubMed]
- Adejobi, A.; Ojo, O.; Alaka, O.; Odetoyin, B.; Onipede, A. Antibiotic resistance pattern of Pseudomonas spp. from patients in a tertiary hospital in South-West Nigeria. Germs 2021, 11, 238. [Google Scholar] [CrossRef]
- Sader, H.S.; Huband, M.D.; Castanheira, M.; Flamm, R.K. Pseudomonas aeruginosa antimicrobial susceptibility results from four years (2012 to 2015) of the international network for optimal resistance monitoring program in the United States. Antimicrob. Agents Chemother. 2017, 61, e02216–e02252. [Google Scholar] [CrossRef]
- Javiya, V.A.; Ghatak, S.B.; Patel, K.R.; Patel, J.A. Antibiotic susceptibility patterns of Pseudomonas aeruginosa at a tertiary care hospital in Gujarat, India. Indian J. Pharmacol. 2008, 40, 230. [Google Scholar]
- Estepa, V.; Rojo-Bezares, B.; Azcona-Gutiérrez, J.M.; Olarte, I.; Torres, C.; Sáenz, Y. Caracterización de mecanismos de resistencia a carbapenémicos en aislados clínicos de Pseudomonas aeruginosa en un hospital español. Enferm. Infecc. Y Microbiol. Clín. 2017, 35, 141–147. [Google Scholar] [CrossRef]
- Katchanov, J.; Asar, L.; Klupp, E.-M.; Both, A.; Rothe, C.; König, C.; Rohde, H.; Kluge, S.; Maurer, F.P. Carbapenem-resistant Gram-negative pathogens in a German university medical center: Prevalence, clinical implications and the role of novel β-lactam/β-lactamase inhibitor combinations. PLoS ONE 2018, 13, e0195757. [Google Scholar] [CrossRef]
- Ahmed, S.; Jakribettu, R.; Kottakutty, S.; Arya, B.; Shakir, V. An emerging multi-drug resistant pathogen in a tertiary care centre in north Kerala. Ann. Biol. Res. 2012, 3, 2794–2799. [Google Scholar]
- Roel, T.; Sahoo, B.; Devi, K.S.; Singh, L.D.; Kamki, Y.; Devi, A.V. Susceptibility Pattern of Bacterial Isolates from Catheterized Patients in a Referral Hospital. J. Dent. Med. Sci. 2014, 13, 18–21. [Google Scholar]
- Anil, C.; Shahid, R.M. Antimicrobial susceptibility patterns of Pseudomonas aeruginosa clinical isolates at a tertiary care hospital in Kathmandu, Nepal. Asian J. Pharm. Clin. Res. 2013, 6, 235–238. [Google Scholar]
- Picão, R.C.; Carrara-Marroni, F.E.; Gales, A.C.; Venâncio, E.J.; Xavier, D.E.; Tognim, M.C.B.; Pelayo, J.S. Metallo-β-lactamase-production in meropenem-susceptible Pseudomonas aeruginosa isolates: Risk for silent spread. Mem. Inst. Oswaldo Cruz 2012, 107, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The emerging NDM carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Nouer, S.A.; Nucci, M.; de-Oliveira, M.P.; Pellegrino, F.L.; Moreira, B.M. Risk factors for acquisition of multidrug-resistant Pseudomonas aeruginosa producing SPM metallo-beta-lactamase. Antimicrob. Agents Chemother. 2005, 49, 3663–3667. [Google Scholar] [CrossRef]
- Azimi, L.; Lari, A.R.; Alaghehbandan, R.; Alinejad, F.; Mohammadpoor, M.; Rahbar, M. KPC-producer gram negative bacteria among burned infants in Motahari Hospital, Tehran: First report from Iran. Ann. Burn. Fire Disasters 2012, 25, 74. [Google Scholar]
- Goli, H.R.; Nahaei, M.R.; Rezaee, M.A.; Hasani, A.; Kafil, H.S.; Aghazadeh, M. Emergence of colistin resistant Pseudomonas aeruginosa at Tabriz hospitals, Iran. Iran. J. Microbiol. 2016, 8, 62. [Google Scholar]
- Sabharwal, N.; Dhall, S.; Chhibber, S.; Harjai, K. Molecular detection of virulence genes as markers in Pseudomonas aeruginosa isolated from urinary tract infections. Int. J. Mol. Epidemiol. Genet. 2014, 5, 125. [Google Scholar]
- Feltman, H.; Schulert, G.; Khan, S.; Jain, M.; Peterson, L.; Hauser, A.R. Prevalence of type III secretion genes in clinical and environmental isolates of Pseudomonas aeruginosa. Microbiology 2001, 147, 2659–2669. [Google Scholar] [CrossRef]
- Mitov, I.; Strateva, T.; Markova, B. Prevalence of virulence genes among bulgarian nosocomial and cystic fibrosis isolates of Pseudomonas aeruginosa. Braz. J. Microbiol. 2010, 41, 588–595. [Google Scholar] [CrossRef]
- Pournajaf, A.; Razavi, S.; Irajian, G.; Ardebili, A.; Erfani, Y.; Solgi, S.; Yaghoubi, S.; Rasaeian, A.; Yahyapour, Y.; Kafshgari, R.; et al. Integron types, antimicrobial resistance genes, virulence gene profile, alginate production and biofilm formation in Iranian cystic fibrosis Pseudomonas aeruginosa isolates. Infez. Med. 2018, 26, 226–236. [Google Scholar]
- Azimi, S.; Kafil, H.S.; Baghi, H.B.; Shokrian, S.; Najaf, K.; Asgharzadeh, M.; Yousefi, M.; Shahrivar, F.; Aghazadeh, M. Presence of exoY, exoS, exoU and exoT genes, antibiotic resistance and biofilm production among Pseudomonas aeruginosa isolates in Northwest Iran. GMS Hyg. Infect. Control 2016, 11, Doc04. [Google Scholar] [PubMed]
- Wolska, K.; Szweda, P. Genetic features of clinical Pseudomonas aeruginosa strains. Pol. J. Microbiol. 2009, 58, 255–260. [Google Scholar] [PubMed]
- Martins, V.V.; Pitondo-Silva, A.; de Melo Manço, L.; Falcão, J.P.; Freitas, S.d.S.; da Silveira, W.D.; Stehling, E.G. Pathogenic potential and genetic diversity of environmental and clinical isolates of Pseudomonas aeruginosa. Apmis 2014, 122, 92–100. [Google Scholar] [CrossRef]
- Bever, R.A.; Iglewski, B.H. Molecular characterization and nucleotide sequence of the Pseudomonas aeruginosa elastase structural gene. J. Bacteriol. 1988, 170, 4309–4314. [Google Scholar] [CrossRef]
- Galdino, A.C.M.; Viganor, L.; De Castro, A.A.; Da Cunha, E.F.; Mello, T.P.; Mattos, L.M.; Pereira, M.D.; Hunt, M.C.; O’Shaughnessy, M.; Howe, O.; et al. Disarming Pseudomonas aeruginosa virulence by the inhibitory action of 1, 10-phenanthroline-5, 6-dione-based compounds: Elastase B (lasB) as a chemotherapeutic target. Front. Microbiol. 2019, 10, 1701. [Google Scholar] [CrossRef] [PubMed]
- Fito-Boncompte, L.; Chapalain, A.; Bouffartigues, E.; Chaker, H.; Lesouhaitier, O.; Gicquel, G.; Bazire, A.; Madi, A.; Connil, N.; Véron, W.; et al. Full virulence of Pseudomonas aeruginosa requires OprF. Infect. Immun. 2011, 79, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.-F. Biofilm-specific antibiotic resistance. Future Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Ra’oof, W.A.M. Distribution of algD, lasB, pilB and nan1 genes among MDR clinical isolates of Pseudomonas aeruginosa in respect to site of infection. Tikrit Med. J. 2011, 17, 148–160. [Google Scholar]
- Asikyan, M.L.; Kus, J.V.; Burrows, L.L. Novel proteins that modulate type IV pilus retraction dynamics in Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 7022–7034. [Google Scholar] [CrossRef] [PubMed]
- Marko, V.A.; Kilmury, S.L.; MacNeil, L.T.; Burrows, L.L. Pseudomonas aeruginosa type IV minor pilins and PilY1 regulate virulence by modulating FimS-AlgR activity. PLoS Pathog. 2018, 14, e1007074. [Google Scholar] [CrossRef]
- Leighton, T.L.; Buensuceso, R.N.; Howell, P.L.; Burrows, L.L. Biogenesis of P seudomonas aeruginosa type IV pili and regulation of their function. Environ. Microbiol. 2015, 17, 4148–4163. [Google Scholar] [CrossRef] [PubMed]
- Pesci, E.C.; Pearson, J.P.; Seed, P.C.; Iglewski, B.H. Regulation of las and rhl quorum sensing in Pseudomonas aeruginosa. J. Bacteriol. 1997, 179, 3127–3132. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.P.; Pesci, E.C.; Iglewski, B.H. Roles of Pseudomonas aeruginosa las and rhl quorum-sensing systems in control of elastase and rhamnolipid biosynthesis genes. J. Bacteriol. 1997, 179, 5756–5767. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.; Koch, G.; Thompson, J.; Xavier, K.; Cool, R.; Quax, W. The multiple signaling systems regulating virulence in Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 2012, 76, 46–65. [Google Scholar] [CrossRef] [PubMed]
- Müsken, M.; Di Fiore, S.; Römling, U.; Häussler, S. A 96-well-plate–based optical method for the quantitative and qualitative evaluation of Pseudomonas aeruginosa biofilm formation and its application to susceptibility testing. Nat. Protoc. 2010, 5, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Abidi, S.H.; Sherwani, S.K.; Siddiqui, T.R.; Bashir, A.; Kazmi, S.U. Drug resistance profile and biofilm forming potential of Pseudomonas aeruginosa isolated from contact lenses in Karachi-Pakistan. BMC Ophthalmol. 2013, 13, 57. [Google Scholar] [CrossRef]
- Saxena, S.; Banerjee, G.; Garg, R.; Singh, M. Comparative study of biofilm formation in Pseudomonas aeruginosa isolates from patients of lower respiratory tract infection. J. Clin. Diagn. Res. 2014, 8, DC09. [Google Scholar]
- Kim, J.; Lee, J.Y.; Lee, H.; Choi, J.Y.; Kim, D.H.; Wi, Y.M.; Peck, K.R.; Ko, K.S. Microbiological features and clinical impact of the type VI secretion system (T6SS) in Acinetobacter baumannii isolates causing bacteremia. Virulence 2017, 8, 1378–1389. [Google Scholar] [CrossRef]
- Heidari, R.; Farajzadeh Sheikh, A.; Hashemzadeh, M.; Farshadzadeh, Z.; Salmanzadeh, S.; Saki, M. Antibiotic resistance, biofilm production ability and genetic diversity of carbapenem-resistant Pseudomonas aeruginosa strains isolated from nosocomial infections in southwestern Iran. Mol. Biol. Rep. 2022, 49, 3811–3822. [Google Scholar] [CrossRef]
- Lima, J.L.d.C.; Alves, L.R.; Jacomé, P.R.L.d.A.; Bezerra Neto, J.P.; Maciel, M.A.V.; Morais, M.M.C.d. Biofilm production by clinical isolates of Pseudomonas aeruginosa and structural changes in LasR protein of isolates non biofilm-producing. Braz. J. Infect. Dis. 2018, 22, 129–136. [Google Scholar] [CrossRef]
- Moskowitz, S.M.; Foster, J.M.; Emerson, J.; Burns, J.L. Clinically feasible biofilm susceptibility assay for isolates of Pseudomonas aeruginosa from patients with cystic fibrosis. J. Clin. Microbiol. 2004, 42, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Dey, P.; Parai, D.; Banerjee, M.; Hossain, S.T.; Mukherjee, S.K. Naringin sensitizes the antibiofilm effect of ciprofloxacin and tetracycline against Pseudomonas aeruginosa biofilm. Int. J. Med. Microbiol. 2020, 310, 151410. [Google Scholar] [CrossRef] [PubMed]
- Deligianni, E.; Pattison, S.; Berrar, D.; Ternan, N.G.; Haylock, R.W.; Moore, J.E.; Elborn, S.J.; Dooley, J.S. Pseudomonas aeruginosa cystic fibrosis isolates of similar RAPD genotype exhibit diversity in biofilm forming ability in vitro. BMC Microbiol. 2010, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Kazmierczak, B.I.; Schniederberend, M.; Jain, R. Cross-regulation of Pseudomonas motility systems: The intimate relationship between flagella, pili and virulence. Curr. Opin. Microbiol. 2015, 28, 78–82. [Google Scholar] [CrossRef]
- Toutain, C.M.; Zegans, M.E.; O’Toole, G.A. Evidence for two flagellar stators and their role in the motility of Pseudomonas aeruginosa. J. Bacteriol. 2005, 187, 771–777. [Google Scholar] [CrossRef]
- Khong, N.Z.-J.; Zeng, Y.; Lai, S.-K.; Koh, C.-G.; Liang, Z.-X.; Chiam, K.-H.; Li, H.-Y. Dynamic swimming pattern of Pseudomonas aeruginosa near a vertical wall during initial attachment stages of biofilm formation. Sci. Rep. 2021, 11, 1952. [Google Scholar] [CrossRef]
- Lin, Q.; Pilewski, J.M.; Di, Y.P. Acidic microenvironment determines antibiotic susceptibility and biofilm formation of Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 747834. [Google Scholar] [CrossRef]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J. Antibiotics as intermicrobial signaling agents instead of weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef]
- Overhage, J.; Bains, M.; Brazas, M.D.; Hancock, R.E. Swarming of Pseudomonas aeruginosa is a complex adaptation leading to increased production of virulence factors and antibiotic resistance. Am. Soc. Microbiol. 2008, 190, 2671–2679. [Google Scholar] [CrossRef]
- Berthelot, R.; Doxsee, K.; Neethirajan, S. Electroceutical approach for impairing the motility of pathogenic bacterium using a microfluidic platform. Micromachines 2017, 8, 207. [Google Scholar] [CrossRef]
- Ho, S.G.; Bureau, L.; John, K.; Débarre, D.; Lecuyer, S. Substrate stiffness impacts early biofilm formation by modulating Pseudomonas aeruginosa twitching motility. bioRxiv 2022, 12, e81112. [Google Scholar]
- Chang, Y.-R.; Weeks, E.R.; Barton, D.; Dobnikar, J.; Ducker, W.A. Effect of topographical steps on the surface motility of the bacterium Pseudomonas aeruginosa. ACS Biomater. Sci. Eng. 2019, 5, 6436–6445. [Google Scholar] [CrossRef] [PubMed]
- Persat, A.; Inclan, Y.F.; Engel, J.N.; Stone, H.A.; Gitai, Z. Type IV pili mechanochemically regulate virulence factors in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2015, 112, 7563–7568. [Google Scholar] [CrossRef]
- Kühn, M.J.; Talà, L.; Inclan, Y.F.; Patino, R.; Pierrat, X.; Vos, I.; Al-Mayyah, Z.; Macmillan, H.; Negrete, J., Jr.; Engel, J.N. Mechanotaxis directs Pseudomonas aeruginosa twitching motility. Proc. Natl. Acad. Sci. USA 2021, 118, e2101759118. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′ → 3′) | Length (bp) | Reference |
|---|---|---|---|
| blaTEM | F: ATTCTTGAAGACGAAAGGGC R: ACGCTCAGTGGAACGAAAAC | 1150 | [45] |
| blaSHV | F: CACTCAAGGATGTATTGTG R: TTAGCGTTGCCAGTGCTCG | 885 | [46] |
| blaCTX | F: CGATGTGCAGTACCAGTAA R: TTAGTGACCAGAATCAGCGG | 585 | [47] |
| blaPER | F: ATGAATGTCATTATAAAAGC R: AATTTGGGCTTAGGGCAGAA | 920 | [48] |
| blaSME | F: ACTTTGATGGGAGGATTGGC R: ACGAATTCGAGCATCACCAG | 551 | [49] |
| blaKPC | F: GTATCGCCGTCTAGTTCTGC R: GGTCGTGTTTCCCTTTAGCC | 638 | [50] |
| blaSmp | F: AAAATCTGGGTACGCAAACG R: ACATTATCCGCTGGAACAGG | 271 | [51] |
| blaVim | F: TTTGGTCGCATATCGCAACG R: CCATTCAGCCAGATCGGCAT | 500 | [52] |
| blaVim-2 | F: AAAGTTATGCCGCACTCACC R: TGCAACTTCATGTTATGCCG | 815 | [53] |
| blaNDM | F: GGTTTGGCGATCTGGTTTTC R: CGGAATGGCTCATCACGATC | 621 | [50] |
| blaOXA | F: CCAAAGACGTGGATG R: GTTAAATTCGACCCCAAGTT | 813 | [46] |
| aac(6′)-Ie-aph(2″)-Ia | F: CCAAGAGCAATAAGGGCATA R: CACTATCATAACCACTACCG | 220 | [25] |
| aph(3′)-IIIa | F: GCCGATGTGGATTGCGAAAA R: GCTTGATCCCCAGTAAGTCA | 292 | [25] |
| aac(3)-II | F: ACTGTGATGGGATACGCGTC R: CTCCGTCAGCGTTTCAGCTA | 237 | [24] |
| aac(3)-III | F: CACAAGAACGTGGTCCGCTA R: AACAGGTAAGCATCCGCATC | 195 | [24] |
| aac(3)-IV | F: CTTCAGGATGGCAAGTTGGT R: TACTCTCGTTCTCCGCTCAT | 286 | [25] |
| ant(4′)-Ia | F: GCAAGGACCGACAACATTTC R: TGGCACAGATGGTCATAACC | 165 | [25] |
| ant(2′)-I | F: ATGTTACGCAGCAGGGCAGTCG R: CGTCAGATCAATATCATCGTGC | 188 | [24] |
| oprD | F: TCCGCAGGTAGCACTCAGTTC R: AAGCCGGATTCATAGGTGGTG | 191 | [54] |
| pilB | F: TCGAACTGATGATCGTGG R: CTTTCGGAGTGAACATCG | 408 | [55] |
| pilA | F: ACAGCATCCAACTGAGCG R: TTGACTTCCTCCAGGCTG | 1675 | [55] |
| aprA | F: ACCCTGTCCTATTCGTTCC R: GATTGCAGCGACAACTTGG | 140 | [56] |
| toxA | F: GGTAACCACGTCAGCCACAT R: TGATGTCCAGGTCATGCTTC | 352 | [57] |
| tssC | F: CTCCAACGACGCGATCAAGT R: TCGGTGTTGTTGACCAGGTA | 150 | [58] |
| plcH | F: GCACGTGGTCATCCTGATGC R: TCCGTAGGCGTCGACGTAC | 608 | [55] |
| lasA | F: GCAGCACAAAAGATCCC R: GAAATGCAGGTGCGGTC | 1075 | [55] |
| lasB | F: GGAATGAACGAAGCGTTCTC R: GGTCCAGTAGTAGCGGTTGG | 284 | [59] |
| lasR | F: CGGGTATCGTACTAGGTGCATCA R: GACGGGAAAGCCAGGAAACTT | 1100 | [56] |
| lasI | F: ATGATCGTACAAATTGGTCGGC R: GTCATGAAACCGCCAGTCG | 605 | [60] |
| exoU | F: ATGCATATCCAATCGTTG R: TCATGTGAACTCCTTATT | 2000 | [56] |
| exoS | F: CGTCGTGTTCAAGCAGATGGTGCTG R: CCGAACCGCTTCACCAGGC | 444 | [61] |
| exoA | F: GACAACGCCCTCAGCATCACCAGC R: CGCTGGCCCATTCGCTCCAGCGCT | 396 | [62] |
| exoY | F: CGGATTCTATGGCAGGGAGG R: GCCCTTGATGCACTCGACCA | 289 | [62] |
| exoT | F: AATCGCCGTCCAACTGCATGCG R: TGTTCGCCGAGGTACTGCTC | 159 | [62] |
| rhlR | F: CAATGAGGAATGACGGAGGC R: GCTTCAGATGAGGCCCAGC | 730 | [60] |
| rhlI | F: CTTGGTCATGATCGAATTGCTC R: ACGGCTGACGACCTCACAC | 625 | [60] |
| rhlA/B | F: TCATGGAATTGTCACAACCGC R: ATACGGCAAAATCATGGCAAC | 151 | [63] |
| algD | F: CGTCTGCCGCGAGATCGGCT R: GACCTCGACGGTCTTGCGGA | 313 | [55] |
| Isolate | Antimicrobial | ||
|---|---|---|---|
| Susceptible | Resistant | Genotype | |
| 1 | TOB, AK, CN, CS | PTZ, CAZ, ATM, CIP, TTC | blaNDM, blaOXA, oprD |
| 2 | TOB, AK, CN, CS | blaOXA, oprD | |
| 3 | TOB, AK, CS | CN | blaOXA, aph(3′)-IIIa, oprD |
| 4 | TOB, AK, CN, CS | LEV | blaNDM, oprD |
| 5 | TOB, AK, CN, CS | blaCTX-M-UN, oprD | |
| 6 | TOB, AK, CN, CS | blaKPC, blaSPM, oprD | |
| 7 | TOB, AK, CN, CS | blaKPC, oprD | |
| 8 | TOB, AK, CN, CS | PTZ, CAZ | blaKPC, blaSPM, oprD |
| 9 | TOB, AK, CN, CS | blaKPC, blaNDM, blaSPM, oprD | |
| 10 | TOB, AK, CN, CS | PTZ, CAZ, ATM, TTC | blaKPC, blaSPM, blaVIM2, oprD |
| 11 | TOB, AK, CS | CN | blaKPC, blaSPM, blaOXA, oprD |
| 12 | TOB, AK, CN, CS | blaCTX-M-UN, blaKPC, oprD | |
| 13 | TOB, AK, CN, CS | blaKPC, oprD | |
| 14 | TOB, AK, CN, CS | PTZ, CAZ, CIP | blaKPC, oprD |
| 15 | TOB, AK, CN, CS | blaKPC, blaSPM, oprD | |
| 16 | TOB, AK, CN, CS | blaSPM, blaVIM, oprD | |
| 17 | TOB, AK, CS | CN | blaKPC, blaNDM, blaSPM, oprD |
| 18 | TOB, AK, CN, CS | blaSPM, blaVIM2, oprD | |
| 19 | TOB, AK, CN, CS | blaKPC, oprD | |
| 20 | TOB, AK, CN, CS | blaKPC,blaSPM, oprD | |
| 21 | TOB, AK, CN, CS | IMI | blaSPM, oprD |
| 22 | TOB, AK, CN, CS | blaSPM, oprD | |
| 23 | TOB, AK, CN, CS | blaSPM, oprD | |
| 24 | TOB, AK, CS | PTZ, CN, TTC, PRL | blaSPM, oprD |
| 25 | TOB, AK, CN, CS | blaSPM, oprD | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa, T.; Hébraud, M.; Alves, O.; Costa, E.; Maltez, L.; Pereira, J.E.; Martins, Â.; Igrejas, G.; Poeta, P. Study of Antimicrobial Resistance, Biofilm Formation, and Motility of Pseudomonas aeruginosa Derived from Urine Samples. Microorganisms 2023, 11, 1345. https://doi.org/10.3390/microorganisms11051345
de Sousa T, Hébraud M, Alves O, Costa E, Maltez L, Pereira JE, Martins Â, Igrejas G, Poeta P. Study of Antimicrobial Resistance, Biofilm Formation, and Motility of Pseudomonas aeruginosa Derived from Urine Samples. Microorganisms. 2023; 11(5):1345. https://doi.org/10.3390/microorganisms11051345
Chicago/Turabian Stylede Sousa, Telma, Michel Hébraud, Olimpia Alves, Eliana Costa, Luís Maltez, José Eduardo Pereira, Ângela Martins, Gilberto Igrejas, and Patricia Poeta. 2023. "Study of Antimicrobial Resistance, Biofilm Formation, and Motility of Pseudomonas aeruginosa Derived from Urine Samples" Microorganisms 11, no. 5: 1345. https://doi.org/10.3390/microorganisms11051345
APA Stylede Sousa, T., Hébraud, M., Alves, O., Costa, E., Maltez, L., Pereira, J. E., Martins, Â., Igrejas, G., & Poeta, P. (2023). Study of Antimicrobial Resistance, Biofilm Formation, and Motility of Pseudomonas aeruginosa Derived from Urine Samples. Microorganisms, 11(5), 1345. https://doi.org/10.3390/microorganisms11051345