
Sporadic Detection of Escherichia coli O104:H4 Strain C227/11Φcu in the Edible Parts of Lamb’s Lettuce Cultured in Contaminated Agricultural Soil Samples



Abstract
:1. Introduction
2. Materials and Methods
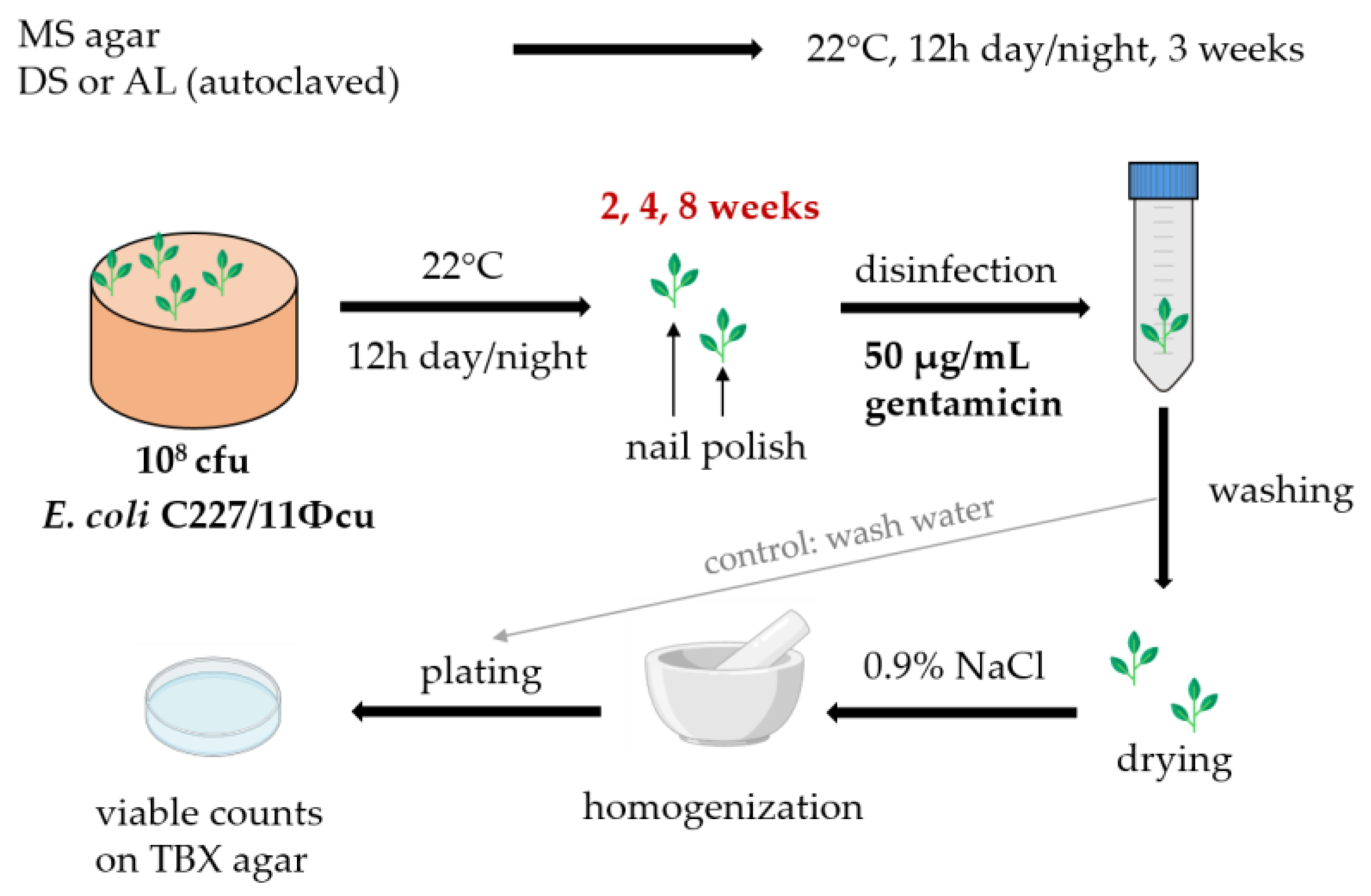
2.1. Bacterial Strain
2.2. Seed Sterilization and Plant Cultivation
2.3. Plant Inoculation
2.4. Plant Harvest
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greig, J.D.; Ravel, A. Analysis of foodborne outbreak data reported internationally for source attribution. Int. J. Food Microbiol. 2009, 130, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Carstens, C.K.; Salazar, J.K.; Darkoh, C. Multistate outbreaks of foodborne illness in the United States associated with fresh produce from 2010 to 2017. Front. Microbiol. 2019, 10, 2667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centres for Disease Control and Prevention (CDC). 2022. National Outbreak Reporting System (NORS). Available online: https://wwwn.cdc.gov/norsdashboard/ (accessed on 26 July 2023).
- Merget, B.; Forbes, K.J.; Brennan, F.; McAteer, S.; Shepherd, T.; Strachan, N.J.C.; Holden, N.J. Influence of plant species, tissue type, and temperature on the capacity of Shiga-toxigenic Escherichia coli to colonize, grow, and be internalized by plants. Appl. Environ. Microbiol. 2019, 85, e00123-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darlison, J.; Mieli, M.; Bengtsson, T.; Hartmann, R.; Mogren, L.; Vågsholm, I.; Karlsson, M.; Alsanius, B.W. Plant species affects the establishment of Escherichia coli O157:H7 gfp+ on leafy vegetables. J. Appl. Microbiol. 2019, 127, 292–305. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.B.; Yaron, S.; Matthews, K.R. Transmission of Escherichia coli O157:H7 from contaminated manure and irrigation water to lettuce plant tissue and its subsequent internalization. Appl. Environ. Microbiol. 2002, 68, 397–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, K.M.; Holden, N.J. Quantification and colonisation dynamics of Escherichia coli O157:H7 inoculation of microgreens species and plant growth substrates. Int. J. Food Microbiol. 2018, 273, 1–10. [Google Scholar] [CrossRef]
- Wright, K.M.; Wright, P.J.; Holden, N.J. Plant species-dependent transmission of Escherichia coli O157:H7 from the spermosphere to cotyledons and first leaves. Environ. Microbiol. Rep. 2022, 14, 926–933. [Google Scholar] [CrossRef]
- Wright, K.M.; Crozier, L.; Marshall, J.; Merget, B.; Holmes, A.; Holden, N.J. Differences in internalization and growth of Escherichia coli O157:H7 within the apoplast of edible plants, spinach and lettuce, compared with the model species Nicotiana benthamiana. Microb. Biotechnol. 2017, 10, 555–569. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Rodríguez, E.; Gundersen, A.; Sbodio, A.; Koike, S.; Suslow, T.V. Evaluation of post-contamination survival and persistence of applied attenuated E. coli O157:H7 and naturally-contaminating E. coli O157:H7 on spinach under field conditions and following postharvest handling. Food Microbiol. 2019, 77, 173–184. [Google Scholar]
- Slayton, R.B.; Turabelidze, G.; Bennett, S.D.; Schwensohn, C.A.; Yaffee, A.Q.; Khan, F.; Butler, C.; Trees, E.; Ayers, T.L.; Davis, M.L. Outbreak of Shiga toxin-producing Escherichia coli (STEC) O157:H7 associated with romaine lettuce consumption, 2011. PLoS ONE 2013, 8, e55300. [Google Scholar] [CrossRef] [Green Version]
- Waltenburg, M.A.; Schwensohn, C.; Madad, A.; Seelman, S.L.; Peralta, V.; Koske, S.E.; Boyle, M.M.; Arends, K.; Patel, K.; Mattioli, M.; et al. Two multistate outbreaks of a reoccurring Shiga toxin-producing Escherichia coli strain associated with romaine lettuce: USA, 2018–2019. Epidemiol. Infect. 2022, 150, e16. [Google Scholar] [CrossRef] [PubMed]
- Holden, N.; Pritchard, L.; Toth, I. Colonization outwith the colon: Plants as an alternative environmental reservoir for human pathogenic enterobacteria. FEMS Microbiol. Rev. 2009, 33, 689–703. [Google Scholar] [CrossRef] [PubMed]
- Bufe, T.; Hennig, A.; Klumpp, J.; Weiss, A.; Nieselt, K.; Schmidt, H. Differential transcriptome analysis of enterohemorrhagic Escherichia coli strains reveals differences in response to plant-derived compounds. BMC Microbiol. 2019, 19, 212. [Google Scholar] [CrossRef] [Green Version]
- Erickson, M.C.; Liao, J.; Payton, A.S.; Webb, C.C.; Ma, L.; Zhang, G.; Flitcroft, I.; Doyle, M.P.; Beuchat, L.R. Fate of Escherichia coli O157:H7 and Salmonella in soil and lettuce roots as affected by potential home gardening practices. J. Sci. Food Agric. 2013, 93, 3841–3849. [Google Scholar] [CrossRef] [PubMed]
- Teplitski, M.; Warriner, K.; Bartz, J.; Schneider, K.R. Untangling metabolic and communication networks: Interactions of enterics with phytobacteria and their implications in produce safety. Trends Microbiol. 2011, 19, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Chitarra, W.; Decastelli, L.; Garibaldi, A.; Gullino, M.L. Potential uptake of Escherichia coli O157:H7 and Listeria monocytogenes from growth substrate into leaves of salad plants and basil grown in soil irrigated with contaminated water. Int. J. Food Microbiol. 2014, 189, 139–145. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Holley, R.A. Factors influencing the microbial safety of fresh produce: A review. Food Microbiol. 2012, 32, 1–19. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Watt, P.M.; Shouse, P.J.; Grieve, C.M. Fate of Escherichia coli O157:H7 in irrigation water on soils and plants as validated by culture method and real-time PCR. Can. J. Microbiol. 2011, 50, 1007–1014. [Google Scholar] [CrossRef]
- Wachtel, M.R.; Whitehand, L.C.; Mandrell, R.E. Association of Escherichia coli O157:H7 with preharvest leaf lettuce upon exposure to contaminated irrigation water. J. Food Prot. 2002, 65, 18–25. [Google Scholar] [CrossRef]
- Renter, D.G.; Sargeant, J.M.; Oberst, R.D.; Samadpour, M. Diversity, frequency, and persistence of Escherichia coli O157 strains from range cattle environments. Appl. Environ. Microbiol. 2003, 69, 542–547. [Google Scholar] [CrossRef] [Green Version]
- Fegan, N.; Vanderlinde, P.; Higgs, G.; Desmarchelier, P. The prevalence and concentration of Escherichia coli O157 in faeces of cattle from different production systems at slaughter. J. Appl. Microbiol. 2004, 97, 362–370. [Google Scholar] [CrossRef] [PubMed]
- McCabe, E.; Burgess, C.M.; Lawal, D.; Whyte, P.; Duffy, G. An investigation of shedding and super-shedding of Shiga toxigenic Escherichia coli O157 and E. coli O26 in cattle presented for slaughter in the Republic of Ireland. Zoonoses Public Health 2019, 66, 83–91. [Google Scholar] [PubMed] [Green Version]
- Shaw, R.K.; Berger, C.N.; Feys, B.; Knutton, S.; Pallen, M.J.; Frankel, G. Enterohemorrhagic Escherichia coli exploits EspA filaments for attachment to salad leaves. Appl. Environ. Microbiol. 2008, 74, 2908–2914. [Google Scholar] [CrossRef] [Green Version]
- Saldaña, Z.; Sánchez, E.; Xicohtencatl-Cortes, J.; Puente, J.L.; Girón, J.A. Surface structures involved in plant stomata and leaf colonization by Shiga-toxigenic Escherichia coli O157: H7. Front. Microbiol. 2011, 2, 119. [Google Scholar] [CrossRef] [Green Version]
- Jechalke, S.; Schierstaedt, J.; Becker, M.; Flemer, B.; Grosch, R.; Smalla, K.; Schikora, A. Salmonella establishment in agricultural soil and colonization of crop plants depend on soil type and plant species. Front. Microbiol. 2019, 10, 967. [Google Scholar] [CrossRef] [Green Version]
- Kroupitski, Y.; Gollop, R.; Belausov, E.; Pinto, R.; Sela Saldinger, S. Salmonella enterica growth conditions influence lettuce leaf internalization. Front. Microbiol. 2019, 10, 639. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C.; Webb, C.C.; Davey, L.E.; Payton, A.S.; Flitcroft, I.D.; Doyle, M.P. Biotic and abiotic variables affecting internalization and fate of Escherichia coli O157:H7 isolates in leafy green roots. J. Food Prot. 2014, 77, 872–879. [Google Scholar] [CrossRef]
- Islam, M.; Doyle, M.P.; Phatak, S.C.; Millner, P.; Jiang, X. Persistence of enterohemorrhagic Escherichia coli O157:H7 in soil and on leaf lettuce and parsley grown in fields treated with contaminated manure composts or irrigation water. J. Food Prot. 2004, 67, 1365–1370. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Gao, C.; Ji, D.; Walker, S.L.; Huang, Q.; Cai, P. Survival of Escherichia coli O157:H7 in various soil particles: Importance of the attached bacterial phenotype. Biol. Fertil. Soils 2017, 53, 209–219. [Google Scholar] [CrossRef]
- NandaKafle, G.; Christie, A.A.; Vilain, S.; Brözel, V.S. Growth and extended survival of Escherichia coli O157:H7 in soil organic matter. Front. Microbiol. 2018, 9, 762. [Google Scholar] [CrossRef] [Green Version]
- Detert, K.; Schmidt, H. Survival of enterohemorrhagic Escherichia coli O104:H4 strain C227/11Φcu in agricultural soils depends on rpoS and environmental factors. Pathogens 2021, 10, 1443. [Google Scholar] [CrossRef] [PubMed]
- Hirneisen, K.A.; Sharma, M.; Kniel, K.E. Human enteric pathogen internalization by root uptake into food crops. Foodborne Pathog. Dis. 2012, 9, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Kutter, S.; Hartmann, A.; Schmid, M. Colonization of barley (Hordeum vulgare) with Salmonella enterica and Listeria spp. FEMS Microbiol. Ecol. 2006, 56, 262–271. [Google Scholar] [CrossRef]
- Bernstein, N.; Sela, S.; Pinto, R.; Ioffe, M. Evidence for internalization of Escherichia coli into the aerial parts of maize via the root system. J. Food Prot. 2007, 70, 471–475. [Google Scholar] [CrossRef]
- Franz, E.; Visser, A.A.; Van Diepeningen, A.D.; Klerks, M.M.; Termorshuizen, A.J.; van Bruggen, A.H.C. Quantification of contamination of lettuce by GFP-expressing Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium. Food Microbiol. 2007, 24, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Ingram, D.T.; Patel, J.R.; Millner, P.D.; Wang, X.; Hull, A.E.; Donnenberg, M.S. A novel approach to investigate the uptake and internalization of Escherichia coli O157:H7 in spinach cultivated in soil and hydroponic medium. J. Food Prot. 2009, 72, 1513–1520. [Google Scholar] [CrossRef]
- Erickson, M.C.; Webb, C.C.; Diaz-Perez, J.C.; Phatak, S.C.; Silvoy, J.J.; Davey, L.; Payton, A.S.; Liao, J.; Ma, L.; Doyle, M.P. Infrequent internalization of Escherichia coli O157:H7 into field-grown leafy greens. J. Food Prot. 2010, 73, 500–506. [Google Scholar] [CrossRef]
- Eißenberger, K.; Moench, D.; Drissner, D.; Weiss, A.; Schmidt, H. Adherence factors of enterohemorrhagic Escherichia coli O157:H7 strain Sakai influence its uptake into the roots of Valerianella locusta grown in soil. Food Microbiol. 2018, 76, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Esmael, A.; Al-Hindi, R.R.; Albiheyri, R.S.; Alharbi, M.G.; Filimban, A.A.R.; Alseghayer, M.S.; Almaneea, A.M.; Alhadlaq, M.A.; Ayubu, J.; Teklemariam, A.D. Fresh produce as a potential vector and reservoir for human bacterial pathogens: Revealing the ambiguity of interaction and transmission. Microorganisms 2023, 11, 753. [Google Scholar] [CrossRef]
- Mellmann, A.; Harmsen, D.; Cummings, C.A.; Zentz, E.B.; Leopold, S.R.; Rico, A.; Prior, K.; Szczepanowski, R.; Ji, Y.; Zhang, W.; et al. Prospective genomic characterization of the german enterohemorrhagic Escherichia coli O104:H4 outbreak by rapid next generation sequencing technology. PLoS ONE 2011, 6, e22751. [Google Scholar] [CrossRef]
- Zangari, T.; Melton-Celsa, A.R.; Panda, A.; Boisen, N.; Smith, M.A.; Tatarov, I.; De Tolla, L.J.; Nataro, J.P.; O’Brien, A.D. Virulence of the Shiga toxin type 2-expressing Escherichia coli O104:H4 german outbreak isolate in two animal models. Infect. Immun. 2013, 81, 1562–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knödler, M.; Berger, M.; Dobrindt, U. Long-term survival of the Shiga toxin-producing Escherichia coli O104: H4 outbreak strain on fenugreek seeds. Food Microbiol. 2016, 59, 190–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eißenberger, K.; Drissner, D.; Walsh, F.; Weiss, A.; Schmidt, H. Plant variety and soil type influence Escherichia coli O104:H4 strain C227/11ϕcu adherence to and internalization into the roots of lettuce plants. Food Microbiol. 2020, 86, 103316. [Google Scholar] [CrossRef]
- Tuttle, J.; Gomez, T.; Doyle, M.P.; Wells, J.G.; Zhao, T.; Tauxe, R.V.; Griffin, P.M. Lessons from a large outbreak of Escherichia coli O157:H7 infections: Insights into the infectious dose and method of widespread contamination of hamburger patties. Epidemiol. Infect. 1999, 122, 185–192. [Google Scholar] [CrossRef]
- Schreiter, S.; Ding, G.-C.; Heuer, H.; Neumann, G.; Sandmann, M.; Grosch, R.; Kropf, S.; Smalla, K. Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce. Front Microbiol. 2014, 5, 144. [Google Scholar] [CrossRef]
- Schierstaedt, J.; Jechalke, S.; Nesme, J.; Neuhaus, K.; Sørensen, S.J.; Grosch, R.; Smalla, K.; Schikora, A. Salmonella persistence in soil depends on reciprocal interactions with indigenous microorganisms. Environ. Microbiol. 2020, 22, 2639–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Overbeek, L.; Duhamel, M.; Aanstoot, S.; van der Plas, C.L.; Nijhuis, E.; Poleij, L.; Russ, L.; van der Zouwen, P.; Andreo-Jimenez, B. Transmission of Escherichia coli from manure to root zones of field-grown lettuce and leek plants. Microorganisms 2021, 9, 2289. [Google Scholar] [CrossRef]
- Underthun, K.; De, J.; Gutierrez, A.; Silverberg, R.; Schneider, K.R. Survival of Salmonella and Escherichia coli in two different soil types at various moisture levels and temperatures. J. Food Prot. 2018, 81, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.A.; Lee, S.; De, J.; Jeong, K.C.; Schneider, K.R. Survival of Escherichia coli O157 in autoclaved and natural sandy soil mesocosms. PLoS ONE 2020, 15, e0234562. [Google Scholar] [CrossRef]
- Klerks, M.M.; Franz, E.; van Gent-Pelzer, M.; Zijlstra, C.; van Bruggen, A.H.C. Differential interaction of Salmonella enterica serovars with lettuce cultivars and plant-microbe factors influencing the colonization efficiency. ISME J. 2007, 1, 620–631. [Google Scholar] [CrossRef] [Green Version]
- Mootian, G.; Wu, W.-H.; Matthews, K.R. Transfer of Escherichia coli O157:H7 from soil, water, and manure contaminated with low numbers of the pathogen to lettuce plants. J. Food Prot. 2009, 72, 2308–2312. [Google Scholar] [CrossRef] [PubMed]
- van Overbeek, L.S.; Lombaers-van der Plas, C.; van der Zouwen, P. The role of pea (Pisum sativum) seeds in the transmission of entero-aggregative Escherichia coli to growing plants. Microorganisms 2020, 8, 1271. [Google Scholar] [CrossRef]
- Cooley, M.B.; Miller, W.G.; Mandrell, R.E. Colonization of Arabidopsis thaliana with Salmonella enterica and enterohemorrhagic Escherichia coli O157:H7 and competition by Enterobacter asburiae. Appl. Environ. Microbiol. 2003, 69, 4915–4926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honjoh, K.; Mishima, T.; Kido, N.; Shimamoto, M.; Miyamoto, T. Investigation of routes of Salmonella contamination via soils and the use of mulch for contamination control during lettuce cultivation. Food Sci. Technol. Res. 2014, 20, 961–969. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Time Point | C227/11Φcu-Positive Samples * (n) | CFU/g Lamb’s Lettuce ** | ||
|---|---|---|---|---|
| Sample 1 | Sample 2 | |||
| MS agar | 2 weeks | 2 (4) | 3.6 × 103 | 1.1 × 103 |
| 4 weeks | 2 (4) | 2.5 × 101 | 1.5 × 102 | |
| 8 weeks | 0 (4) | n.d. | n.d. | |
| Alluvial loam (AL) | 2 weeks | 0 (4) | n.d. | n.d. |
| 4 weeks | 2 (4) | 1.3 × 101 | 1.3 × 101 | |
| 8 weeks | 1 (4) | 6.9 × 103 | n.d. | |
| Diluvial sand (DS) | 2 weeks | 1 (4) | 2.5 × 101 | n.d. |
| 4 weeks | 0 (4) | n.d. | n.d. | |
| 8 weeks | 1 (4) | 2.0 × 103 | n.d. | |
| Soil Type/Agar | Samples | Positive Samples | Recovery Rate |
|---|---|---|---|
| MS | 22 | 6 | 27% |
| AL | 22 | 3 | 13.5% |
| DS | 22 | 2 | 9% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Detert, K.; Schmidt, H. Sporadic Detection of Escherichia coli O104:H4 Strain C227/11Φcu in the Edible Parts of Lamb’s Lettuce Cultured in Contaminated Agricultural Soil Samples. Microorganisms 2023, 11, 2072. https://doi.org/10.3390/microorganisms11082072
Detert K, Schmidt H. Sporadic Detection of Escherichia coli O104:H4 Strain C227/11Φcu in the Edible Parts of Lamb’s Lettuce Cultured in Contaminated Agricultural Soil Samples. Microorganisms. 2023; 11(8):2072. https://doi.org/10.3390/microorganisms11082072
Chicago/Turabian StyleDetert, Katharina, and Herbert Schmidt. 2023. "Sporadic Detection of Escherichia coli O104:H4 Strain C227/11Φcu in the Edible Parts of Lamb’s Lettuce Cultured in Contaminated Agricultural Soil Samples" Microorganisms 11, no. 8: 2072. https://doi.org/10.3390/microorganisms11082072
APA StyleDetert, K., & Schmidt, H. (2023). Sporadic Detection of Escherichia coli O104:H4 Strain C227/11Φcu in the Edible Parts of Lamb’s Lettuce Cultured in Contaminated Agricultural Soil Samples. Microorganisms, 11(8), 2072. https://doi.org/10.3390/microorganisms11082072