The Role of Glycans in Bacterial Adhesion to Mucosal Surfaces: How Can Single-Molecule Techniques Advance Our Understanding?

 , ,
, ,
Abstract
:1. Introduction
2. Sensitive Force Probes and Their Use for Single Molecule and Single Cell Interaction Studies
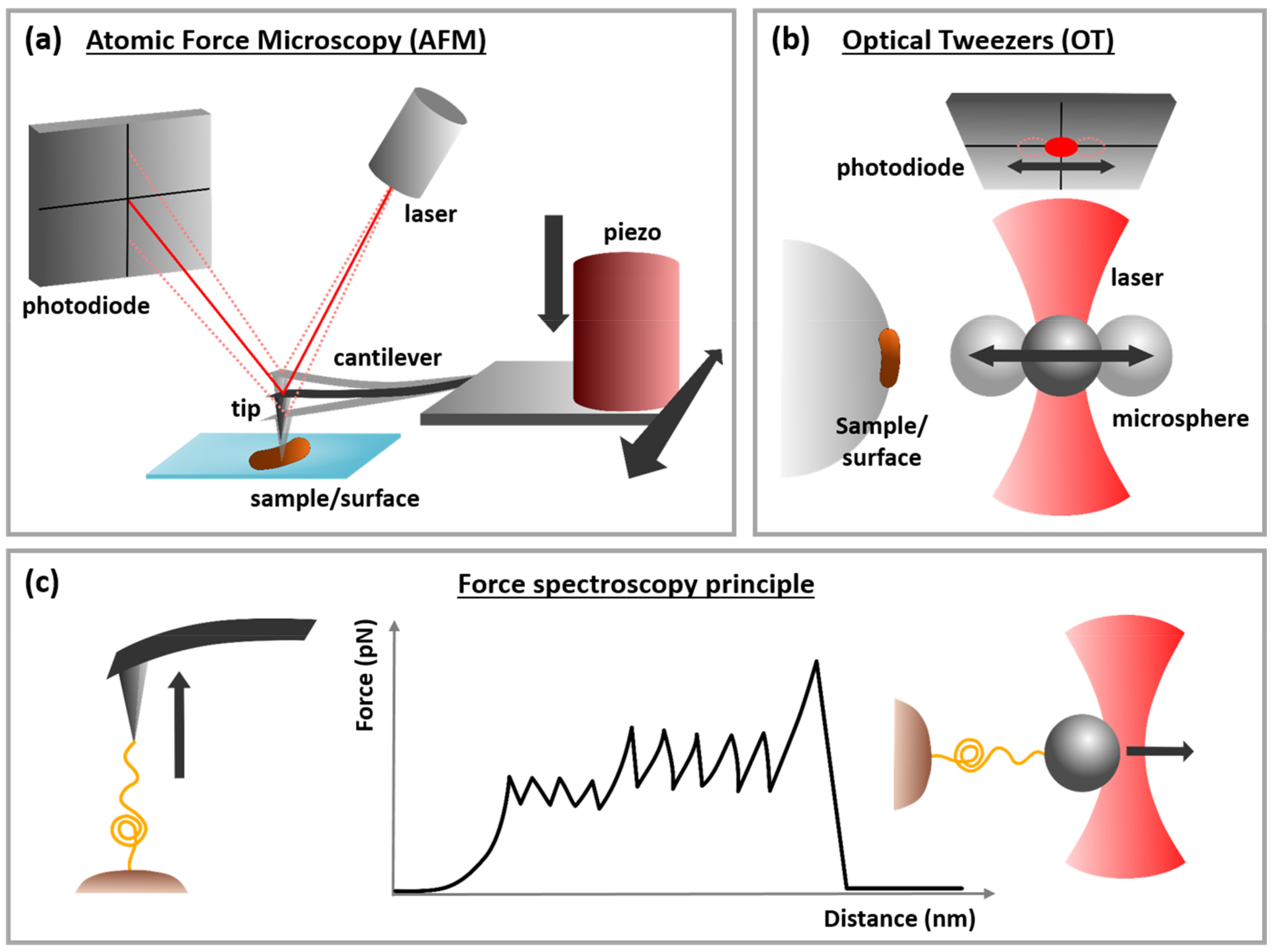
2.1. Optical Tweezers (OT): Working Principle and Applicability for Studies of the Mechanisms Underlying Microbial Adhesion
2.2. Atomic Force Microscopy (AFM): Working Principle and Applicability for Studies of the Mechanisms Underlying Microbial Adhesion
2.3. Strategies Used to Functionalize Surfaces Used in OT or AFM Studies
3. Mechanisms of Glycan-Based Adhesion of Bacteria
3.1. General Considerations about the Mechanisms Underlying Microbial Adhesion to Mucosal Surfaces
3.2. Pili-Based Attachment to Mucosal Surfaces
3.3. Non Pili-Based Interactions
3.4. Glycan-Glycan Interactions
3.5. The Importance of Glycan Patches for Bacterial Adhesion
3.6. Comparison of the Strength of Bacterial Surface Adhesion with Single Molecular Pair Interactions
4. Concluding Remarks
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Tytgat, H.L.P.; de Vos, W.M. Sugar Coating the Envelope: Glycoconjugates for Microbe-Host Crosstalk. Trends Microbiol. 2016, 24, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, C.; Chemla, Y.R.; Forde, N.R.; Izhaky, D. Mechanical Processes in Biochemistry. Annu. Rev. Biochem. 2004, 73, 705–748. [Google Scholar] [CrossRef] [PubMed]
- Dufrêne, Y.F. Sticky microbes: Forces in microbial cell adhesion. Trends Microbiol. 2015, 23, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Ashkin, A. Acceleration and trapping of particles by radiation pressure. Phys. Rev. Lett. 1970, 24, 156–159. [Google Scholar] [CrossRef]
- Ashkin, A. Optical trapping and manipulation of neutral particles using lasers. Proc. Natl. Acad. Sci. USA 1997, 94, 4853–4860. [Google Scholar] [CrossRef] [PubMed]
- Ashkin, A. Forces of a single-beam gradient laser trap on a dielectric sphere in the ray optics regime. Biophys. J. 1992, 61, 569–582. [Google Scholar] [CrossRef]
- Wang, M.D.; Yin, H.; Landick, R.; Gelles, J.; Block, S.M. Stretching DNA with optical tweezers. Biophys. J. 1997, 72, 1335–1346. [Google Scholar] [CrossRef]
- Kellermayer, M.S. Folding-Unfolding Transitions in Single Titin Molecules Characterized with Laser Tweezers. Science 1997, 276, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Jass, J.; Schedin, S.; Fallman, E.; Ohlsson, J.; Nilsson, U.J.; Uhlin, B.E.; Axner, O. Physical properties of Escherichia coli P pili measured by optical tweezers. Biophys. J. 2004, 87, 4271–4283. [Google Scholar] [CrossRef] [PubMed]
- Castelain, M.; Rouxhet, P.G.; Pignon, F.; Magnin, A.; Piau, J.-M. Single-cell adhesion probed in-situ using optical tweezers: A case study with Saccharomyces cerevisiae. J. Appl. Phys. 2012, 111, 114701. [Google Scholar] [CrossRef]
- Simpson, K.H.; Bowden, G.; Hook, M.; Anvari, B. Measurement of adhesive forces between individual Staphylococcus aureus MSCRAMMs and protein-coated surfaces by use of optical tweezers. J. Bacteriol. 2003, 185, 2031–2035. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.H.; Bowden, M.G.; Hook, M.; Anvari, B. Measurement of adhesive forces between S-epidermidis and fibronectin-coated surfaces using optical tweezers. Lasers Surg. Med. 2002, 31, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.H.; Bowden, M.G.; Peacock, S.J.; Arya, M.; Höök, M.; Anvari, B. Adherence of Staphylococcus aureus fibronectin binding protein A mutants: An investigation using optical tweezers. Biomol. Eng. 2004, 21, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Björnham, O.; Fallman, E.; Axner, O.; Ohlsson, J.; Nilsson, U.J.; Boren, T.; Schedin, S. Measurements of the binding force between the Helicobacter pylori adhesin BabA and the Lewis b blood group antigen using optical tweezers. J. Biomed. Opt. 2005, 10, 44024. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Axner, O.; Uhlin, B.E.; Fällman, E. Characterization of the Mechanical Properties of Fimbrial Structures by Optical Tweezers; Hinterdorfer, P., Schutz, G., Pohl, P., Eds.; Trauner: New York, NY, USA, 2006; pp. 19–22. [Google Scholar]
- Castelain, M.; Pignon, F.; Piau, J.-M.; Magnin, A.; Mercier-Bonin, M.; Schmitz, P. Removal forces and adhesion properties of Saccharomyces cerevisiae on glass substrates probed by optical tweezer. J. Chem. Phys. 2007, 127, 10B602. [Google Scholar] [CrossRef] [PubMed]
- Axner, O.; Björnham, O.; Castelain, M.; Koutris, E.; Schedin, S.; Fällman, E.; Andersson, M. Unraveling the Secrets of Bacterial Adhesion Organelles Using Single-Molecule Force Spectroscopy. In Single Molecule Spectroscopy in Chemistry, Physics and Biology; Gräslund, A., Rigler, R., Widengren, J., Eds.; Springer Series in Chemical Physics; Springer: Berlin/Heidelberg, Germany, 2010; Volume 96, pp. 337–362. ISBN 978-3-642-02597-6. [Google Scholar]
- Axner, O.; Andersson, M.; Bjornham, O.; Castelain, M.; Klinth, J.; Koutris, E.; Schedin, S. Assessing bacterial adhesion on an individual adhesin and single pili level using optical tweezers. Adv. Exp. Med. Biol. 2011, 715, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Björnham, O.; Bugaytsova, J.; Boren, T.; Schedin, S. Dynamic force spectroscopy of the Helicobacter pylori BabA-Lewis b binding. Biophys. Chem. 2009, 143, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Bell, G.I. Models for the specific adhesion of cells to cells. Science 1978, 200, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.; Ritchie, K. Dynamic strength of molecular adhesion bonds. Biophys. J. 1997, 72, 1541–1555. [Google Scholar] [CrossRef]
- Evans, E. Energy landscapes of biomolecular adhesion and receptor anchoring at interfaces explored with dynamic force spectroscopy. Faraday Discuss. 1998, 111, 1–16. [Google Scholar] [CrossRef]
- Evans, E. Looking inside molecular bonds at biological interfaces with dynamic force spectroscopy. Biophys. Chem. 1999, 82, 83–97. [Google Scholar] [CrossRef]
- Evans, E. Probing the Relation between Force-Lifetime—And Chemistry in Single Molecular Bonds. Annu. Rev. Biophys. Biomem. 2001, 30, 105–128. [Google Scholar] [CrossRef] [PubMed]
- Merkel, R.; Nassoy, P.; Leung, A.; Ritchie, K.; Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature 1999, 397, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Neuert, G.; Albrecht, C.; Pamir, E.; Gaub, H.E. Dynamic force spectroscopy of the digoxigenin-antibody complex. FEBS Lett. 2006, 580, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Liphardt, J. Reversible Unfolding of Single RNA Molecules by Mechanical Force. Science 2001, 292, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, C.; Shank, E.A.; Bustamante, C.; Marqusee, S. Direct observation of the three-state folding of a single protein molecule. Science 2005, 309, 2057–2060. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ma, L.; Zhang, Y. High-Resolution Optical Tweezers for Single-Molecule Manipulation. Yale J Biol. Med. 2013, 86, 367–383. [Google Scholar] [PubMed]
- Ma, L.; Cai, Y.; Li, Y.; Jiao, J.; Wu, Z.; O’Shaughnessy, B.; De Camilli, P.; Karatekin, E.; Zhang, Y. Single-molecule force spectroscopy of protein-membrane interactions. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Flyvbjerg, H.; Jülicher, F.; Ormos, P.; David, F. Physics of Bio-Molecules and Cells: Les Houches Session LXXV, 2–27 July 2001; Springer Science & Business Media: Berlin, Germany, 2003; ISBN 978-3-540-45701-5. [Google Scholar]
- Björnham, O.; Andersson, M. Theory for nonlinear dynamic force spectroscopy. Eur. Biophys. J. 2017, 46, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Pillet, F.; Chopinet, L.; Formosa, C.; Dague, É. Atomic Force Microscopy and pharmacology: From microbiology to cancerology. Biochim. Biophys. Acta 2014, 1840, 1028–1050. [Google Scholar] [CrossRef] [PubMed]
- Castelain, M.; Duviau, M.-P.; Canette, A.; Schmitz, P.; Loubiere, P.; Cocaign-Bousquet, M.; Piard, J.-C.; Mercier-Bonin, M. The Nanomechanical Properties of Lactococcus lactis Pili Are Conditioned by the Polymerized Backbone Pilin. PLoS ONE 2016, 11, e0152053. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W. High-resolution imaging of the microbial cell surface. J. Microbiol. 2016, 54, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Rheinlaender, J.; Gräbner, A.; Ott, L.; Burkovski, A.; Schäffer, T.E. Contour and persistence length of Corynebacterium diphtheriae pili by atomic force microscopy. Eur. Biophys. J. 2012, 41, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Formosa, C.; Herold, M.; Vidaillac, C.; Duval, R.E.; Dague, E. Unravelling of a mechanism of resistance to colistin in Klebsiella pneumoniae using atomic force microscopy. J. Antimicrob. Chemother. 2015, 70, 2261–2270. [Google Scholar] [CrossRef] [PubMed]
- Dufrêne, Y.F.; Ando, T.; Garcia, R.; Alsteens, D.; Martinez-Martin, D.; Engel, A.; Gerber, C.; Müller, D.J. Imaging modes of atomic force microscopy for application in molecular and cell biology. Nat. Nanotechnol. 2017, 12, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Radmacher, M.; Cleveland, J.P.; Fritz, M.; Hansma, H.G.; Hansma, P.K. Mapping interaction forces with the atomic force microscope. Biophys. J. 1994, 66, 2159–2165. [Google Scholar] [CrossRef]
- Senapati, S.; Lindsay, S. Recent Progress in Molecular Recognition Imaging Using Atomic Force Microscopy. Available online: https://pubs.acs.org/doi/full/10.1021/acs.accounts.5b00533 (accessed on 22 March 2018).
- Chopinet, L.; Formosa, C.; Rols, M.P.; Duval, R.E.; Dague, E. Imaging living cells surface and quantifying its properties at high resolution using AFM in QITM mode. Micron 2013, 48, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Smolyakov, G.; Formosa-Dague, C.; Severac, C.; Duval, R.E.; Dague, E. High speed indentation measures by FV, QI and QNM introduce a new understanding of bionanomechanical experiments. Micron 2016, 85, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ehrhardt, C.J.; Yadavalli, V.K. Single cell profiling of surface carbohydrates on Bacillus cereus. J. R. Soc. Interface 2015, 12, 20141109. [Google Scholar] [CrossRef] [PubMed]
- Formosa-Dague, C.; Speziale, P.; Foster, T.J.; Geoghegan, J.A.; Dufrêne, Y.F. Zinc-dependent mechanical properties of Staphylococcus aureus biofilm-forming surface protein SasG. Proc. Natl. Acad. Sci. USA 2016, 113, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Dague, E.; Alsteens, D.; Latgé, J.-P.; Verbelen, C.; Raze, D.; Baulard, A.R.; Dufrêne, Y.F. Chemical force microscopy of single live cells. Nano Lett. 2007, 7, 3026–3030. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, I.; Iyer, S.; Woodworth, C.D. Recovery of elasticity of aged human epithelial cells in vitro. Nanomed. Nanotechnol. Biol. Med. 2006, 2, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Ong, Q.K.; Sokolov, I. Attachment of nanoparticles to the AFM tips for direct measurements of interaction between a single nanoparticle and surfaces. J. Coll. Interface Sci. 2007, 310, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Hinterdorfer, P.; Baumgartner, W.; Gruber, H.J.; Schilcher, K.; Schindler, H. Detection and localization of individual antibody-antigen recognition events by atomic force microscopy. Proc. Natl. Acad. Sci. USA 1996, 93, 3477–3481. [Google Scholar] [CrossRef] [PubMed]
- Beaussart, A.; El-Kirat-Chatel, S.; Sullan, R.M.A.; Alsteens, D.; Herman, P.; Derclaye, S.; Dufrêne, Y.F. Quantifying the forces guiding microbial cell adhesion using single-cell force spectroscopy. Nat. Protocols 2014, 9, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yadavalli, V.K. Investigating biomolecular recognition at the cell surface using atomic force microscopy. Micron 2014, 60, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Valotteau, C.; Prystopiuk, V.; Pietrocola, G.; Rindi, S.; Peterle, D.; De Filippis, V.; Foster, T.J.; Speziale, P.; Dufrêne, Y.F. Single-Cell and Single-Molecule Analysis Unravels the Multifunctionality of the Staphylococcus aureus Collagen-Binding Protein Cna. ACS Nano 2017, 11, 2160–2170. [Google Scholar] [CrossRef] [PubMed]
- Ebner, A.; Wildling, L.; Zhu, R.; Rankl, C.; Haselgrübler, T.; Hinterdorfer, P.; Gruber, H.J. Functionalization of probe tips and supports for single-molecule recognition force microscopy. Top. Curr. Chem. 2008, 285, 29–76. [Google Scholar] [CrossRef] [PubMed]
- Wildling, L.; Unterauer, B.; Zhu, R.; Rupprecht, A.; Haselgrübler, T.; Rankl, C.; Ebner, A.; Vater, D.; Pollheimer, P.; Pohl, E.E.; et al. Linking of Sensor Molecules with Amino Groups to Amino-Functionalized AFM Tips. Bioconjugate Chem. 2011, 22, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Jauvert, E.; Dague, E.; Séverac, M.; Ressier, L.; Caminade, A.-M.; Majoral, J.-P.; Trévisiol, E. Probing single molecule interactions by AFM using bio-functionalized dendritips. Sens. Actuators B Chem. 2012, 168, 436–441. [Google Scholar] [CrossRef]
- Helenius, J.; Heisenberg, C.-P.; Gaub, H.E.; Muller, D.J. Single-cell force spectroscopy. J. Cell. Sci. 2008, 121, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Razatos, A.; Ong, Y.L.; Sharma, M.M.; Georgiou, G. Molecular determinants of bacterial adhesion monitored by atomic force microscopy. Proc. Natl. Acad. Sci. USA 1998, 95, 11059–11064. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.L.; Guérardel, Y.; Loubière, P.; Mercier-Bonin, M.; Dague, E. Measuring Kinetic Dissociation/Association Constants between Lactococcus lactis Bacteria and Mucins Using Living Cell Probes. Biophys. J. 2011, 101, 2843–2853. [Google Scholar] [CrossRef] [PubMed]
- Lower, S.K.; Hochella, M.F.; Beveridge, T.J. Bacterial Recognition of Mineral Surfaces: Nanoscale Interactions between Shewanella and α-FeOOH. Science 2001, 292, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Emerson, R.J.; Bergstrom, T.S.; Liu, Y.; Soto, E.R.; Brown, C.A.; McGimpsey, W.G.; Camesano, T.A. Microscale Correlation between Surface Chemistry, Texture, and the Adhesive Strength of Staphylococcus epidermidis. Langmuir 2006, 22, 11311–11321. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Elimelech, M. Bioinspired single bacterial cell force spectroscopy. Langmuir 2009, 25, 9656–9659. [Google Scholar] [CrossRef] [PubMed]
- Meister, A.; Gabi, M.; Behr, P.; Studer, P.; Vörös, J.; Niedermann, P.; Bitterli, J.; Polesel-Maris, J.; Liley, M.; Heinzelmann, H.; et al. FluidFM: combining atomic force microscopy and nanofluidics in a universal liquid delivery system for single cell applications and beyond. Nano Lett. 2009, 9, 2501–2507. [Google Scholar] [CrossRef] [PubMed]
- Potthoff, E.; Ossola, D.; Zambelli, T.; Vorholt, J.A. Bacterial adhesion force quantification by fluidic force microscopy. Nanoscale 2015, 7, 4070–4079. [Google Scholar] [CrossRef] [PubMed]
- Cone, R.A. Barrier properties of mucus. Adv. Drug Deliv. Rev. 2009, 61, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, K.S.; Kissoon-Singh, V.; Gibson, D.L.; Ma, C.; Montero, M.; Sham, H.P.; Ryz, N.; Huang, T.; Velcich, A.; Finlay, B.B.; et al. Muc2 protects against lethal infectious colitis by disassociating pathogenic and commensal bacteria from the colonic mucosa. PLoS Pathog. 2010, 6, e1000902. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.-S.; Lee, B.S.; Ryu, H.Y.; Sung, J.H.; Chung, K.H.; Yu, I.J. Effects of repeated silver nanoparticles exposure on the histological structure and mucins of nasal respiratory mucosa in rats. Toxicol. Lett. 2008, 182, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Schroeder, H.A.; Nunn, K.L.; Woods, K.; Anderson, D.J.; Lai, S.K.; Cone, R.A. Diffusion of Immunoglobulin G in Shed Vaginal Epithelial Cells and in Cell-Free Regions of Human Cervicovaginal Mucus. PLoS ONE 2016, 11, e0158338. [Google Scholar] [CrossRef] [PubMed]
- Mastrodonato, M.; Mentino, D.; Lopedota, A.; Cutrignelli, A.; Scillitani, G. A histochemical approach to glycan diversity in the urothelium of pig urinary bladder. Microsc. Res. Tech. 2017, 80, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.V.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Bansil, R.; Turner, B.S. Mucin structure, aggregation, physiological functions and biomedical applications. Curr. Opin. Colloid Interface Sci. 2006, 11, 164–170. [Google Scholar] [CrossRef]
- Allen, A.; Cunliffe, W.J.; Pearson, J.P.; Sellers, L.A.; Ward, R. Studies on gastrointestinal mucus. Scand. J. Gastroenterol. 1984, 93, 101–113. [Google Scholar]
- Zhou, D.; Li, J.; Li, N.; Yan, G. Study on viscosity property of gastrointestinal mucus. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi 2004, 21, 72–73. (In Chinese) [Google Scholar] [PubMed]
- Celli, J.P.; Turner, B.S.; Afdhal, N.H.; Ewoldt, R.H.; MaKindley, G.H.; Bansil, R.S. Erramilli Rheology of gastric mucin exhibits a pH-dependent Sol-gel transition. Biomacromolecules 2007, 8, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Kirch, J.; Schneider, A.; Abou, B.; Hopf, A.; Schaefer, U.F.; Schneider, M.; Schall, C.; Wagner, C.; Lehr, C.M. Optical tweezers reveal relationship between microstructure and nanoparticle penetration of pulmonary mucus. Proc. Natl. Acad. Sci. USA 2012, 109, 18355–18360. [Google Scholar] [CrossRef] [PubMed]
- Crater, J.S.; Carrier, R.L. Barrier properties of gastrointestinal mucus to nanoparticle transport. Macromol. Biosci. 2010, 10, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Lieleg, O.; Baumgartel, R.M.; Bausch, A.R. Selective Filtering of Particles by the Extracellular Matrix: An Electrostatic Bandpass. Biophys. J. 2009, 97, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, S.; Corner, G.; Dhima, E.; Houston, M.; Guha, C.; Augenlicht, L.; Velcich, A. MUC2 mucin deficiency alters inflammatory and metabolic pathways in the mouse intestinal mucosa. Oncotarget 2017, 8, 71456–71470. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, A.; Rossez, Y.; Robbe-Masselot, C.; Maes, E.; Gomes, J.; Shevtsova, A.; Bugaytsova, J.; Boren, T.; Reis, C.A. Muc5ac gastric mucin glycosylation is shaped by FUT2 activity and functionally impacts Helicobacter pylori binding. Sci. Rep. 2016, 6, 25575. [Google Scholar] [CrossRef] [PubMed]
- Lindén, S.K.; Florin, T.H.J.; McGuckin, M.A. Mucin Dynamics in Intestinal Bacterial Infection. PLoS ONE 2008, 3, e3952. [Google Scholar] [CrossRef] [PubMed]
- Juge, N. Microbial adhesins to gastrointestinal mucus. Trends Microbiol. 2012, 20, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Celli, J.P.; Turner, B.S.; Afdhal, N.H.; Keates, S.; Ghiran, I.; Kelly, C.P.; Ewoldt, R.H.; McKinley, G.H.; So, P.; Erramilli, S. Helicobacter pylori moves through mucus by reducing mucin viscoelasticity. Proc. Natl. Acad. Sci. USA 2009, 106, 14321–14326. [Google Scholar] [CrossRef] [PubMed]
- Bansil, R.; Celli, J.P.; Hardcastle, J.M.; Turner, B.S. The Influence of Mucus Microstructure and Rheology in Helicobacter pylori Infection. Front. Immunol. 2013, 4, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hage, N.; Howard, T.; Phillips, C.; Brassington, C.; Overman, R.; Debreczeni, J.; Gellert, P.; Stolnik, S.; Winkler, G.S.; Falcone, F.H. Structural basis of Lewisb antigen binding by the Helicobacter pylori adhesin BabA. Sci. Adv. 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Van Loosdrecht, M.C.M.; Lyklema, J.; Norde, W.; Zehnder, A.J.B. Bacterial adhesion: A physicochemical approach. Microb. Ecol. 1989, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Van Oss, C.J. Interfacial Forces in Aqueous Media; Taylor & Francis: Abingdon, UK, 1994; ISBN 978-0-8247-9168-1. [Google Scholar]
- Hermansson, M. The DLVO theory in microbial adhesion. Colloids Surf. B 1999, 14, 105–119. [Google Scholar] [CrossRef]
- Castelain, M.; Pignon, F.; Piau, J.M.; Magnin, A. The initial single yeast cell adhesion on glass via optical trapping and Derjaguin-Landau-Verwey-Overbeek predictions. J. Chem. Phys. 2008, 128, 135101. [Google Scholar] [CrossRef] [PubMed]
- Sieben, C.; Kappel, C.; Zhu, R.; Wozniak, A.; Rankl, C.; Hinterdorfer, P.; Grubmüller, H.; Herrmann, A. Influenza virus binds its host cell using multiple dynamic interactions. Proc. Natl. Acad. Sci. USA 2012, 109, 13626–13631. [Google Scholar] [CrossRef] [PubMed]
- Pyrpassopoulos, S.; Shuman, H.; Ostap, E.M. Adhesion force and attachment lifetime of the KIF16B-PX domain interaction with lipid membranes. Mol. Biol. Cell 2017, 28, 3315–3322. [Google Scholar] [CrossRef] [PubMed]
- Stillmark, P.H. Dorpat: Schnakenburg, Germany, 1888. Available online: http://hdl.handle.net/10062/2332 (accessed on 1 May 2018).
- Ofek, I.; Beachey, E.H. Mannose Binding and Epithelial Cell Adherence of Escherichia coli. Infect. Immun. 1978, 22, 247–254. [Google Scholar] [PubMed]
- Sharon, N. Carbohydrates as future anti-adhesion drugs for infectious diseases. Biochim. Biophys. Acta 2006, 1760, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.N.; Sun, D.; Dale, J.B.; Beachey, E.H. Conservation of the D-mannose-adhesion protein among type 1 fimbriated members of the family Enterobacteriaceae. Nature 1988, 336, 682–684. [Google Scholar] [CrossRef] [PubMed]
- Day, C.J.; Paton, A.W.; Harvey, R.M.; Hartley-Tassell, L.E.; Seib, K.L.; Tiralongo, J.; Bovin, N.; Savino, S.; Masignani, V.; Paton, J.C.; et al. Lectin activity of the pneumococcal pilin proteins. Sci. Rep. 2017, 7, 17784. [Google Scholar] [CrossRef] [PubMed]
- Morello, E.; Mallet, A.; Konto-Ghiorghi, Y.; Chaze, T.; Mistou, M.-Y.; Oliva, G.; Oliveira, L.; Guilmi, A.-M.D.; Trieu-Cuot, P.; Dramsi, S. Evidence for the Sialylation of PilA, the PI-2a Pilus-Associated Adhesin of Streptococcus agalactiae Strain NEM316. PLoS ONE 2015, 10, e0138103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercier-Bonin, M.; Chapot-Chartier, M.-P. Surface Proteins of Lactococcus lactis: Bacterial Resources for Muco-adhesion in the Gastrointestinal Tract. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Boekhorst, J.; Helmer, Q.; Kleerebezem, M.; Siezen, R.J. Comparative analysis of proteins with a mucus-binding domain found exclusively in lactic acid bacteria. Microbiology 2006, 152, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, D.T.; Tran, T.L.; Duviau, M.P.; Meyrand, M.; Guerardel, Y.; Castelain, M.; Loubiere, P.; Chapot-Chartier, M.P.; Dague, E.; Mercier-Bonin, M. Unraveling the role of surface mucus-binding protein and pili in muco-adhesion of Lactococcus lactis. PLoS ONE 2013, 8, e79850. [Google Scholar] [CrossRef] [PubMed]
- Dague, E.; Le, D.T.; Zanna, S.; Marcus, P.; Loubiere, P.; Mercier-Bonin, M. Probing in vitro interactions between Lactococcus lactis and mucins using AFM. Langmuir 2010, 26, 11010–11017. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Zanna, S.; Frateur, I.; Marcus, P.; Loubiere, P.; Dague, E.; Mercier-Bonin, M. Real-time investigation of the muco-adhesive properties of Lactococcus lactis using a quartz crystal microbalance with dissipation monitoring. Biofouling 2012, 28, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Beaussart, A.; Alsteens, D.; Dupres, V.; Claes, I.; von Ossowski, I.; de Vos, W.M.; Palva, A.; Lebeer, S.; Vanderleyden, J.; et al. Adhesion and nanomechanics of pili from the probiotic Lactobacillus rhamnosus GG. ACS Nano 2013, 7, 3685–3697. [Google Scholar] [CrossRef] [PubMed]
- Hilleringmann, M.; Giusti, F.; Baudner, B.C.; Masignani, V.; Covacci, A.; Rappuoli, R.; Barocchi, M.A.; Ferlenghi, I. Pneumococcal Pili Are Composed of Protofilaments Exposing Adhesive Clusters of Rrg A. PLoS Pathog. 2008, 4, e1000026. [Google Scholar] [CrossRef] [PubMed]
- Hilleringmann, M.; Ringler, P.; Muller, S.A.; De Angelis, G.; Rappuoli, R.; Ferlenghi, I.; Engel, A. Molecular architecture of Streptococcus pneumoniae TIGR4 pili. EMBO J. 2009, 28, 3921–3930. [Google Scholar] [CrossRef] [PubMed]
- Castelain, M.; Koutris, E.; Andersson, M.; Wiklund, K.; Bjornham, O.; Schedin, S.; Axner, O. Characterization of the Biomechanical Properties of T4 Pili Expressed by Streptococcus pneumoniae—A Comparison between Helix-like and Open Coil-like Pili. Chemphyschem 2009, 10, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Castelain, M.; Ehlers, S.; Klinth, J.; Lindberg, S.; Andersson, M.; Uhlin, B.E.; Axner, O. Fast uncoiling kinetics of F1C pili expressed by uropathogenic Escherichia coli are revealed on a single pilus level using force-measuring optical tweezers. Eur. Biophys. J. 2011, 40, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Zakrisson, J.; Wiklund, K.; Axner, O.; Andersson, M. The Shaft of the Type 1 Fimbriae Regulates an External Force to Match the FimH Catch Bond. Biophys. J. 2013, 104, 2137–2148. [Google Scholar] [CrossRef] [PubMed]
- Björnham, O.; Nilsson, H.; Andersson, M.; Schedin, S. Physical properties of the specific PapG-galabiose binding in E. coli P pili-mediated adhesion. Eur. Biophys. J. 2009, 38, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Zakrisson, J.; Wiklund, K.; Axner, O.; Andersson, M. Helix-like biopolymers can act as dampers of force for bacteria in flows. Eur. Biophys. J. 2012, 41, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Björnham, O.; Axner, O. Multipili attachment of bacteria with helixlike pili exposed to stress. J. Chem. Phys. 2009, 130, 235102. [Google Scholar] [CrossRef] [PubMed]
- Epler Barbercheck, C.R.; Bullitt, E.; Andersson, M. Bacterial Adhesion Pili. In Membrane Protein Complexes: Structure and Function; Harris, J.R., Boekema, E.J., Eds.; Springer: Singapore, 2018; Volume 87, pp. 1–18. ISBN 978-981-10-7756-2. [Google Scholar]
- Thomas, W.E.; Nilsson, L.M.; Forero, M.; Sokurenko, E.V.; Vogel, V. Shear-dependent “stick-and-roll” adhesion of type 1 fimbriated Escherichia coli. Mol. Microbiol. 2004, 53, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.; Garcia, T.; Hultgren, S.; Oberhauser, A.F. The mechanical properties of E. coli type 1 pili measured by atomic force microscopy techniques. Biophys. J. 2006, 91, 3848–3856. [Google Scholar] [CrossRef] [PubMed]
- Bullitt, E.; Gong, M.; Makowski, L. A Molecular Bungee Cord—3-Dimensional Helical Reconstruction of Bacterial Adhesion Pili by Electron-Microscopy. Mol. Biol. Cell 1992, 3, A171. [Google Scholar]
- Bullitt, E.; Makowski, L. Bacterial adhesion pili are heterologous assemblies of similar subunits. Biophys. J. 1998, 74, 623–632. [Google Scholar] [CrossRef]
- Liang, M.N.; Smith, S.P.; Metallo, S.J.; Choi, I.S.; Prentiss, M.; Whitesides, G.M. Measuring the forces involved in polyvalent adhesion of uropathogenic Escherichia coli to mannose-presenting surfaces. Proc. Natl. Acad. Sci. USA 2000, 97, 13092–13096. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Uhlin, B.E.; Fällman, E. The biomechanical properties of E. coli pili for urinary tract attachment reflect the host environment. Biophys. J. 2007, 93, 3008–3014. [Google Scholar] [CrossRef] [PubMed]
- Castelain, M.; Sjostrom, A.E.; Fallman, E.; Uhlin, B.E.; Andersson, M. Unfolding and refolding properties of S pili on extraintestinal pathogenic Escherichia coli. Eur. Biophys. J. 2010, 39, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.E.; Trintchina, E.; Forero, M.; Vogel, V.; Sokurenko, E.V. Bacterial Adhesion to Target Cells Enhanced by Shear Force. Cell 2002, 109, 913–923. [Google Scholar] [CrossRef]
- Zhou, G.; Mo, W.-J.; Sebbel, P.; Min, G.; Neubert, T.A.; Glockshuber, R.; Wu, X.-R.; Sun, T.-T.; Kong, X.-P. Uroplakin Ia is the urothelial receptor for uropathogenic Escherichia coli: Evidence from in vitro FimH binding. J. Cell Sci. 2001, 114, 4095–4103. [Google Scholar] [PubMed]
- Ofek, I.; Hasty, D.L.; Sharon, N. Anti-adhesion therapy of bacterial diseases: Prospects and problems. FEMS Immunol. Med. Microbiol. 2003, 38, 181–191. [Google Scholar] [CrossRef]
- Dansuk, K.C.; Keten, S. Tunable seat belt behavior in nanocomposite interfaces inspired from bacterial adhesion pili. Soft Matter 2018, 14, 1530–1539. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.M.; Yakovenko, O.; Tchesnokova, V.; Thomas, W.E.; Schembri, M.A.; Vogel, V.; Klemm, P.; Sokurenko, E.V. The cysteine bond in the Escherichia coli FimH adhesin is critical for adhesion under flow conditions. Mol. Microbiol. 2007, 65, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- Sauer, M.M.; Jakob, R.P.; Eras, J.; Baday, S.; Eriş, D.; Navarra, G.; Bernèche, S.; Ernst, B.; Maier, T.; Glockshuber, R. Catch-bond mechanism of the bacterial adhesin FimH. Nat. Commun. 2016, 7, 10738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquot, A.; Sakamoto, C.; Razafitianamaharavo, A.; Caillet, C.; Merlin, J.; Fahs, A.; Ghigo, G.; Beloin, C.; Duval, J.F.L.; Francius, G. Dynamic modulation of fimbrial extension and FimH-mannose binding force on live bacteria under pH changes: A molecular atomic force microscopy analysis. J. Biomed. Nanotechnol. 2014, 10, 3361–3372. [Google Scholar] [CrossRef] [PubMed]
- Klinth, J.E.; Castelain, M.; Uhlin, B.E.; Axner, O. The Influence of pH on the Specific Adhesion of P Piliated Escherichia coli. PLoS ONE 2012, 7, e38548. [Google Scholar] [CrossRef] [PubMed]
- Merz, A.J.; So, M.; Sheetz, M.P. Pilus retraction powers bacterial twitching motility. Nature 2000, 407, 98–102. [Google Scholar] [PubMed]
- Maier, B. Using laser tweezers to measure twitching motility in Neisseria. Curr. Opin. Microbiol. 2005, 8, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Maier, B.; Potter, L.; So, M.; Seifert, H.S.; Sheetz, M.P. Single pilus motor forces exceed 100 pN. Proc. Natl. Acad. Sci. USA 2002, 99, 16012–16017. [Google Scholar] [CrossRef] [PubMed]
- Biais, N.; Higashi, D.L.; Brujić, J.; So, M.; Sheetz, M.P. Force-dependent polymorphism in type IV pili reveals hidden epitopes. Proc. Natl. Acad. Sci. USA 2010, 107, 11358–11363. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.L.; Biais, N.; Tama, F. Steered Molecular Dynamics Simulations of a Type IV Pilus Probe Initial Stages of a Force-Induced Conformational Transition. PLoS Comput. Biol. 2013, 9, e1003032. [Google Scholar] [CrossRef] [PubMed]
- Maier, B.; Wong, G.C.L. How Bacteria Use Type IV Pili Machinery on Surfaces. Trends Microbiol. 2015, 23, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Gunning, A.P.; Kavanaugh, D.; Thursby, E.; Etzold, S.; MacKenzie, D.A.; Juge, N. Use of Atomic Force Microscopy to Study the Multi-Modular Interaction of Bacterial Adhesins to Mucins. Int. J. Mol. Sci. 2016, 17, 1854. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Gabius, H.-J.; André, S.; Kaltner, H.; Sabesan, S.; Roy, R.; Liu, B.; Macaluso, F.; Brewer, C.F. Galectin-3 Precipitates as a Pentamer with Synthetic Multivalent Carbohydrates and Forms Heterogeneous Cross-linked Complexes. J. Biol. Chem. 2004, 279, 10841–10847. [Google Scholar] [CrossRef] [PubMed]
- Sletmoen, M.; Dam, T.K.; Gerken, T.A.; Stokke, B.T.; Brewer, C.F. Single-Molecule Pair Studies of the Interactions of the alpha-GalNAc (Tn-Antigen) Form of Porcine Submaxillary Mucin with Soybean Agglutinin. Biopolymers 2009, 91, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Dam, T.K.; Brewer, C.F. Probing Lectin–Mucin Interactions by Isothermal Titration Microcalorimetry. In Galectins; Stowell, S.R., Cummings, R.D., Eds.; Springer New York: New York, NY, USA, 2015; Volume 1207, pp. 75–90. ISBN 978-1-4939-1395-4. [Google Scholar]
- Hadjialirezaei, S.; Picco, G.; Beatson, R.; Burchell, J.; Stokke, B.T.; Sletmoen, M. Interactions between the breast cancer-associated MUC1 mucins and C-type lectin characterized by optical tweezers (vol. 12, e0175323, 2017). PLoS ONE 2018, 13, e0190814. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kotsuchibashi, Y.; Liu, Y.; Narain, R. Study of Bacterial Adhesion on Biomimetic Temperature Responsive Glycopolymer Surfaces. Available online: http://pubs.acs.org/doi/abs/10.1021/am508792k (accessed on 15 March 2018).
- Varki, A.; Kornfeld, S. Historical Background and Overview. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015. [Google Scholar]
- Mahdavi, J.; Sondén, B.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Angstrom, J.; Larsson, T.; Teneberg, S.; Karlsson, K.-A.; et al. Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Day, C.J.; Tran, E.N.; Semchenko, E.A.; Tram, G.; Hartley-Tassell, L.E.; Ng, P.S.K.; King, R.M.; Ulanovsky, R.; McAtamney, S.; Apicella, M.A.; et al. Glycan:glycan interactions: High affinity biomolecular interactions that can mediate binding of pathogenic bacteria to host cells. Proc. Natl. Acad. Sci. USA 2015, 112, E7266–E7275. [Google Scholar] [CrossRef] [PubMed]
- Mubaiwa, T.D.; Hartley-Tassell, L.E.; Semchenko, E.A.; Jen, F.E.-C.; Srikhanta, Y.N.; Day, C.J.; Jennings, M.P.; Seib, K.L. The glycointeractome of serogroup B Neisseria meningitidis strain MC58. Sci. Rep. 2017, 7, 5693. [Google Scholar] [CrossRef] [PubMed]
- Mubaiwa, T.D.; Semchenko, E.A.; Hartley-Tassell, L.E.; Day, C.J.; Jennings, M.P.; Seib, K.L. The sweet side of the pathogenic Neisseria: The role of glycan interactions in colonisation and disease. Pathog. Dis. 2017, 75. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, N.; Kilcoyne, M.; Joshi, L.; Hickey, R.M. Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria. Sensors 2017, 17, 1515. [Google Scholar] [CrossRef] [PubMed]
- Belotserkovsky, I.; Brunner, K.; Pinaud, L.; Rouvinski, A.; Dellarole, M.; Baron, B.; Dubey, G.; Samassa, F.; Parsot, C.; Sansonetti, P.; et al. Glycan-Glycan Interaction Determines Shigella Tropism toward Human T Lymphocytes. mBio 2018, 9, e02309-17. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-H.; Hütter, J.; Hsu, C.-W.; Tanaka, H.; Varela-Aramburu, S.; De Cola, L.; Lepenies, B.; Seeberger, P.H. Analysis of Carbohydrate-Carbohydrate Interactions Using Sugar-Functionalized Silicon Nanoparticles for Cell Imaging. Nano Lett. 2016, 16, 807–811. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.M.; Barrientos, A.G.; Rojas, T.C.; Rojo, J.; Cañada, J.; Fernández, A.; Penadés, S. Gold Glyconanoparticles as Water-Soluble Polyvalent Models To Study Carbohydrate Interactions. Angew. Chem. Int. Ed. 2001, 40, 2257–2261. [Google Scholar] [CrossRef]
- Hernáiz, M.J.; de la Fuente, J.M.; Barrientos, Á.G.; Penadés, S. A Model System Mimicking Glycosphingolipid Clusters to Quantify Carbohydrate Self-Interactions by Surface Plasmon Resonance. Angew. Chem. 2002, 41, 1554–1557. [Google Scholar] [CrossRef]
- Santacroce, P.V.; Basu, A. Probing specificity in carbohydrate-carbohydrate interactions with micelles and Langmuir monolayers. Angew. Chem. Int. Ed. Engl. 2003, 42, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, Y.; Park, H.-J.; Boggs, J.M.; Basu, A. Carbohydrate-coated fluorescent silica nanoparticles as probes for the galactose/3-sulfogalactose carbohydrate-carbohydrate interaction using model systems and cellular binding studies. Bioconjugate Chem. 2012, 23, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Misevic, G.N. Molecular self-recognition and adhesion via proteoglycan to proteoglycan interactions as a pathway to multicellularity: Atomic force microscopy and color coded bead measurements in sponges. Microsc. Res. Tech. 1999, 44, 304–309. [Google Scholar] [CrossRef]
- Tromas, C.; Rojo, J.; de la Fuente, J.M.; Barrientos, A.G.; García, R.; Penadés, S. Adhesion Forces between Lewis(X) Determinant Antigens as Measured by Atomic Force Microscopy. Angew. Chem. Int. Ed. Engl. 2001, 40, 3052–3055. [Google Scholar] [CrossRef]
- Carvalho de Souza, A.; Ganchev, D.N.; Snel, M.M.E.; van der Eerden, J.P.J.M.; Vliegenthart, J.F.G.; Kamerling, J.P. Adhesion forces in the self-recognition of oligosaccharide epitopes of the proteoglycan aggregation factor of the marine sponge Microciona prolifera. Glycoconj. J. 2009, 26, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Haugstad, K.E.; Gerken, T.A.; Stokke, B.T.; Dam, T.K.; Brewer, C.F.; Sletmoen, M. Enhanced self-association of mucins possessing the T and Tn carbohydrate cancer antigens at the single-molecule level. Biomacromolecules 2012, 13, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Haugstad, K.E.; Hati, A.G.; Nordgard, C.T.; Adl, P.S.; Maurstad, G.; Sletmoen, M.; Draget, K.I.; Dias, R.S.; Stokke, B.T. Direct Determination of Chitosan-Mucin Interactions Using a Single-Molecule Strategy: Comparison to Alginate-Mucin Interactions. Polymers 2015, 7, 161–185. [Google Scholar] [CrossRef]
- Haugstad, K.E.; Hadjialirezaei, S.; Stokke, B.T.; Brewer, C.F.; Gerken, T.A.; Burchell, J.; Picco, G.; Sletmoen, M. Interactions of mucins with the Tn or Sialyl Tn cancer antigens including MUC1 are due to GalNAc-GalNAc interactions. Glycobiology 2016, 26, 1338–1350. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S. Carbohydrate-to-carbohydrate interaction, through glycosynapse, as a basis of cell recognition and membrane organization. Glycoconj. J. 2004, 21, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Zhao, W.; Shang, X.; Jiang, J.; Ji, H.; Tang, Z.; Wang, H. Direct evidence of lipid rafts by in situ atomic force microscopy. Small 2012, 8, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Gellermann, G.P.; Appel, T.R.; Tannert, A.; Radestock, A.; Hortschansky, P.; Schroeckh, V.; Leisner, C.; Lütkepohl, T.; Shtrasburg, S.; Röcken, C.; et al. Raft lipids as common components of human extracellular amyloid fibrils. Proc. Natl. Acad. Sci. USA 2005, 102, 6297–6302. [Google Scholar] [CrossRef] [PubMed]
- González-Maeso, J. GPCR oligomers in pharmacology and signaling. Mol. Brain 2011, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Adamcik, J.; Jung, J.-M.; Flakowski, J.; De Los Rios, P.; Dietler, G.; Mezzenga, R. Understanding amyloid aggregation by statistical analysis of atomic force microscopy images. Nat. Nano 2010, 5, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.M.; Koulov, A.V.; Balch, W.E.; Kelly, J.W. Functional amyloid—From bacteria to humans. Trends Biochem. Sci. 2007, 32, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Varki, A. Chapter Three—Modulation of Glycan Recognition by Clustered Saccharide Patches. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 308, pp. 75–125. [Google Scholar]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.S.; Chishti, A.H. Host Receptors in Malaria Merozoite Invasion. In Malaria: Drugs, Disease and Post-genomic Biology; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 203–232. ISBN 978-3-540-25363-1. [Google Scholar]
- Matsuo, K.; Ota, H.; Akamatsu, T.; Sugiyama, A.; Katsuyama, T. Histochemistry of the surface mucous gel layer of the human colon. Gut 1997, 40, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Baos, S.C.; Phillips, D.B.; Wildling, L.; McMaster, T.J.; Berry, M. Distribution of Sialic Acids on Mucins and Gels: A Defense Mechanism. Biophys. J. 2012, 102, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Bazaka, K.; Crawford, R.J.; Nazarenko, E.L.; Ivanova, E.P. Bacterial Extracellular Polysaccharides. In Bacterial Adhesion; Advances in Experimental Medicine and Biology; Springer: Dordrecht, The Netherlands, 2011; pp. 213–226. ISBN 978-94-007-0939-3. [Google Scholar]
- Te Riet, J.; Joosten, B.; Reinieren-Beeren, I.; Figdor, C.G.; Cambi, A. N-glycan mediated adhesion strengthening during pathogen-receptor binding revealed by cell-cell force spectroscopy. Sci. Rep. 2017, 7, 6713. [Google Scholar] [CrossRef] [PubMed]
- Schillers, H.; Rianna, C.; Schäpe, J.; Luque, T.; Doschke, H.; Wälte, M.; Uriarte, J.J.; Campillo, N.; Michanetzis, G.P.A.; Bobrowska, J.; et al. Standardized Nanomechanical Atomic Force Microscopy Procedure (SNAP) for Measuring Soft and Biological Samples. Sci. Rep. 2017, 7, 5117. [Google Scholar] [CrossRef] [PubMed]
- Beaussart, A.; Baker, A.E.; Kuchma, S.L.; El-Kirat-Chatel, S.; O’Toole, G.A.; Dufrêne, Y.F. Nanoscale Adhesion Forces of Pseudomonas aeruginosa Type IV Pili. ACS Nano 2014, 8, 10723–10733. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, I.J.; Poretz, R.D. 2—Isolation, Physicochemical Characterization, and Carbohydrate-Binding Specificity of Lectins. In The Lectins; Academic Press: Cambridge, MA, USA, 1986; pp. 33–247. ISBN 978-0-12-449945-4. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
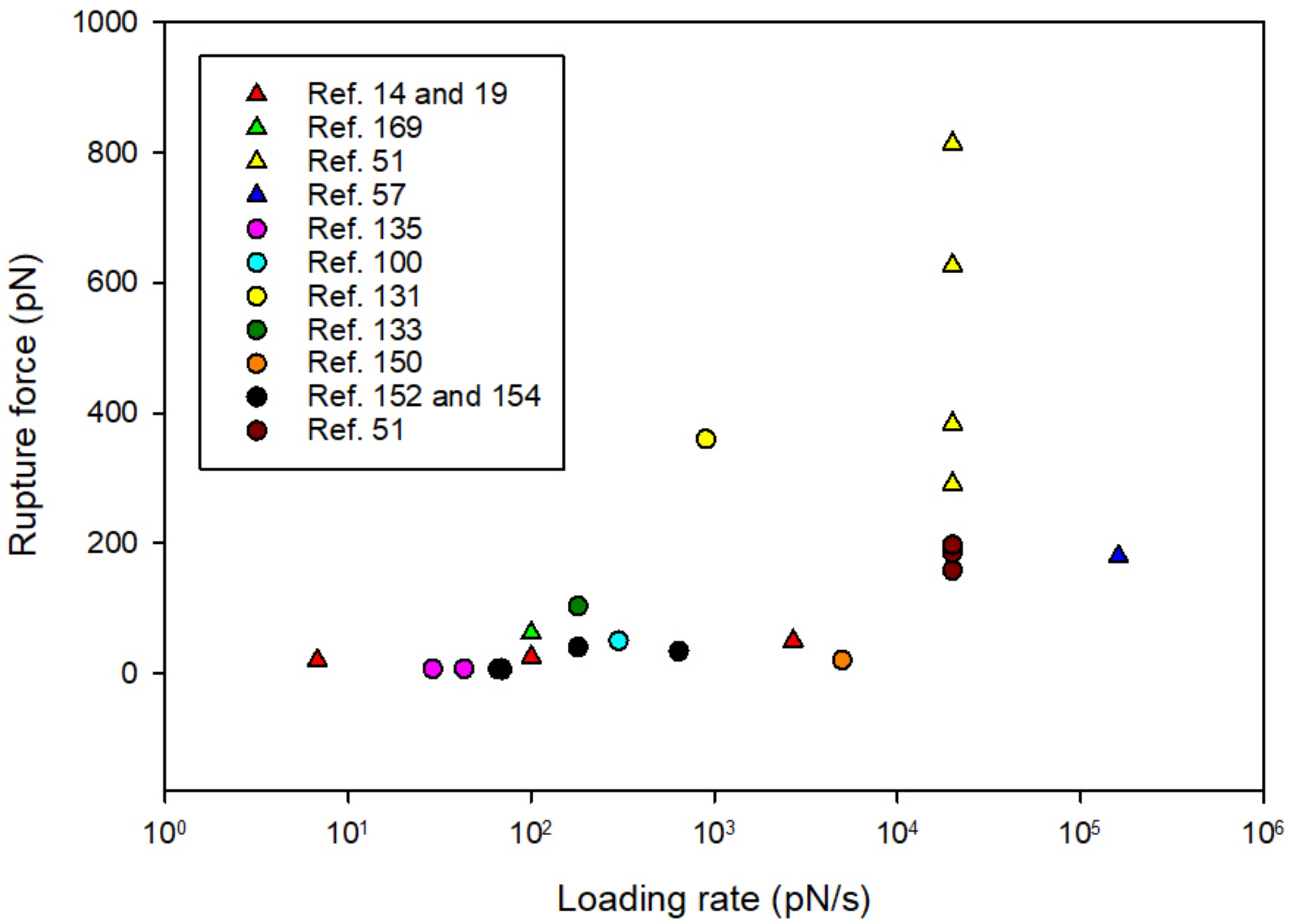
| Reference | System Studied | Force Probe | Rupture Force (pN) | Loading Rate (pN/s) | koff (1/s) | |
|---|---|---|---|---|---|---|
| Single bacterium-substrate interactions | Protein-glycan interactions | |||||
| Björnham [14] | H. pylori BabA—Leb | OT | 25 ± 1.5 | 100 | N/A | |
| Björnham [19] | H. pylori BabA—Leb | OT | 20 * 50 * | 6.8 2700 | 0.86 ± 0.07 | |
| Beaussart [169] | P. aeruginosa pili—Pneumocyte | AFM | 62 ± 11 | 100 | N/A | |
| Simpson [11] | S. aureus—fibrinogen S. aureus—fibronectin | OT | 20 25 | N/A N/A | N/A | |
| Le [57] | L. lactis—PGM | AFM | 180 ± 4 | 161,250 | N/A | |
| Protein-protein interactions | ||||||
| Valotteau et al. [51] | S. aureus—complement system protein (C1q) | AFM | 291 ± 97 (JPK instrument) 384 ± 48 (Bruker instrument) | ~20,000 # | N/A | |
| S. aureus—Laminin | AFM | 814 ± 179 (JPK instrument) 627 ± 122 (Bruker instrument) | ~20,000 # | N/A | ||
| Single molecule interactions | Protein-glycan interactions | |||||
| Hadjialirezaei [135] | MUC1(Tn)—MGL | OT | 6.8 ± 0.8 | 29 ± 2 | 2.0 | |
| MUC1(STn)—MGL | OT | 7.1 ± 1.1 | 43 ± 3 | 3.3 | ||
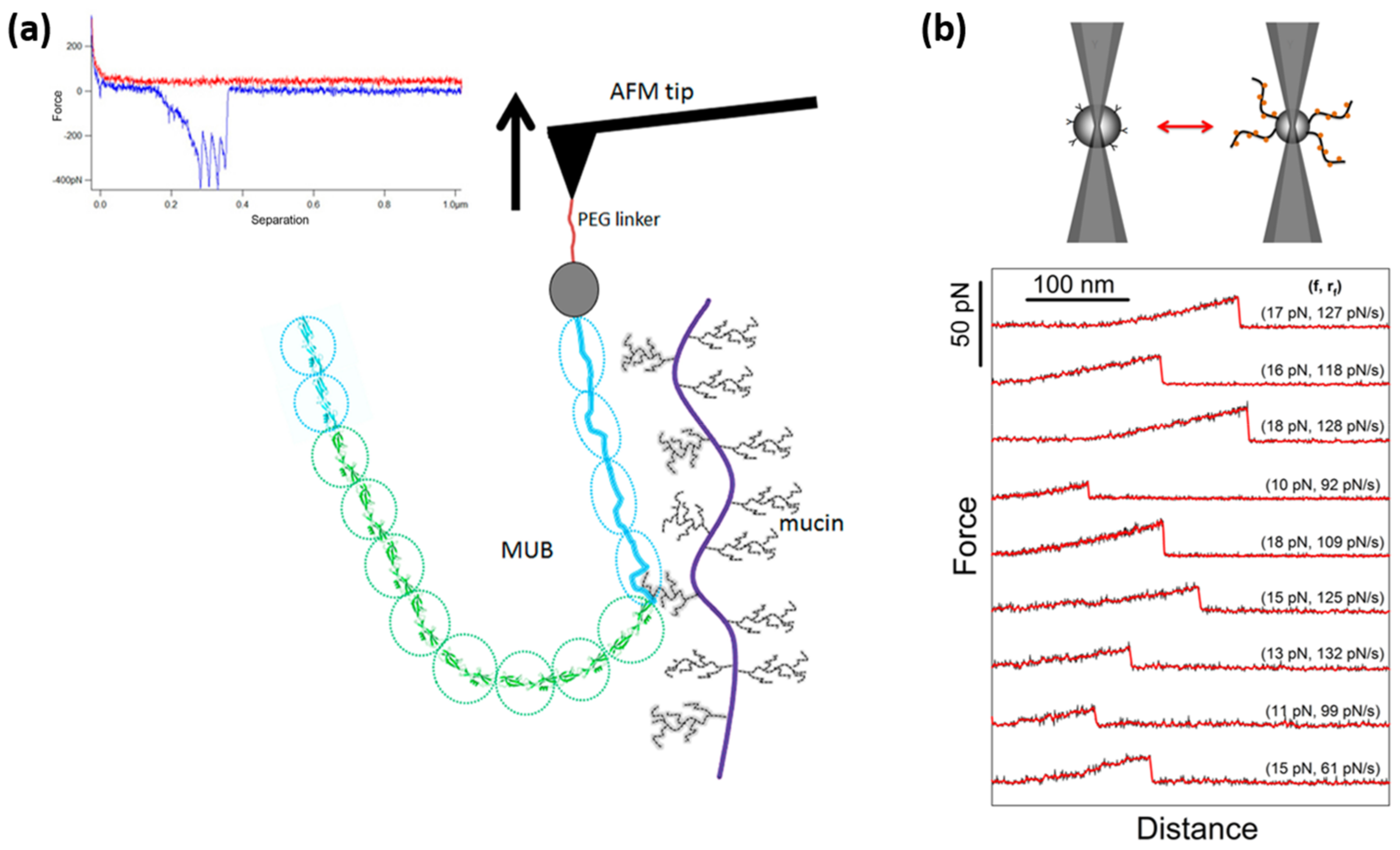
| Tripathi [100] | SpaC pili-protein—mucin | AFM | 50 | 300 * | 0.05 | |
| Gunning [131] | MUB—mucin | AFM | 380–342 | 900 * | N/A | |
| Sletmoen [133] | SBA—PGM | AFM | 103 | 180 | 0.76 ± 0.09 | |
| Glycan-glycan interactions | ||||||
| Tromas [150] | Lex—Lex self-interaction | AFM | 20 ± 4 | ~5000 | N/A | |
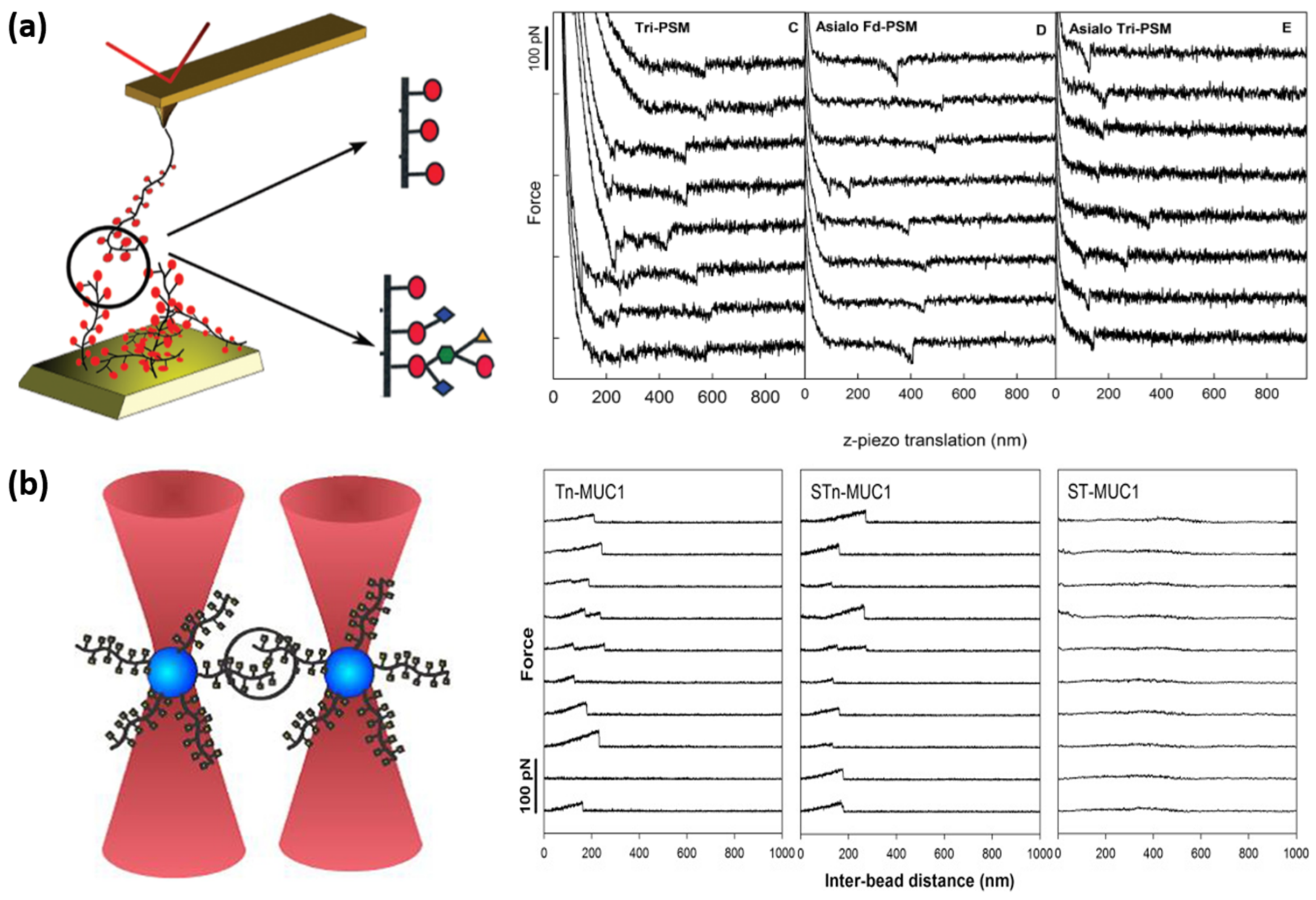
| Haugstad [152] | Tn-PSM—Tn-PSM | AFM | 34 | 640 | 1.2 ± 0.27 | |
| Fd-PSM—Fd-PSM | 40 | 180 | 0.64 ± 0.8 | |||
| Haugstad [154] | Tn-PSM—Tn-PSM | OT | 7.2 | 69 | 3.8 | |
| Tri-PSM—Tri-PSM | 5.6 | 69 | 5.0 | |||
| STn-OSM—STn-OSM | 6.2 | 66 | 6.2 | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Formosa-Dague, C.; Castelain, M.; Martin-Yken, H.; Dunker, K.; Dague, E.; Sletmoen, M. The Role of Glycans in Bacterial Adhesion to Mucosal Surfaces: How Can Single-Molecule Techniques Advance Our Understanding? Microorganisms 2018, 6, 39. https://doi.org/10.3390/microorganisms6020039
Formosa-Dague C, Castelain M, Martin-Yken H, Dunker K, Dague E, Sletmoen M. The Role of Glycans in Bacterial Adhesion to Mucosal Surfaces: How Can Single-Molecule Techniques Advance Our Understanding? Microorganisms. 2018; 6(2):39. https://doi.org/10.3390/microorganisms6020039
Chicago/Turabian StyleFormosa-Dague, Cécile, Mickaël Castelain, Hélène Martin-Yken, Karen Dunker, Etienne Dague, and Marit Sletmoen. 2018. "The Role of Glycans in Bacterial Adhesion to Mucosal Surfaces: How Can Single-Molecule Techniques Advance Our Understanding?" Microorganisms 6, no. 2: 39. https://doi.org/10.3390/microorganisms6020039
APA StyleFormosa-Dague, C., Castelain, M., Martin-Yken, H., Dunker, K., Dague, E., & Sletmoen, M. (2018). The Role of Glycans in Bacterial Adhesion to Mucosal Surfaces: How Can Single-Molecule Techniques Advance Our Understanding? Microorganisms, 6(2), 39. https://doi.org/10.3390/microorganisms6020039