Binding of Helicobacter pylori to Human Gastric Mucins Correlates with Binding of TFF1

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Harvest and Purification of Animal and Human Gastric and Colonic Mucins
2.3. Construction of Animal and Human Gastrointestinal Tract (GIT) Mucin Microarrays
2.4. Profiling of Mucin Microarrays with Lectins
2.5. Profiling of Mucin Microarrays with TFF1
2.6. Interrogation of Human Mucin Microarrays with H. pylori and with C. jejuni
2.7. Microarray Data Extraction and Analysis
2.8. Statistical and Hierarchical Clustering Analysis
3. Results
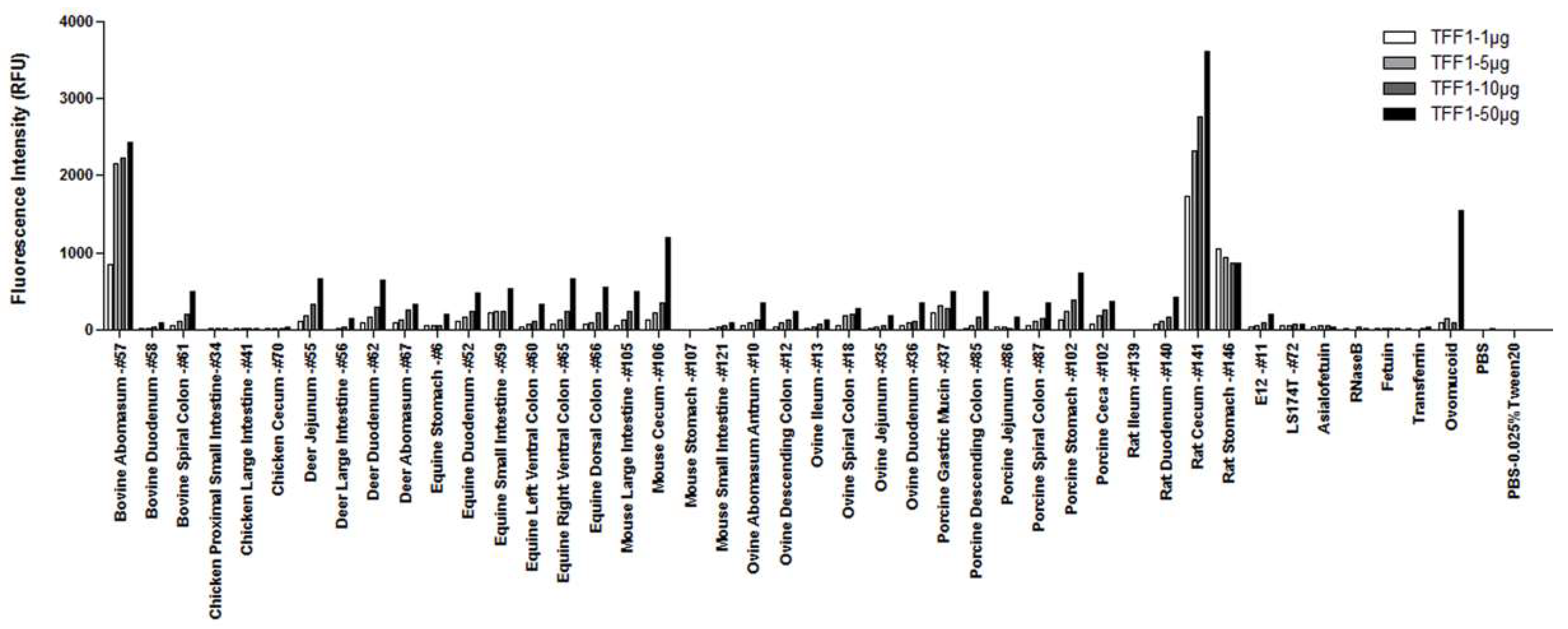
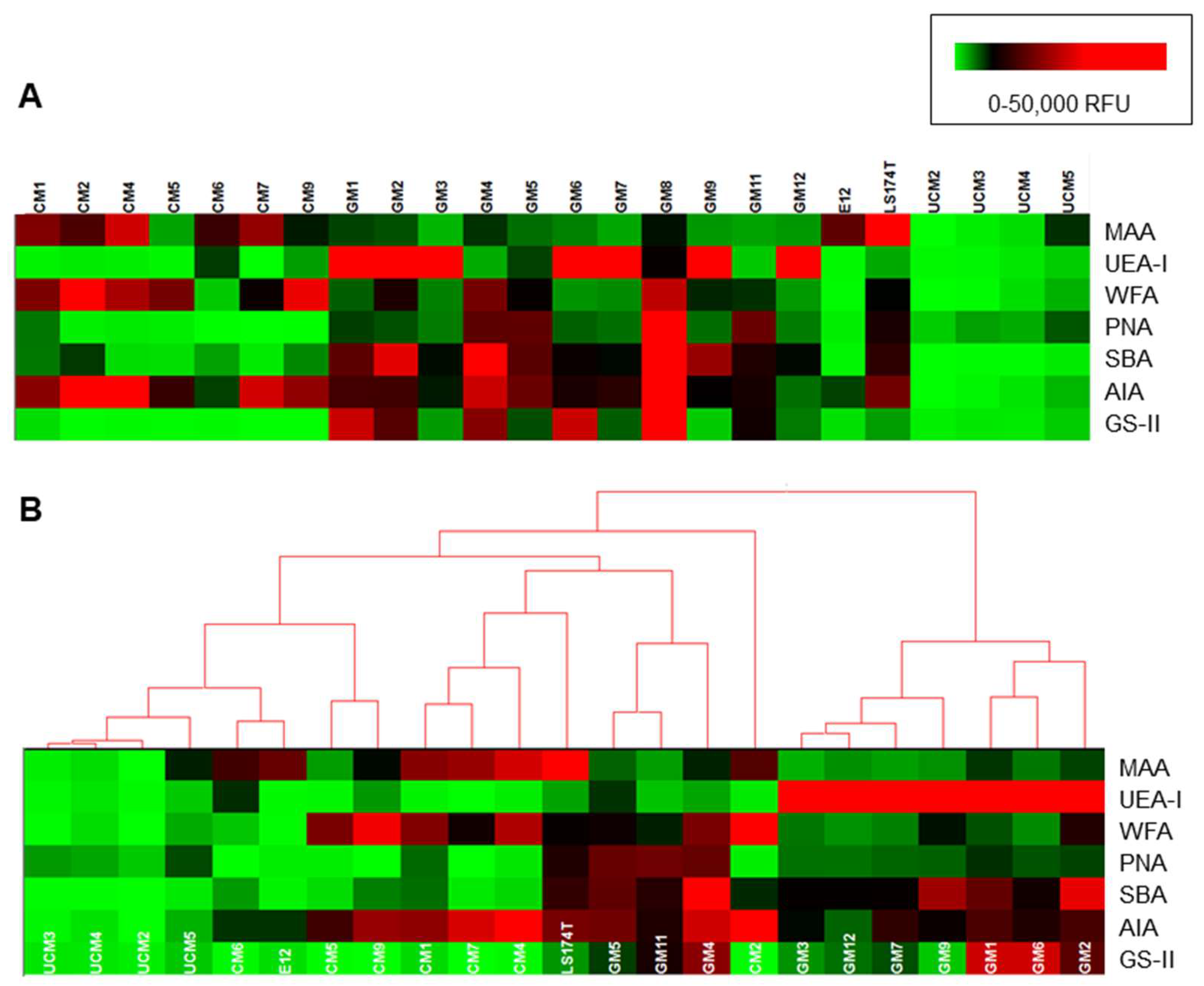
3.1. TFF1 Interacts Specifically with Human and Animal Gastric Mucins
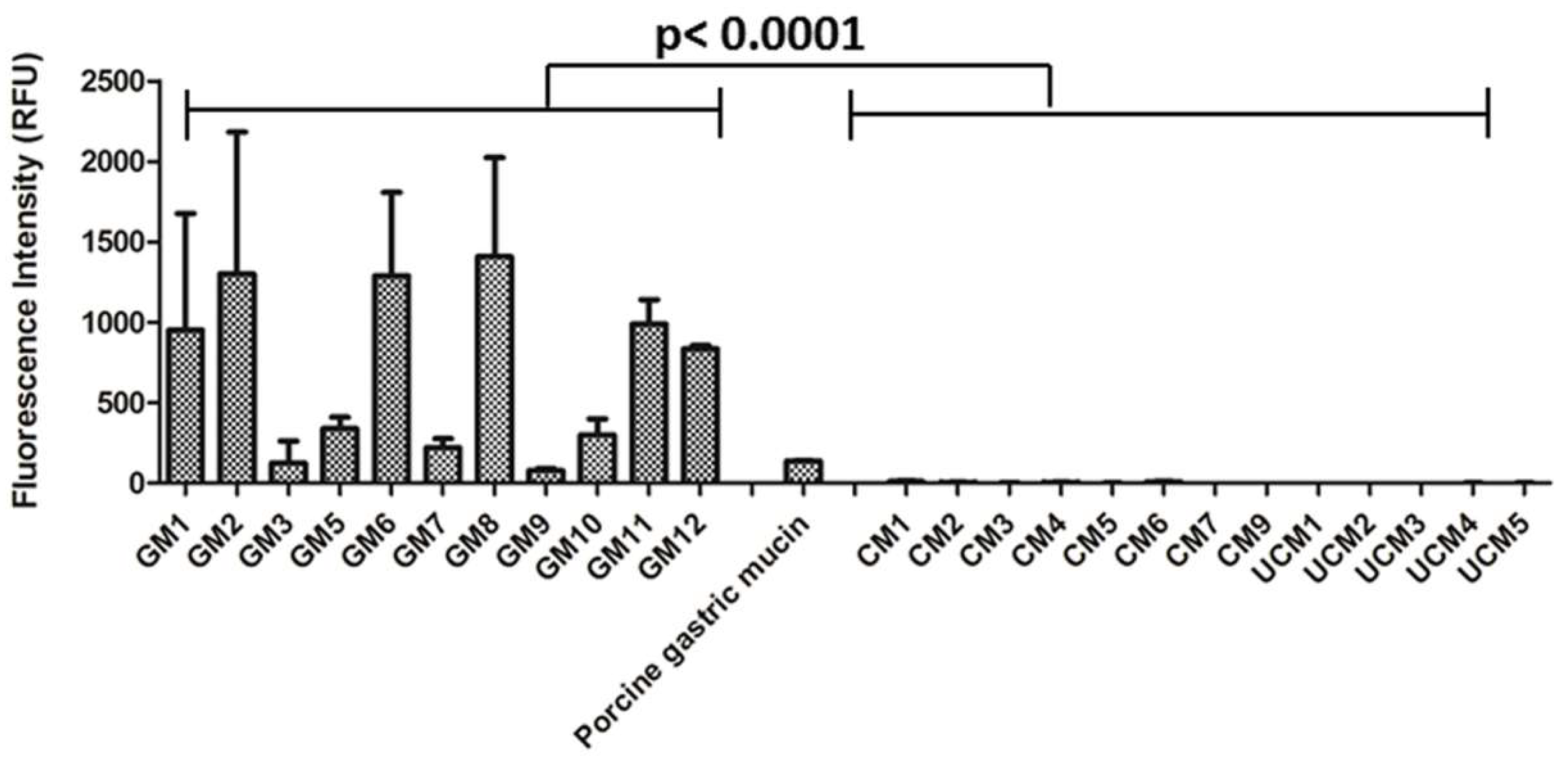
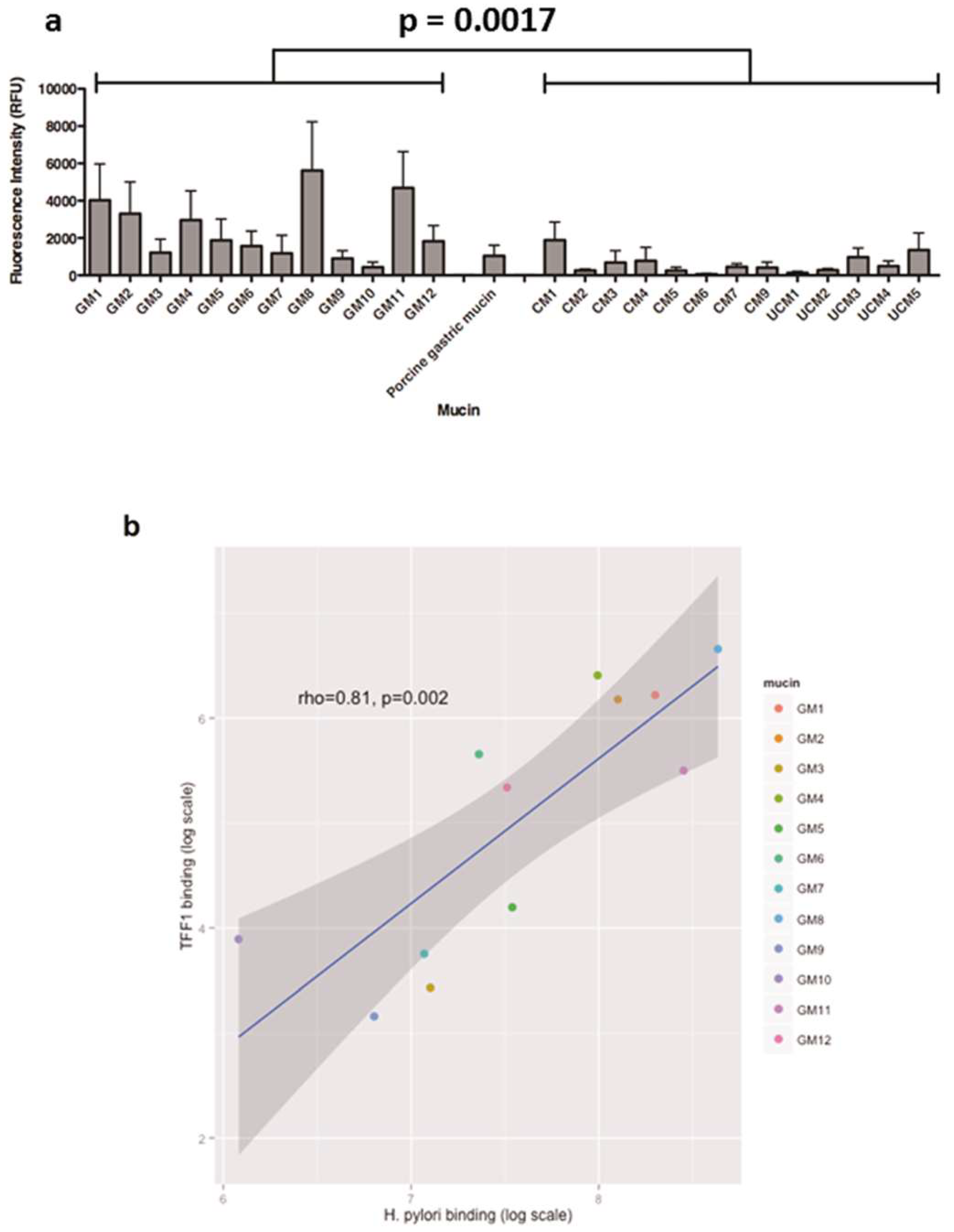
3.2. The Interaction of H. pylori with Human Gastric Mucins Correlates Closely with TFF1 Binding
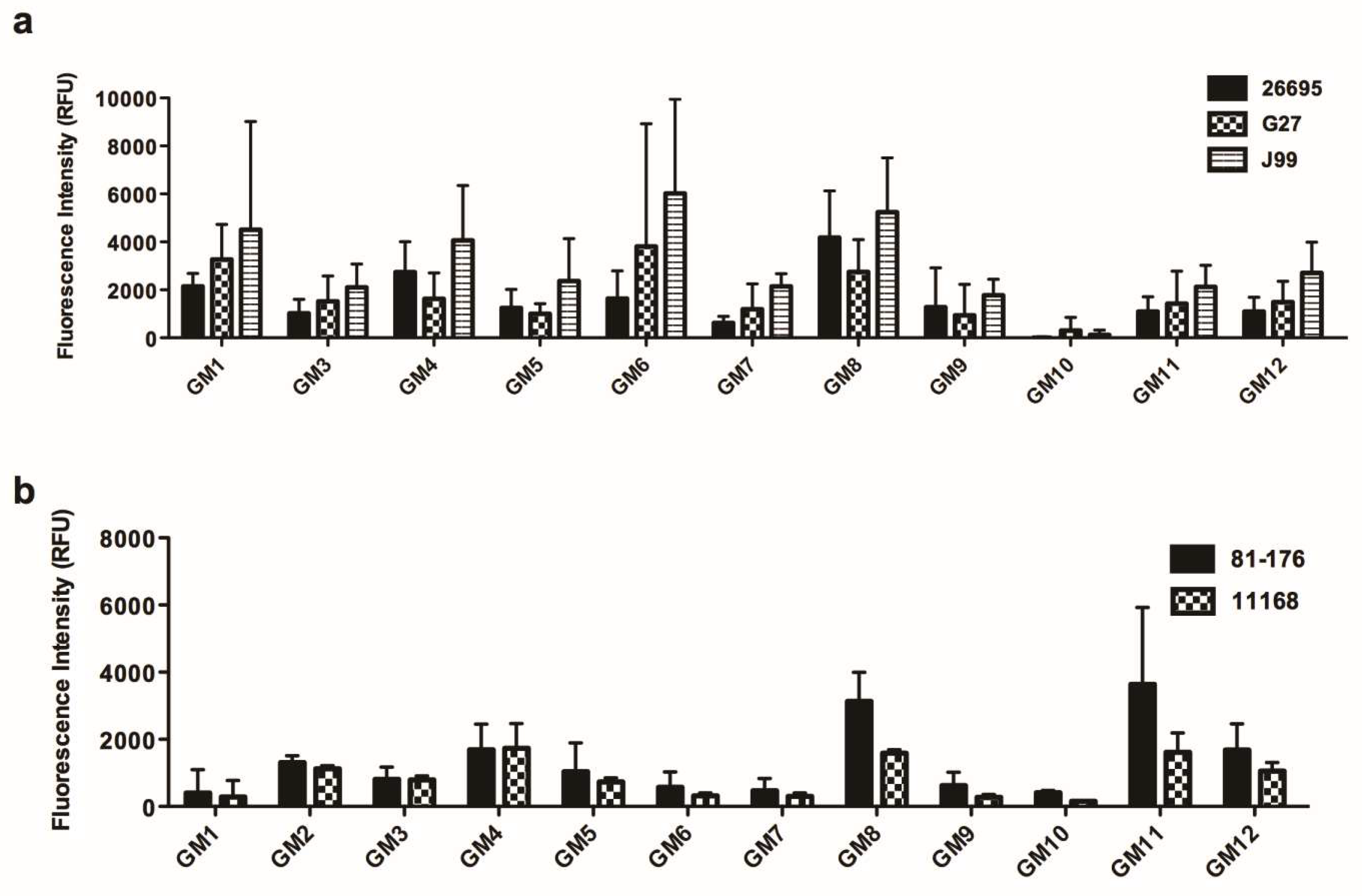
3.3. The Interaction of Various H. pylori and C. jejuni Strains to Human Gastric Mucins and TFF1
3.4. Binding of TFF1 and H. pylori to Gastric Mucins Correlates with Binding of the Lectin GS-II
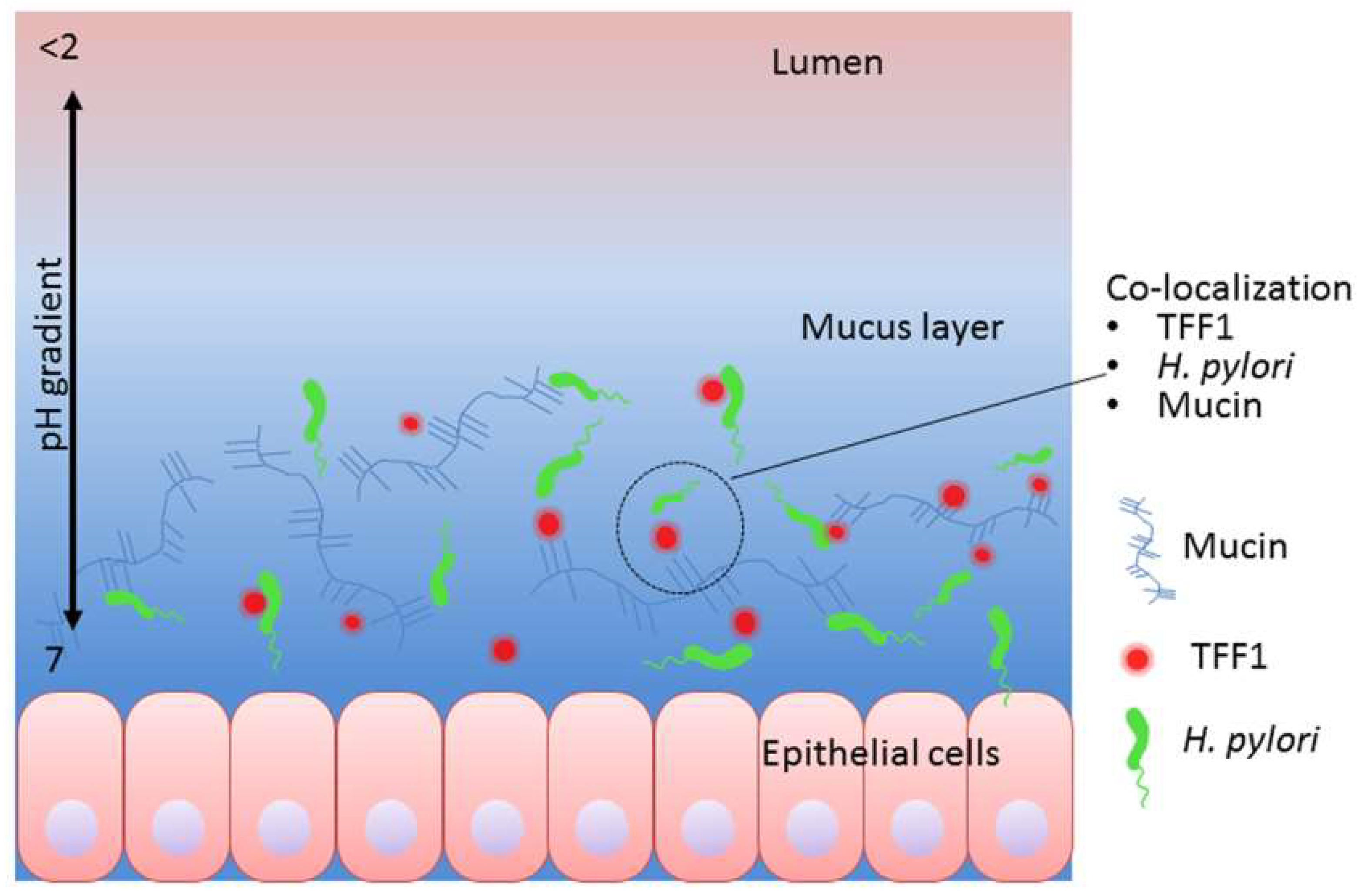
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Falush, D.; Wirth, T.; Linz, B.; Pritchard, J.K.; Stephens, M.; Kidd, M.; Blaser, M.J.; Graham, D.Y.; Vacher, S.; Perez-Perez, G.I.; et al. Traces of human migrations in Helicobacter pylori populations. Science 2003, 299, 1582–1585. [Google Scholar] [CrossRef] [PubMed]
- Rowland, M.; Daly, L.; Vaughan, M.; Higgins, A.; Bourke, B.; Drumm, B. Age-specific incidence of Helicobacter pylori. Gastroenterology 2006, 130, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Marshall, B.J.; Warren, J.R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1984, 1, 1311–1315. [Google Scholar] [CrossRef]
- Uemura, N.; Okamoto, S.; Yamamoto, S.; Matsumura, N.; Yamaguchi, S.; Yamakido, M.; Taniyama, K.; Sasaki, N.; Schlemper, R.J. Helicobacter pylori infection and the development of gastric cancer. N. Engl. J. Med. 2001, 345, 784–789. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer, World Health Organization. Schistosomes, liver flukes and Helicobacter pylori. IARC Monogr. Eval. Carcinog. Risks Hum. 1994, 61, 1–241. [Google Scholar]
- Thomsen, L.L.; Gavin, J.B.; Tasman-Jones, C. Relation of Helicobacter pylori to the human gastric mucosa in chronic gastritis of the antrum. Gut 1990, 31, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- Boren, T.; Falk, P.; Roth, K.A.; Larson, G.; Normark, S. Attachment of Helicobacter pylori to human gastric epithelium mediated by blood group antigens. Science 1993, 262, 1892–1895. [Google Scholar] [CrossRef] [PubMed]
- Van de Bovenkamp, J.H.; Mahdavi, J.; Korteland-Van Male, A.M.; Buller, H.A.; Einerhand, A.W.; Boren, T.; Dekker, J. The muc5ac glycoprotein is the primary receptor for Helicobacter pylori in the human stomach. Helicobacter 2003, 8, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Rossez, Y.; Gosset, P.; Boneca, I.G.; Magalhaes, A.; Ecobichon, C.; Reis, C.A.; Cieniewski-Bernard, C.; Joncquel Chevalier Curt, M.; Leonard, R.; Maes, E.; et al. The lacdinac-specific adhesin laba mediates adhesion of Helicobacter pylori to human gastric mucosa. J. Infect. Dis. 2014, 210, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, J.; Sonden, B.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Angstrom, J.; Larsson, T.; Teneberg, S.; Karlsson, K.A.; et al. Helicobacter pylori saba adhesin in persistent infection and chronic inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Ilver, D.; Arnqvist, A.; Ogren, J.; Frick, I.M.; Kersulyte, D.; Incecik, E.T.; Berg, D.E.; Covacci, A.; Engstrand, L.; Boren, T. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science 1998, 279, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Clyne, M.; Dillon, P.; Daly, S.; O’Kennedy, R.; May, F.E.B.; Westley, B.R.; Drumm, B. Helicobacter pylori interacts with the human single-domain trefoil protein tff1. Proc. Natl. Acad. Sci. USA 2004, 101, 7409–7414. [Google Scholar] [CrossRef] [PubMed]
- Reeves, E.P.; Ali, T.; Leonard, P.; Hearty, S.; O’Kennedy, R.; May, F.E.B.; Westley, B.R.; Josenhans, C.; Rust, M.; Suerbaum, S.; et al. Helicobacter pylori lipopolysaccharide interacts with TFF1 in a pH-dependent manner. Gastroenterology 2008, 135, 2043–2054. [Google Scholar] [CrossRef] [PubMed]
- Longman, R.J.; Douthwaite, J.; Sylvester, P.A.; Poulsom, R.; Corfield, A.P.; Thomas, M.G.; Wright, N.A. Coordinated localisation of mucins and trefoil peptides in the ulcer associated cell lineage and the gastrointestinal mucosa. Gut 2000, 47, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Newton, J.L.; Allen, A.; Westley, B.R.; May, F.E.B. The human trefoil peptide, TFF1, is present in different molecular forms that are intimately associated with mucus in normal stomach. Gut 2000, 46, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Westley, B.R.; Griffin, S.M.; May, F.E.B. Interaction between TFF1, a gastric tumor suppressor trefoil protein, and tfiz1, a brichos domain-containing protein with homology to SP-C. Biochemistry 2005, 44, 7967–7975. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, M.P.; Westley, B.R.; May, F.E.B. Homodimerization and hetero-oligomerization of the single-domain trefoil protein PNR-2/PS2 through cysteine 58. Biochem. J. 1997, 327, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; Westley, B.R.; May, F.E.B.; Feeney, J. The solution structure of the disulphide-linked homodimer of the human trefoil protein TFF1. FEBS Lett. 2001, 493, 70–74. [Google Scholar] [CrossRef]
- Ruchaud-Sparagano, M.H.; Westley, B.R.; May, F.E.B. The trefoil protein TFF1 is bound to muc5ac in human gastric mucosa. Cell. Mol. Life Sci. CMLS 2004, 61, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Dolan, B.; Naughton, J.; Tegtmeyer, N.; May, F.E.B.; Clyne, M. The interaction of Helicobacter pylori with the adherent mucus gel layer secreted by polarized HT29-MTX-E12 cells. PLoS ONE 2012, 7, e47300. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.; Windhager, L.; Rohrer, S.; Zeiller, M.; Karnholz, A.; Hoffmann, R.; Zimmer, R.; Haas, R. Strain-specific genes of Helicobacter pylori: Genome evolution driven by a novel Type iv secretion system and genomic island transfer. Nucleic Acids Res. 2010, 38, 6089–6101. [Google Scholar] [CrossRef] [PubMed]
- Tomb, J.F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Dougherty, B.A.; et al. The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature 1997, 388, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Alm, R.A.; Ling, L.S.; Moir, D.T.; King, B.L.; Brown, E.D.; Doig, P.C.; Smith, D.R.; Noonan, B.; Guild, B.C.; deJonge, B.L.; et al. Genomic-sequence comparison of two unrelated isolates of the human gastric pathogen Helicobacter pylori. Nature 1999, 397, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Baltrus, D.A.; Amieva, M.R.; Covacci, A.; Lowe, T.M.; Merrell, D.S.; Ottemann, K.M.; Stein, M.; Salama, N.R.; Guillemin, K. The complete genome sequence of Helicobacter pylori strain G27. J. Bacteriol. 2009, 191, 447–448. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Kopecko, D.J. Campylobacter jejuni 81-176 associates with microtubules and dynein during invasion of human intestinal cells. Infect. Immun. 1999, 67, 4171–4182. [Google Scholar] [PubMed]
- Korlath, J.A.; Osterholm, M.T.; Judy, L.A.; Forfang, J.C.; Robinson, R.A. A point-source outbreak of campylobacteriosis associated with consumption of raw milk. J. Infect. Dis. 1985, 152, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Parkhill, J.; Wren, B.W.; Mungall, K.; Ketley, J.M.; Churcher, C.; Basham, D.; Chillingworth, T.; Davies, R.M.; Feltwell, T.; Holroyd, S.; et al. The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 2000, 403, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, M.; Gerlach, J.Q.; Gough, R.; Gallagher, M.E.; Kane, M.; Carrington, S.D.; Joshi, L. Construction of a natural mucin microarray and interrogation for biologically relevant glyco-epitopes. Anal. Chem. 2012, 84, 3330–3338. [Google Scholar] [CrossRef] [PubMed]
- Naughton, J.A.; Marino, K.; Dolan, B.; Reid, C.; Gough, R.; Gallagher, M.E.; Kilcoyne, M.; Gerlach, J.Q.; Joshi, L.; Rudd, P.; et al. Divergent mechanisms of interaction of Helicobacter pylori and Campylobacter jejuni with mucus and mucins. Infect. Immun. 2013, 81, 2838–2850. [Google Scholar] [CrossRef] [PubMed]
- Rossez, Y.; Maes, E.; Lefebvre Darroman, T.; Gosset, P.; Ecobichon, C.; Joncquel Chevalier Curt, M.; Boneca, I.G.; Michalski, J.C.; Robbe-Masselot, C. Almost all human gastric mucin o-glycans harbor blood group a, b or h antigens and are potential binding sites for Helicobacter pylori. Glycobiology 2012, 22, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Earley, H.; Lennon, G.; Balfe, A.; Kilcoyne, M.; Clyne, M.; Joshi, L.; Carrington, S.; Martin, S.T.; Coffey, J.C.; Winter, D.C.; et al. A preliminary study examining the binding capacity of Akkermansia muciniphila and Desulfovibrio spp., to colonic mucin in health and ulcerative colitis. PLoS ONE 2015, 10, e0135280. [Google Scholar] [CrossRef] [PubMed]
- Behrens, I.; Stenberg, P.; Artursson, P.; Kissel, T. Transport of lipophilic drug molecules in a new mucus-secreting cell culture model based on HT29MTX cells. Pharm. Res. 2001, 18, 1138–1145. [Google Scholar] [CrossRef] [PubMed]
- Tom, B.H.; Rutzky, L.P.; Jakstys, M.M.; Oyasu, R.; Kaye, C.I.; Kahan, B.D. Human colonic adenocarcinoma cells. I. Establishment and description of a new line. In Vitro 1976, 12, 180–191. [Google Scholar] [CrossRef] [PubMed]
- University of Maryland, Hierarchical Clustering Explorer. Available online: http://www.cs.umd.edu/hcil/hce/ (accessed on 26 January 2017).
- Odenbreit, S.; Swoboda, K.; Barwig, I.; Ruhl, S.; Boren, T.; Koletzko, S.; Haas, R. Outer membrane protein expression profile in Helicobacter pylori clinical isolates. Infect. Immun. 2009, 77, 3782–3790. [Google Scholar] [CrossRef] [PubMed]
- Rio, M.C.; Bellocq, J.P.; Daniel, J.Y.; Tomasetto, C.; Lathe, R.; Chenard, M.P.; Batzenschlager, A.; Chambon, P. Breast cancer-associated ps2 protein: Synthesis and secretion by normal stomach mucosa. Science 1988, 241, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Yet, M.G.; Chin, C.C.; Wold, F. The covalent structure of individual n-linked glycopeptides from ovomucoid and asialofetuin. J. Biol. Chem. 1988, 263, 111–117. [Google Scholar] [PubMed]
- Tomasetto, C.; Masson, R.; Linares, J.L.; Wendling, C.; Lefebvre, O.; Chenard, M.P.; Rio, M.C. Ps2/tff1 interacts directly with the VWFC cysteine-rich domains of mucins. Gastroenterology 2000, 118, 70–80. [Google Scholar] [CrossRef]
- Desseyn, J.L.; Buisine, M.P.; Porchet, N.; Aubert, J.P.; Degand, P.; Laine, A. Evolutionary history of the 11p15 human mucin gene family. J. Mol. Evol. 1998, 46, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Joncquel Chevalier Curt, M.; Lecointe, K.; Mihalache, A.; Rossez, Y.; Gosset, P.; Leonard, R.; Robbe-Masselot, C. Alteration or adaptation, the two roads for human gastric mucin glycosylation infected by Helicobacter pylori. Glycobiology 2015, 25, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, J.; Yeh, J.C.; Misra, A.K.; Ito, S.; Katsuyama, T.; Fukuda, M. Expression cloning of a human α1,4-N-acetylglucosaminyltransferase that forms GlcNAcα1→4Galβ→R, a glycan specifically expressed in the gastric gland mucous cell-type mucin. Proc. Natl. Acad. Sci. USA 1999, 96, 8991–8996. [Google Scholar] [CrossRef] [PubMed]
- Kawakubo, M.; Ito, Y.; Okimura, Y.; Kobayashi, M.; Sakura, K.; Kasama, S.; Fukuda, M.N.; Fukuda, M.; Katsuyama, T.; Nakayama, J. Natural antibiotic function of a human gastric mucin against Helicobacter pylori infection. Science 2004, 305, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Wang, P.; Hoshino, H.; Ito, Y.; Kobayashi, M.; Nakayama, J.; Seeberger, P.H.; Fukuda, M. Alpha1,4glcnac-capped mucin-type o-glycan inhibits cholesterol alpha-glucosyltransferase from Helicobacter pylori and suppresses H. pylori growth. Glycobiology 2008, 18, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, F.G.; Bonar, D.; Schloerer, N.; Schroten, H. Human trefoil factor 2 is a lectin that binds alpha-glcnac-capped mucin glycans with antibiotic activity against Helicobacter pylori. J. Biol. Chem. 2014, 289, 27363–27375. [Google Scholar] [CrossRef] [PubMed]
- Soutto, M.; Chen, Z.; Katsha, A.M.; Romero-Gallo, J.; Krishna, U.S.; Piazuelo, M.B.; Washington, M.K.; Peek, R.M., Jr.; Belkhiri, A.; El-Rifai, W.M. Trefoil factor 1 expression suppresses Helicobacter pylori-induced inflammation in gastric carcinogenesis. Cancer 2015, 121, 4348–4358. [Google Scholar] [CrossRef] [PubMed]
- Soutto, M.; Romero-Gallo, J.; Krishna, U.; Piazuelo, M.B.; Washington, M.K.; Belkhiri, A.; Peek, R.M., Jr.; El-Rifai, W. Loss of tff1 promotes Helicobacter pylori-induced beta-catenin activation and gastric tumorigenesis. Oncotarget 2015, 6, 17911–17922. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Rho Value | p Value |
|---|---|---|
| H. pylori P12 | 0.82 | 0.002 |
| H. pylori 26695 | 0.74 | 0.01 |
| H. pylori G27 | 0.72 | 0.02 |
| H. pylori J99 | 0.82 | 0.004 |
| C. jejuni 11168 | 0.39 | 0.21 |
| C. jejuni 81-176 | 0.58 | 0.052 |
| Lectin | TFF1 | H. pylori P12 | H. pylori J99 | H. pylori 26695 | H. pylori G27 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| rho | p | rho | p | rho | p | rho | p | rho | p | |
| MAA | 0.636 | 0.03 | 0.734 | 0.009 | 0.436 | 0.18 | 0.636 | 0.04 | 0.591 | 0.06 |
| PNA | 0.566 | 0.06 | 0.769 | 0.005 | 0.464 | 0.15 | 0.491 | 0.13 | 0.473 | 0.14 |
| WFA | 0.308 | 0.33 | 0.482 | 0.11 | 0.209 | 0.54 | 0.536 | 0.09 | 0.273 | 0.42 |
| SBA | 0.392 | 0.21 | 0.587 | 0.05 | 0.300 | 0.37 | 0.636 | 0.04 | 0.327 | 0.33 |
| UEA-1 | −0.252 | 0.43 | −0.189 | 0.55 | 0.364 | 0.27 | −0.064 | 0.86 | 0.191 | 0.57 |
| GS-II | 0.811 | 0.002 | 0.769 | 0.005 | 0.791 | 0.006 | 0.718 | 0.01 | 0.800 | 0.005 |
| AIA | 0.454 | 0.14 | 0.559 | 0.06 | 0.454 | 0.16 | 0.500 | 0.12 | 0.482 | 0.14 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunne, C.; Naughton, J.; Duggan, G.; Loughrey, C.; Kilcoyne, M.; Joshi, L.; Carrington, S.; Earley, H.; Backert, S.; Robbe Masselot, C.; et al. Binding of Helicobacter pylori to Human Gastric Mucins Correlates with Binding of TFF1. Microorganisms 2018, 6, 44. https://doi.org/10.3390/microorganisms6020044
Dunne C, Naughton J, Duggan G, Loughrey C, Kilcoyne M, Joshi L, Carrington S, Earley H, Backert S, Robbe Masselot C, et al. Binding of Helicobacter pylori to Human Gastric Mucins Correlates with Binding of TFF1. Microorganisms. 2018; 6(2):44. https://doi.org/10.3390/microorganisms6020044
Chicago/Turabian StyleDunne, Ciara, Julie Naughton, Gina Duggan, Catherine Loughrey, Michelle Kilcoyne, Lokesh Joshi, Stephen Carrington, Helen Earley, Steffen Backert, Catherine Robbe Masselot, and et al. 2018. "Binding of Helicobacter pylori to Human Gastric Mucins Correlates with Binding of TFF1" Microorganisms 6, no. 2: 44. https://doi.org/10.3390/microorganisms6020044
APA StyleDunne, C., Naughton, J., Duggan, G., Loughrey, C., Kilcoyne, M., Joshi, L., Carrington, S., Earley, H., Backert, S., Robbe Masselot, C., May, F. E. B., & Clyne, M. (2018). Binding of Helicobacter pylori to Human Gastric Mucins Correlates with Binding of TFF1. Microorganisms, 6(2), 44. https://doi.org/10.3390/microorganisms6020044