Use of Exopolysaccharide-Synthesizing Lactic Acid Bacteria and Fat Replacers for Manufacturing Reduced-Fat Burrata Cheese: Microbiological Aspects and Sensory Evaluation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Manufacturing of Burrata Cheese
2.3. Compositional Analysis
2.4. Assessment of Primary Proteolysis
2.5. Cultivable Microbiota
2.6. Sensory Analysis
2.7. Extraction and Sequencing of Total Bacteria DNA
2.8. Statistical Analyses
3. Results
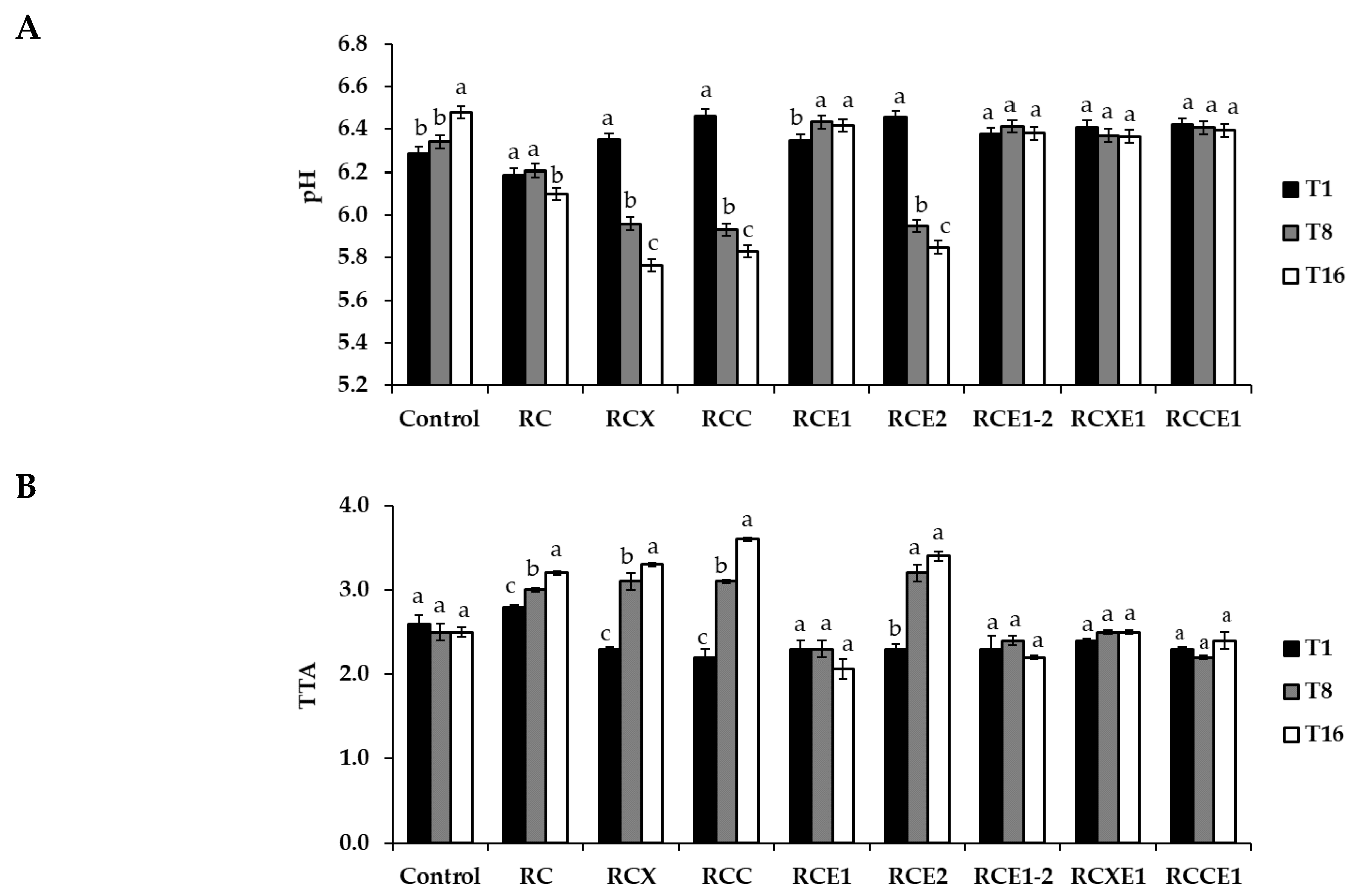
3.1. Compositional Analysis
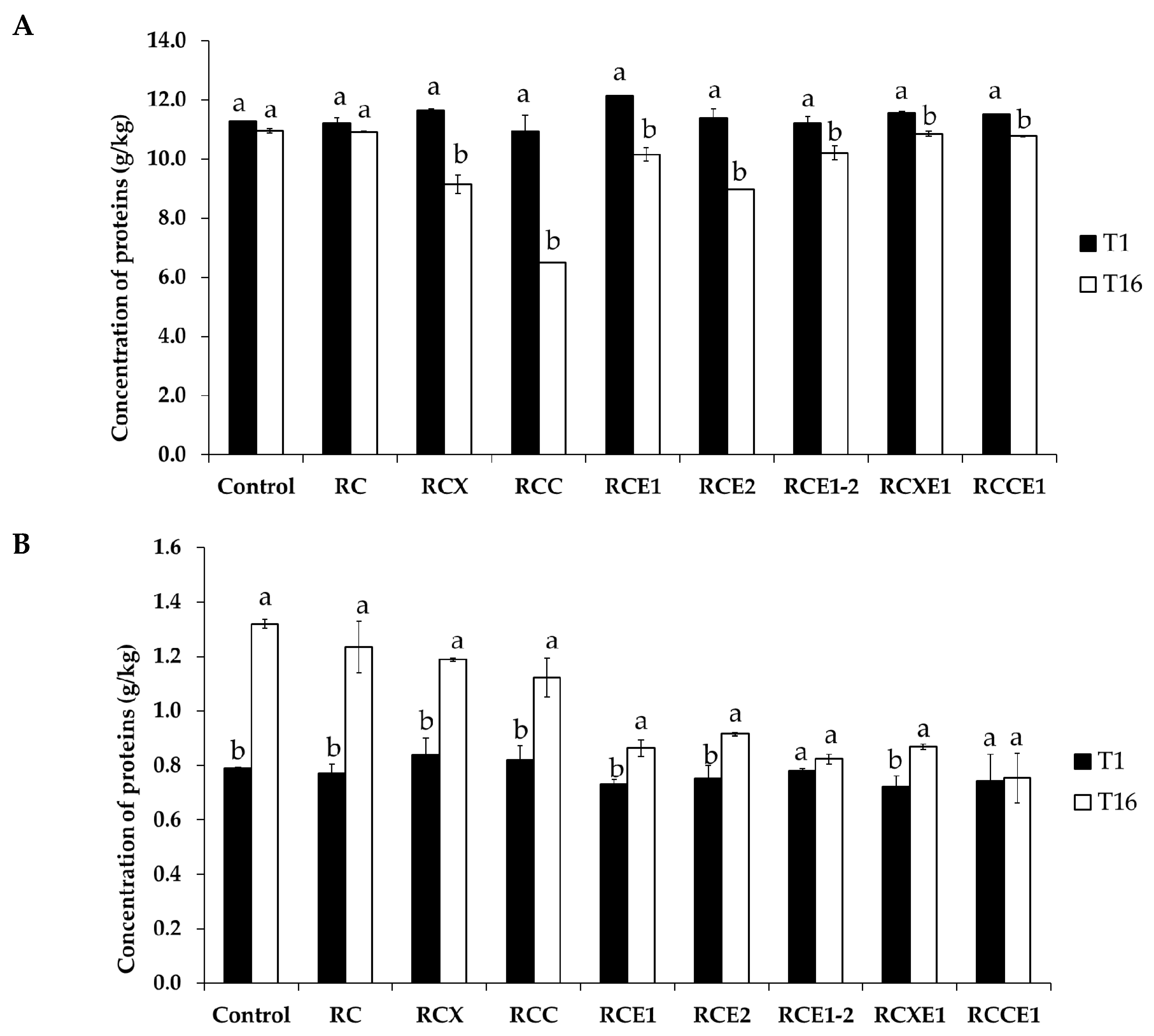
3.2. Assessment of Proteolysis
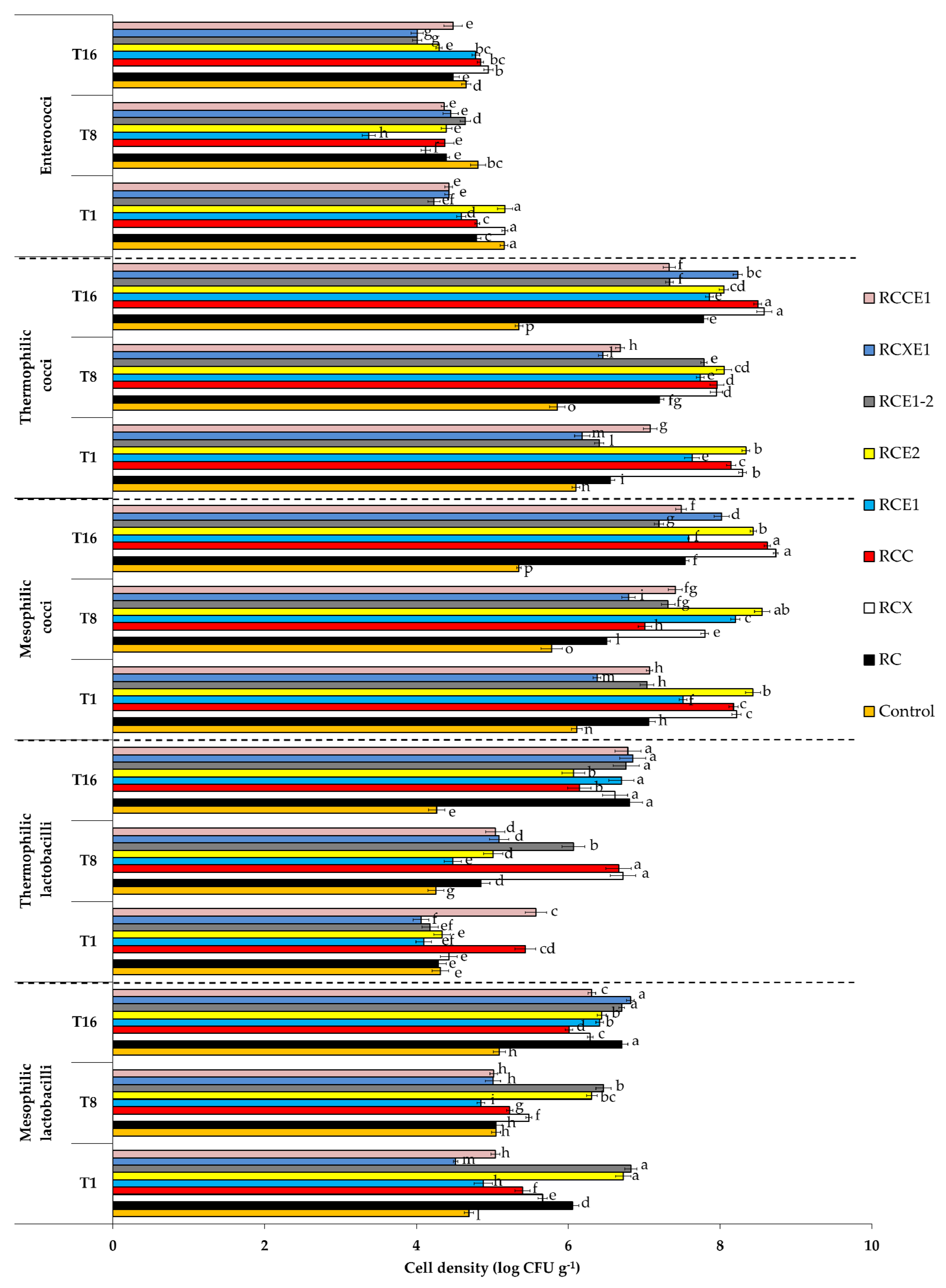
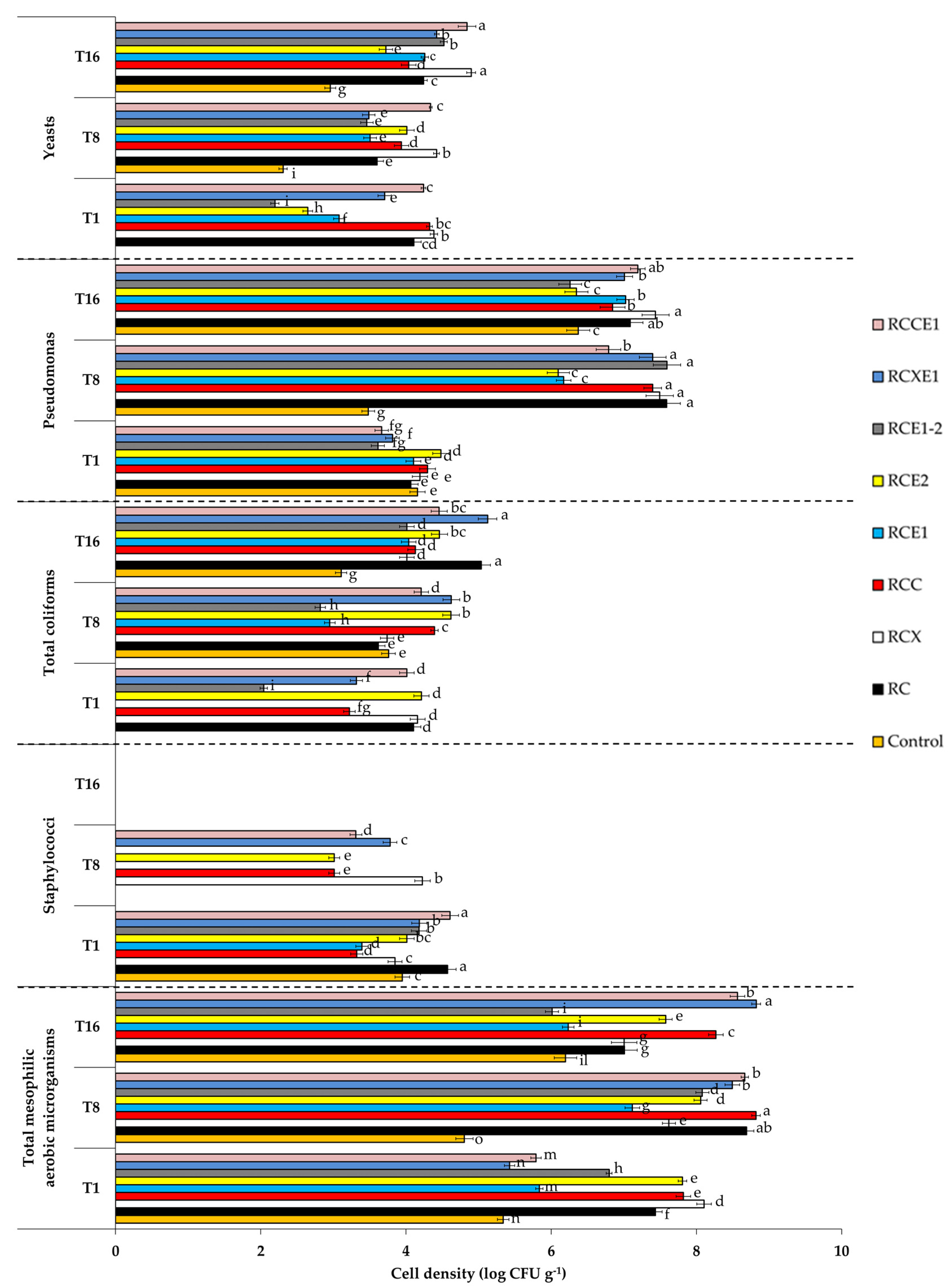
3.3. Cultivable Microbiota
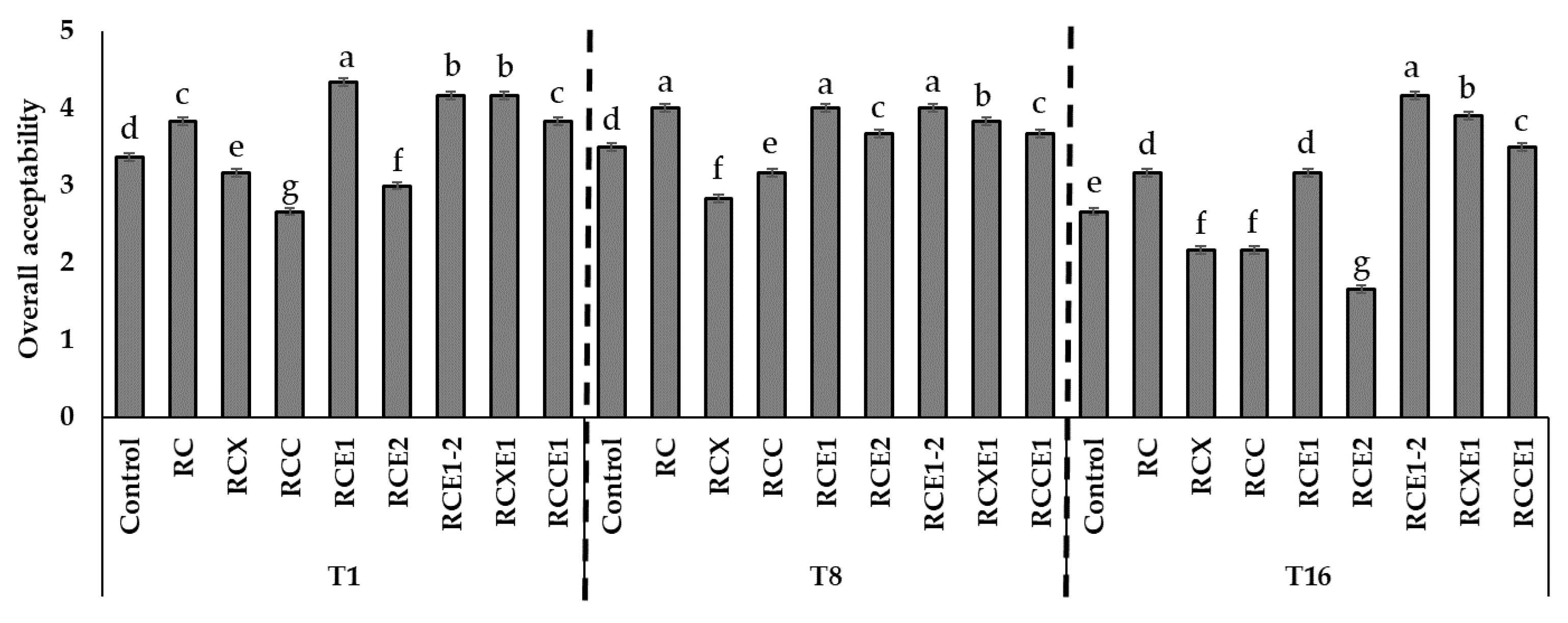
3.4. Panel Test
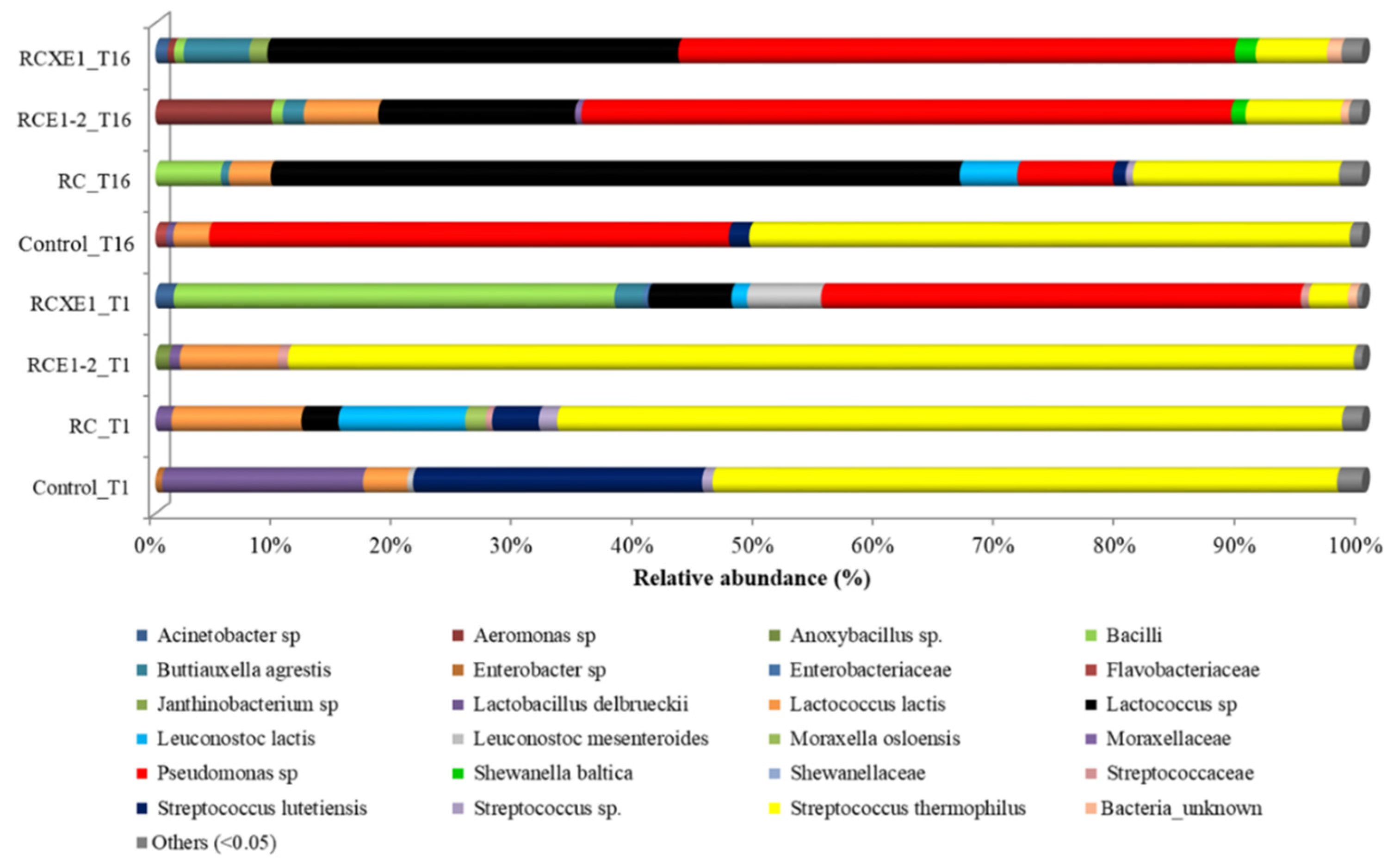
3.5. Burrata Microbiome
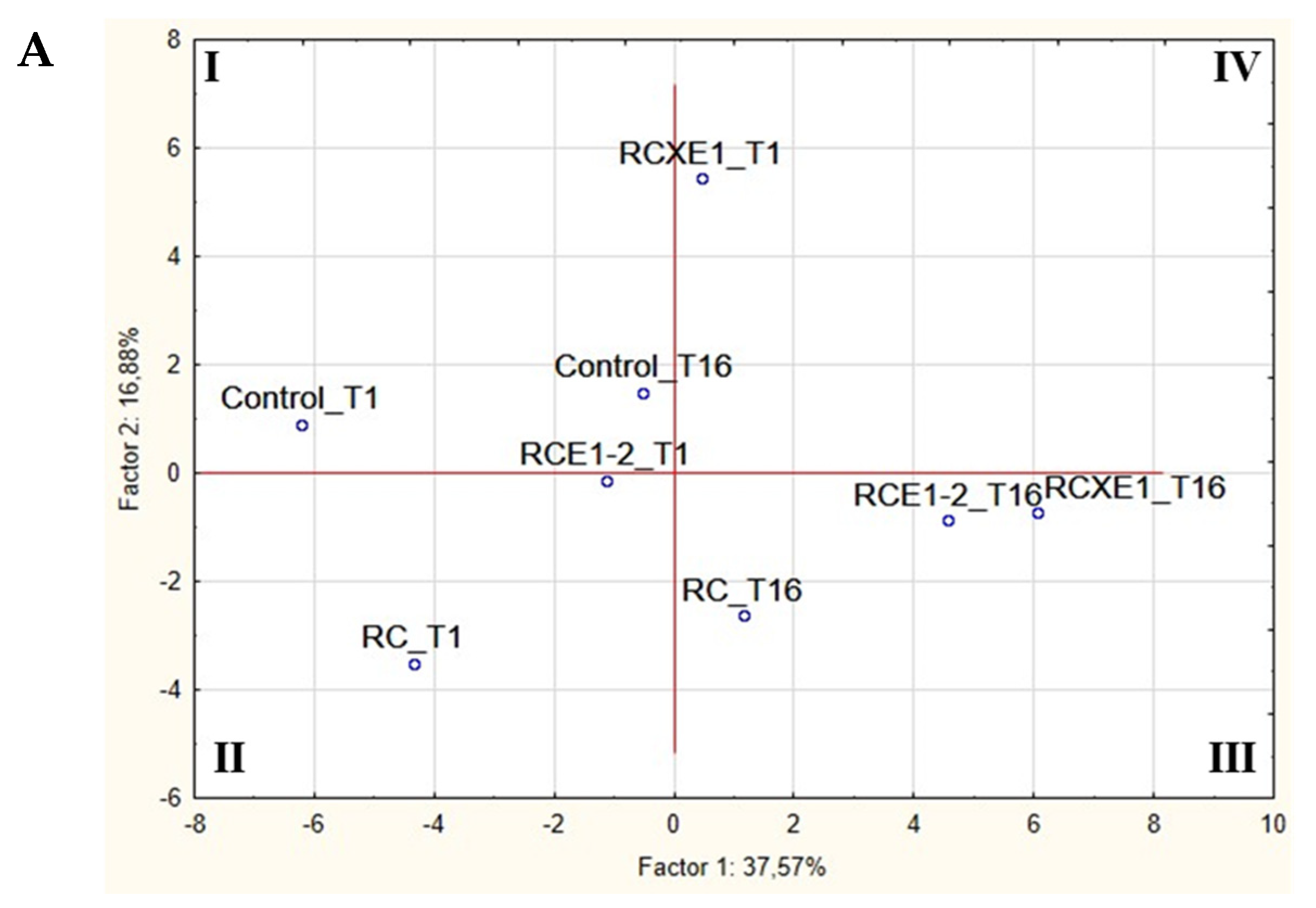
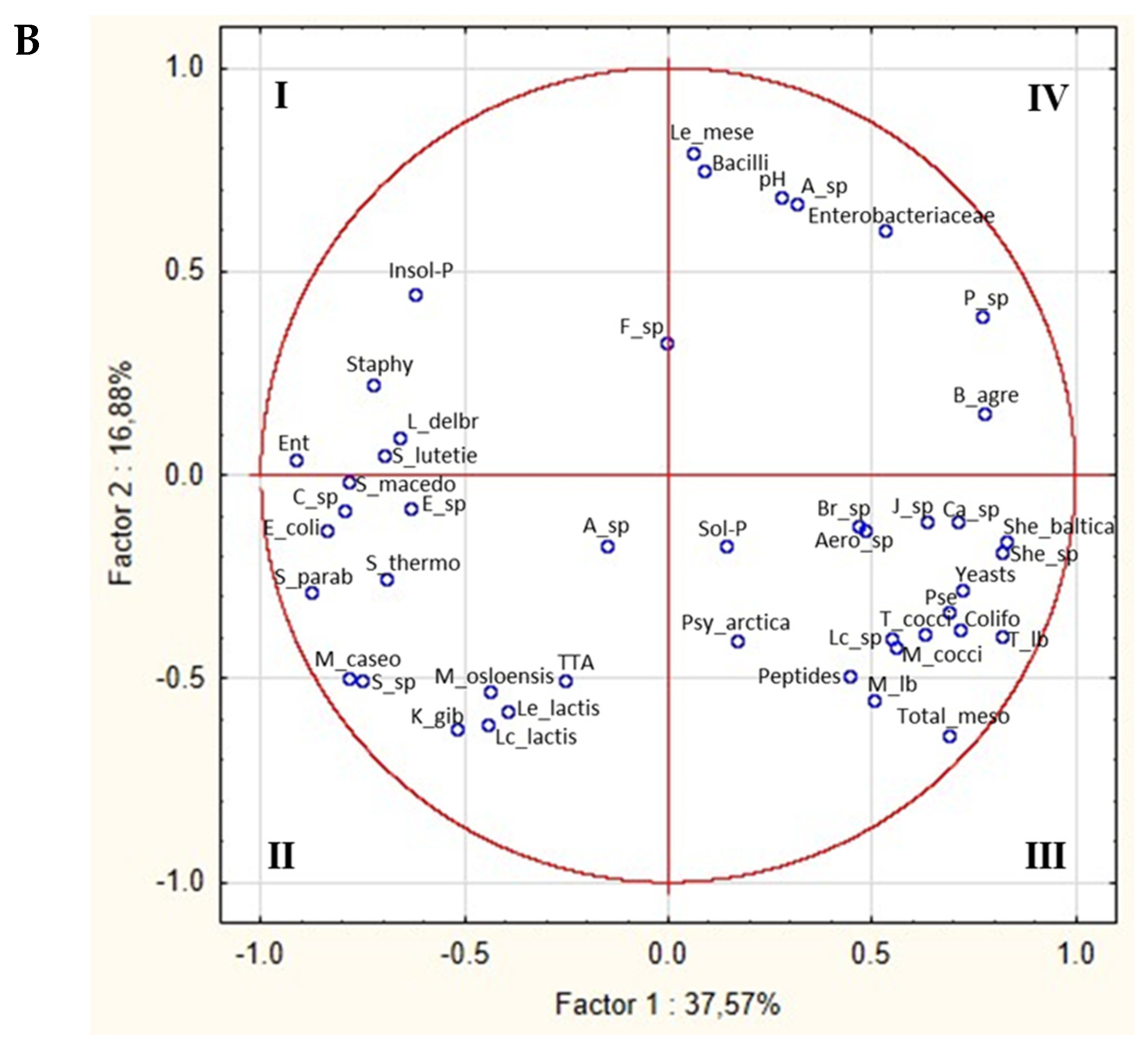
3.6. Correlations between Microbiota and Biochemical Characteristics of Burrata Cheeses
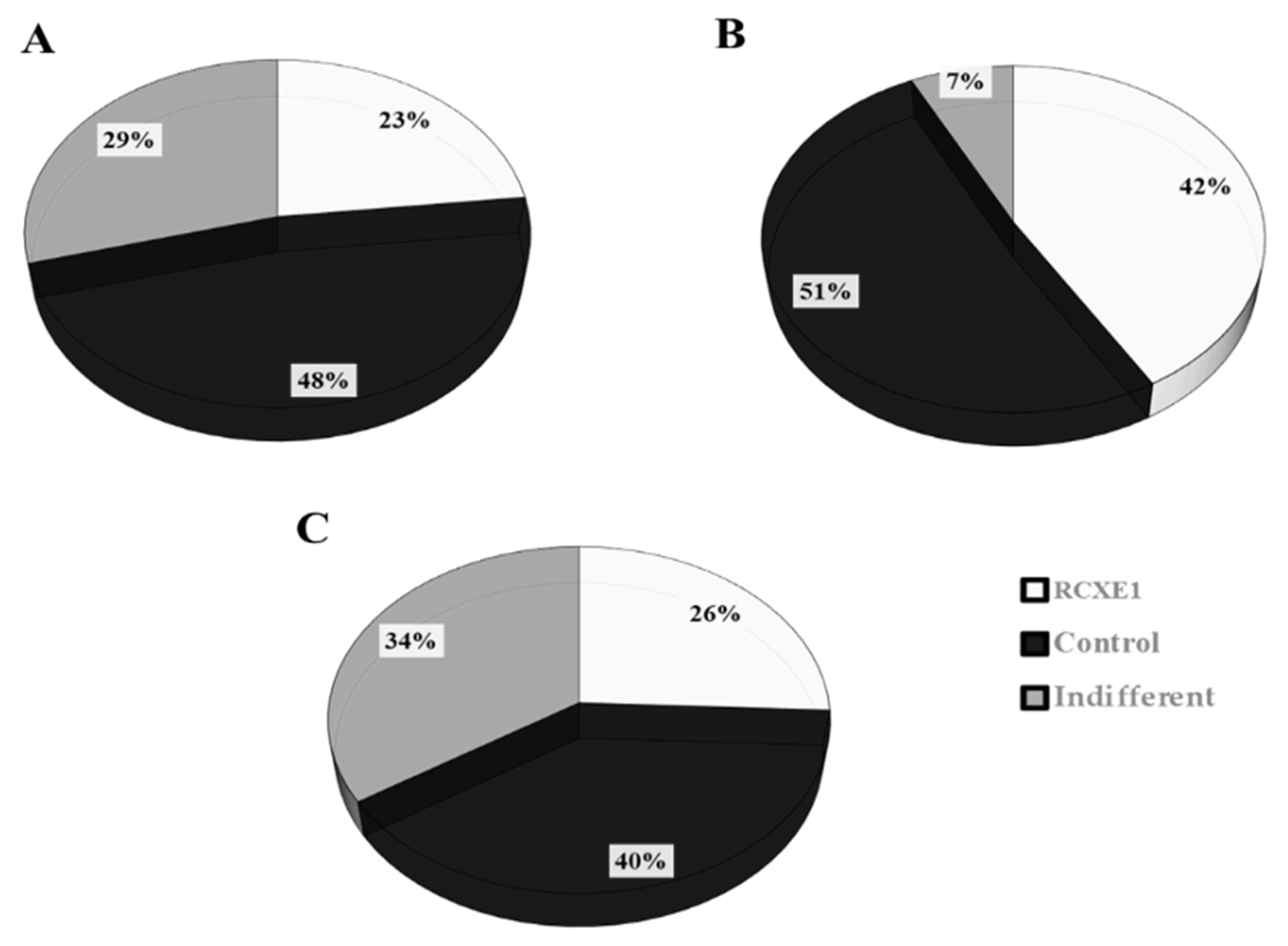
3.7. Consumer Test
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trani, A.; Gambacorta, G.; Gomes, T.F.; Loizzo, P.; Cassone, A.; Faccia, M. Production and characterisation of reduced-fat and PUFA-enriched Burrata cheese. J. Dairy Res. 2016, 83, 236–241. [Google Scholar] [CrossRef]
- Mipaaft. Diciannovesima Revisione dell’elenco dei Prodotti Agroalimentari Tradizionali. 2020. Available online: https://www.politicheagricole.it/flex/cm/pages/ServeBLOB.php/L/IT/IDPagina/13766 (accessed on 16 September 2020).
- European Commission. Regulations: Commission Implementing Regulation (EU) 2016/2103. L 327/16. In Official Journal of the European Union; European Union: Brussels, Belgium, 2 December 2016; Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=OJ:L:2016:327:FULL&from=SV (accessed on 20 September 2020).
- Minervini, F.; Conte, A.; Del Nobile, M.A.; Gobbetti, M.; De Angelis, M. Dietary fibers and protective lactobacilli drive Burrata cheese microbiome. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Conte, A.; Brescia, I.; Del Nobile, M.A. Lysozyme/EDTA disodium salt and modified-atmosphere packaging to prolong the shelf life of Burrata cheese. J. Dairy Sci. 2011, 94, 5289–5297. [Google Scholar] [CrossRef]
- Feeney, E.L.; Nugent, A.P.; Mc Nulty, B.; Walton, J.; Flynn, A.; Gibney, E.R. An overview of the contribution of dairy and cheese intakes to nutrient intakes in the Irish diet: Results from the National Adult Nutrition Survery. Br. J. Nutr. 2016, 115, 709–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaucheron, F. Milk and dairy products: A unique micronutrient combination. J. Am. Coll. Nutr. 2011, 30, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Bates, B.; Lennox, A.; Prentice, A.; Bates, C.J. National Diet and Nutrition Survey: Results from Years 1–4 (Combined) of the Rolling Programme (2008/2009–2011/12); Public Health England: London, UK, 2014. [Google Scholar]
- Fulgoni, V.L.; Keast, D.R.; Auestad, N.; Quann, E.E. Nutrients from dairy foods are difficult to replace in diets of Americans: Food pattern modeling and an analysis of the National Health and Nutrition Examination Survey 2003–2006. Nutr. Res. 2011, 31, 759–765. [Google Scholar] [CrossRef]
- Iannotti, L.; Muehlhoff, E.; Mcmahon, D. Review of milk and dairy programmes affecting nutrition. J. Dev. Effect 2013, 5, 82–115. [Google Scholar] [CrossRef]
- Hjerpsted, J.; Tholstrup, T. Cheese and cardiovascular disease risk: A review of the evidence and discussion of possible mechanisms. Crit. Rev. Food Sci. Nutr. 2015, 56, 1389–1403. [Google Scholar] [CrossRef]
- Gulhane, M.; Murray, L.; Lourie, R.; Tong, H.; Sheng, Y.H.; Wang, R.; Kang, A.; Schreiber, V.; Wong, K.Y.; Magor, G.; et al. High fat diets induce colonic epithelial cell stress and inflammation that is reversed by IL-22. Sci. Rep. 2016, 6, 28990. [Google Scholar] [CrossRef]
- Patterson, R.E.; Rock, C.L.; Kerr, J.; Natarajan, L.; Marshall, S.J.; Pakiz, B.; Cadmus-Bertram, L.A. Metabolism and breast cancer risk: Frontiers in research and practice. J. Acad. Nutr. Diet. 2013, 113, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Butler, L.M.; Wang, R.; Koh, W.P.; Stern, M.C.; Yuan, J.M.; Yu, M.C. Marine n-3 and saturated fatty acids in relation to risk of colorectal cancer in Singapore Chinese: A prospective study. Int. J. Cancer 2009, 124, 678–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arem, H.; Irwin, M.L. Obesity and endometrial cancer survival: A systematic review. Int. J. Obes. 2013, 37, 634–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiebaut, A.C.M.; Kipnis, V.; Chang, S.C.; Subar, A.F.; Thompson, F.E.; Rosenberg, P.S.; Hollenbeck, A.R.; Leitzmann, M.; Schatzkin, A. Dietary fat and postmenopausal invasive breast cancer in the National Institutes of Health-AARP Diet and Health Study Cohort. JNCI J. Nat. Cancer I. 2007, 99, 451–462. [Google Scholar] [CrossRef]
- Jansen, R.J.; Robinson, D.P.; Frank, R.D.; Anderson, K.E.; Bamlet, W.R.; Oberg, A.L.; Rabe, K.G.; Olson, J.E.; Sinha, R.; Petersen, G.M.; et al. Fatty acids found in dairy, protein and unsaturated fatty acids are associated with risk of pancreatic cancer in a case-control study. Int. J. Cancer 2013, 134, 1935–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luck, H.; Tsai, S.; Chung, J.; Clemente-Casares, X.; Ghazarian, M.; Revelo, S.X.; Lei, H.; Luk, C.T.; Yu, S.S.; Surendra, A.; et al. Regulation of obesity-related insulin resistance with gut anti-inflammatory agents. Cell Metab. 2015, 21, 527. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Medina, M.; Denizot, J.; Dreux, N.; Robin, F.; Billard, E.; Bonnet, R.; Darfeuille-Michaud, A.; Barnich, N. Western diet induces dysbiosis with increased E. coli in CEABAC10 mice, alters host barrier function favouring AIEC colonisation. Gut 2014, 63, 116. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470. [Google Scholar] [CrossRef] [Green Version]
- Crean, A.J.; Senior, A.M. High-fat diets reduce male reproductive success in animal models: A systematic review and meta-analysis. Obes. Rev. 2019, 20, 921–933. [Google Scholar] [CrossRef]
- Song, M.; Garrett, W.S.; Chan, A.T. Nutrients, foods, and colorectal cancer prevention. Gastroenterology 2015, 148, 1244–1260. [Google Scholar] [CrossRef] [Green Version]
- McDonald, B.E. Dietary cholesterol: Is there a problem? In IDF Bullettin; International Dairy Federation: Brussels, Belgium, 2000; Volume 355, pp. 22–24. [Google Scholar]
- León-Muñoz, L.M.; Guallar-Castillón, P.; Graciani, A.; López-García, E.; Mesas, A.E.; Aguilera, M.T.; Banegas, J.R.; Rodríguez-Artalejo, F. Adherence to the Mediterranean diet pattern has declined in Spanish adults. J. Nutr. 2012, 142, 1843–1850. [Google Scholar] [CrossRef] [Green Version]
- Drake, M.A.; Swanson, B.G. Reduced and low-fat cheese technology: A review. Trends Food Sci. Technol. 1995, 6, 366–369. [Google Scholar] [CrossRef]
- Banks, J.M. The technology of low-fat cheese manufacture. Int. J. Dairy Technol. 2004, 57, 199–207. [Google Scholar] [CrossRef]
- Juan, B.; Zamora, A.; Quintana, F.; Guamis, B.; Trujillo, A.J. Effect of inulin addition on the sensorial properties of reduced-fat fresh cheese. Int. J. Dairy Technol. 2013, 66, 478–483. [Google Scholar] [CrossRef]
- Koca, N.; Metin, M. Textural, melting and sensory properties of low-fat fresh Kashar cheeses produced by using fat replacers. Int. Dairy J. 2004, 14, 365–373. [Google Scholar] [CrossRef]
- Costa, N.E.; O’Callaghan, D.J.; Mateo, M.J.; Chaurin, V.; Castillo, M.; Hannon, J.A.; McSweeney, P.L.H.; Beresford, T.P. Influence of an exopolysaccharide produced by a starter on milk coagulation and curd syneresis. Int. Dairy J. 2011, 22, 48–57. [Google Scholar] [CrossRef]
- Sánchez-Macías, D.; Fresno, M.; Moreno-Indias, I.; Castro, N.; Morales-delaNuez, A.; Álvarez, S.; Argüello, A. Physicochemical analysis of full-fat, reduced-fat, and low-fat artisan-style goat cheese1. J. Dairy Sci. 2010, 93, 3950–3956. [Google Scholar] [CrossRef]
- Jameson, G.W. Dietary cheeses: Low fat, low salt. Food Technol. Aust. 1987, 39, 99–101. [Google Scholar]
- McGregor, J.U.; White, C.H. Effect of enzyme treatment and ultrafiltration on the quality of low-fat Cheddar cheese. J. Dairy Sci. 1990, 73, 571–578. [Google Scholar] [CrossRef]
- Mistry, V.V.; Metzger, L.E.; Maubois, J.L. Use of ultrafiltered sweet buttermilk in the manufacture of reduced fat Cheddar cheese. J. Dairy Sci. 1996, 79, 1137–1145. [Google Scholar] [CrossRef]
- Mistry, V.V. Low-fat cheese technology. Int. Dairy J. 2001, 11, 413–422. [Google Scholar] [CrossRef]
- Li, H.; Yu, H.; Liu, Y.; Wang, Y.; Li, H.; Yu, J. The use of inulin, maltitol and lecithin as fat replacers and plasticizers in a model reduced fat mozzarella cheese like product. J. Sci. Food Agric. 2019, 99, 5586–5593. [Google Scholar] [CrossRef]
- Khanal, B.K.S.; Bhandari, B.; Prakash, S.; Liu, D.; Zhou, P.; Bansal, N. Modifying textural and microstructural properties of low-fat Cheddar cheese using sodium alginate. Food Hydrocoll. 2018, 83, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wu, T.; Fang, X.; Yang, Z. Manufacture of low-fat Cheddar cheese by exopolysaccharide-producing Lactobacillus plantarum JLK0142 and its functional properties. J. Dairy Sci. 2019, 102, 3825–3838. [Google Scholar] [CrossRef] [PubMed]
- Oberg, E.N.; Oberg, C.J.; Motawee, M.M.; Martini, S.; McMahon, D.J. Increasing stringiness of low-fat mozzarella string cheese using polysaccharides. J. Dairy Sci. 2015, 98, 4243–4254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cagno, R.; De Pasquale, I.; De Angelis, M.; Buchin, S.; Rizzello, C.G.; Gobbetti, M. Use of microparticulated whey protein concentrate, exopolysaccharide-producing Streptococcus thermophilus, and adjunct cultures for making low-fat Italian Caciotta-type cheese. J. Dairy Sci. 2014, 97, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Baldini, M.; Fabietti, F.; Giammarioli, S.; Onori, R.; Orefice, L.; Stacchini, A. Metodi di Analisi Utilizzati per il Controllo Chimico Degli Alimenti; Rapporti ISTISAN 96/34; Rapporti ISTISAN: Roma, Italy, 1996. [Google Scholar]
- ISO. ISO 3433 Cheese-Determination of Fat Content—Van Gulik Method; International Organization for Standardization; ISO: Geneva, Switzerland, 2008. [Google Scholar]
- Kuchroo, C.N.; Fox, P.F. Soluble nitrogen in Cheddar cheese: Comparison of extraction procedures. Milchwissenschaft 1982, 37, 331–335. [Google Scholar]
- Andrews, A.T.; Alichanidis, E. Proteolysis of caseins and the proteose-peptone fraction of bovine milk. J. Dairy Res. 1983, 50, 275–290. [Google Scholar] [CrossRef]
- Blakesley, R.W.; Boezi, J.A. A new staining technique for proteins in polyacrylamide gels using Coomassie brilliant blue G250. Anal. Biochem. 1977, 82, 580–582. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Minervini, F.; Siragusa, S.; Faccia, M.; Dal Bello, F.; Gobbetti, M.; De Angelis, M. Manufacture of fior di latte cheese by incorporation of probiotic lactobacilli. J. Dairy Sci. 2012, 95, 508–520. [Google Scholar] [CrossRef] [Green Version]
- Coppola, S.; Villani, F.; Coppola, R.; Parente, E. Comparison of different starter systems for water-buffalo mozzarella cheese manufacture. Lait 1990, 70, 411–423. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, M.; de Candia, S.; Calasso, M.P.; Faccia, M.; Guinee, T.P.; Simonetti, M.C.; Gobbetti, M. Selection and use of autochthonous multiple strain cultures for the manufacture of high-moisture traditional mozzarella cheese. Int. J. Food Microbiol. 2008, 125, 123–132. [Google Scholar] [CrossRef] [PubMed]
- ISO Sensory Analysis. General Guidance for the Design of Test Rooms; ISO 8589; International Organization of Standardization: Geneva, Switzerland, 1988. [Google Scholar]
- Handl, S.; Dowd, S.E.; Garcia-Mazcorro, J.F.; Steiner, J.M.; Suchodolski, J.S. Massive parallel 16S rRNA gene pyrosequencing reveals highly diverse fecal bacterial and fungal communities in healthy dogs and cats. FEMS Microbiol. Ecol. 2011, 76, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhoads, D.D.; Wolcott, R.D.; Sun, Y.; Dowd, S.E. Comparison of culture and molecular identification of bacteria in chronic wounds. Int. J. Mol. Sci. 2012, 13, 2535–2550. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreotti, R.; Pérez de León, A.; Dowd, S.E.; Guerrero, F.D.; Bendele, K.G.; Scoles, G. Assessment of bacterial diversity in the cattle tick Rhipicephalus (Boophilus) microplus through tag-encoded pyrosequencing. BMC Microbiol. 2011, 11. [Google Scholar] [CrossRef] [Green Version]
- Hooper, L.; Martin, N.; Abdelhamid, A.; Davey, S.G. Reduction in saturated fat intake for cardiovascular disease. Cochrane Database Syst. Rev. 2015. [Google Scholar] [CrossRef]
- World Health Organization. Diet, Nutrition and the Prevention of Chronic Diseases: Report of a Joint WHO/FAO Expert Consultation; World Health Organization: Geneva, Switzerland, 2003; Available online: https://www.who.int/dietphysicalactivity/publications/trs916/en/ (accessed on 20 September 2020).
- Food and Agriculture Organization. Fats and Fatty Acids in Human Nutrition: Report of an Expert Consultation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010; Available online: https://www.who.int/nutrition/publications/nutrientrequirements/fatsandfattyacids_humannutrition/en/ (accessed on 20 September 2020).
- Bimbo, F.; Bonanno, A.; Nocella, G.; Viscecchia, R.; Nardone, G.; De Devitiis, B.; Carlucci, D. Consumers’ acceptance and preferences for nutrition-modified and functional dairy products: A systematic review. Appetite 2017, 113, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Henneberry, S.; O’Sullivan, M.G.; Kilcawley, K.N.; Kelly, P.M.; Wilkinson, M.G.; Guinee, T.P. Sensory quality of unheated and heated Mozzarella-style cheeses with different fat, salt and calcium levels. Int. J. Dairy Technol. 2016, 69, 38–50. [Google Scholar] [CrossRef]
- Rogers, N.; McMahon, D.J.; Daubert, C.R.; Berry, T.; Foegeding, E.A. Rheological properties and microstructure of Cheddar cheese made with different fat contents. J. Dairy Sci. 2010, 93, 4565–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zisu, B. Impact of Preacidification, Fat Replacers and Exopolysaccharide Producing Starter Cultures on Functionality of Low-Fat Mozzarella Cheese. Ph.D. Thesis, Victoria University, Victoria, Australia, 2005. Available online: http://vuir.vu.edu.au/16090/3/ZISU%20Bogdan-thesis_nosignature.pdf (accessed on 20 September 2020).
- Gobbetti, M.; Di Cagno, R.; Calasso, M.; Neviani, E.; Fox, P.F.; De Angelis, M. Drivers that establish and assemble the lactic acid bacteria biota in cheeses. Trends Food Sci. Technol. 2018, 78, 244–254. [Google Scholar] [CrossRef]
- Kamimura, B.A.; De Filippis, F.; Sant’Ana, A.S.; Ercolini, D. Large-scale mapping of microbial diversity in artisanal Brazilian cheeses. Food Microbiol. 2019, 80, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Kavas, G.; Oysun, G.; Kinik, O.; Uysal, H. Effect of some fat replacers on chemical, physical and sensory attributes of low-fat white pickled cheese. Food Chem. 2004, 88, 381–388. [Google Scholar] [CrossRef]
- Oliveira, M.M.; Almeida, K.E.; Damin, M.R.; Rochat, T.; Gratadoux, J.J.; Miyoshi, A.; Langella, P.; Azevedo, V. Behavior and viability of spontaneous oxidative stress-resistant Lactococcus lactis mutants in experimental fermented milk processing. Genet. Mol. Res. 2009, 8, 840–847. [Google Scholar] [CrossRef]
- Kives, J.; Guadarrama, D.; Orgaz, B.; Rivera-Sen, A.; Vazquez, J.; SanJose, C. Interactions in biofilms of Lactococcus lactis ssp. cremoris and Pseudomonas fluorescens cultured in cold UHT milk. J. Dairy Sci. 2005, 88, 4165–4171. [Google Scholar] [CrossRef]
- Calasso, M.; Ercolini, D.; Mancini, L.; Stellato, G.; Minervini, F.; Di Cagno, R.; De Angelis, M.; Gobbetti, M. Relationships among house, rind and core microbiotas during manufacture of traditional Italian cheeses at the same dairy plant. Food Microbiol. 2016, 54, 115–126. [Google Scholar] [CrossRef]
- Calasso, M.; Minervini, F.; De Filippis, F.; Ercolini, D.; De Angelis, M.; Gobbetti, M. Attenuated Lactococcus lactis and surface bacteria as tools for conditioning the microbiota and driving the ripening of semisoft Caciotta cheese. Appl. Environ. Microbiol. 2019, 86, e02165-19. [Google Scholar] [CrossRef] [Green Version]
- Carminati, D.; Bonvini, B.; Rossetti, L.; Zago, M.; Tidona, F.; Giraffa, G. Investigation on the presence of blue pigment-producing Pseudomonas strains along a production line of fresh mozzarella cheese. Food Control 2019, 100, 321–328. [Google Scholar] [CrossRef]
- Marino, M.; Dubsky de Wittenau, G.; Saccà, E.; Cattonaro, F.; Spadotto, A.; Innocente, N.; Radovic, S.; Piasentier, E.; Marroni, F. Metagenomic profiles of different types of Italian high-moisture Mozzarella cheese. Food Microbiol. 2019, 79, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Food Labeling: General Requirements for Health Claims for Food. Federal Register 1993, 58, 2478–2536.
- Drake, M.A. Modern sensory practices. In The Sensory Evaluation of Dairy Products; Clark, S., Costello, M., Drake, M.A., Bodyfelt, F., Eds.; Springer: New York, NY, USA, 2009; pp. 505–530. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Burrata Cheese Variant | Carbohydrates | Proteins | Total Fat | Moisture | Sodium |
|---|---|---|---|---|---|
| Control | 1.2 ± 0.06 a | 10.4 ± 0.03 c | 21.6 ± 0.05 a | 65.4 ± 0.90 c | 0.50 ± 0.02 a |
| RC | 0.8 ± 0.03 b | 11.6 ± 0.01 a | 16.0 ± 0.02 b | 70.5 ± 1.30 b | 0.14 ± 0.02 c |
| RCX | 1.1 ± 0.09 a | 10.5 ± 0.03 bc | 10.0 ± 0.05 e | 72.0 ± 1.40 ab | 0.16 ± 0.02 bc |
| RCC | 1.2 ± 0.06 a | 10.0 ± 0.01 d | 10.6 ± 0.03 e | 71.5 ± 1.20 b | 0.20 ± 0.02 b |
| RCE1 | 0.8 ± 0.05 b | 11.0 ± 0.03 b | 13.0 ± 0.05 d | 70.3 ± 1.40 b | 0.14 ± 0.03 c |
| RCE2 | 1.0 ± 0.05 a | 11.2 ± 0.02 b | 14.5 ± 0.04 c | 70.4 ± 1.20 b | 0.20 ± 0.01 b |
| RCE1-2 | 1.1 ± 0.06 a | 11.0 ± 0.01 b | 14.5 ± 0.05 c | 72.3 ± 1.40 ab | 0.20 ± 0.02 b |
| RCXE1 | 1.2 ± 0.05 a | 10.6 ± 0.01 c | 11.0 ± 0.04 e | 75.7 ± 1.20 a | 0.16 ± 0.03 bc |
| RCCE1 | 1.2 ± 0.07 a | 10.6 ± 0.03 c | 11.5 ± 0.05 e | 73.6 ± 1.20 ab | 0.16 ± 0.03 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costantino, G.; Calasso, M.; Minervini, F.; De Angelis, M. Use of Exopolysaccharide-Synthesizing Lactic Acid Bacteria and Fat Replacers for Manufacturing Reduced-Fat Burrata Cheese: Microbiological Aspects and Sensory Evaluation. Microorganisms 2020, 8, 1618. https://doi.org/10.3390/microorganisms8101618
Costantino G, Calasso M, Minervini F, De Angelis M. Use of Exopolysaccharide-Synthesizing Lactic Acid Bacteria and Fat Replacers for Manufacturing Reduced-Fat Burrata Cheese: Microbiological Aspects and Sensory Evaluation. Microorganisms. 2020; 8(10):1618. https://doi.org/10.3390/microorganisms8101618
Chicago/Turabian StyleCostantino, Giuseppe, Maria Calasso, Fabio Minervini, and Maria De Angelis. 2020. "Use of Exopolysaccharide-Synthesizing Lactic Acid Bacteria and Fat Replacers for Manufacturing Reduced-Fat Burrata Cheese: Microbiological Aspects and Sensory Evaluation" Microorganisms 8, no. 10: 1618. https://doi.org/10.3390/microorganisms8101618
APA StyleCostantino, G., Calasso, M., Minervini, F., & De Angelis, M. (2020). Use of Exopolysaccharide-Synthesizing Lactic Acid Bacteria and Fat Replacers for Manufacturing Reduced-Fat Burrata Cheese: Microbiological Aspects and Sensory Evaluation. Microorganisms, 8(10), 1618. https://doi.org/10.3390/microorganisms8101618