Composted Sewage Sludge Influences the Microbiome and Persistence of Human Pathogens in Soil

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Sewage Sludge Composting
2.3. Microcosm Experiment
2.4. DNA Extraction, 16S rRNA Gene Amplification and Amplicon Sequencing
2.5. Sequence Analysis
2.6. Detection of Salmonella enterica in Soil and Plant Tissues
3. Results
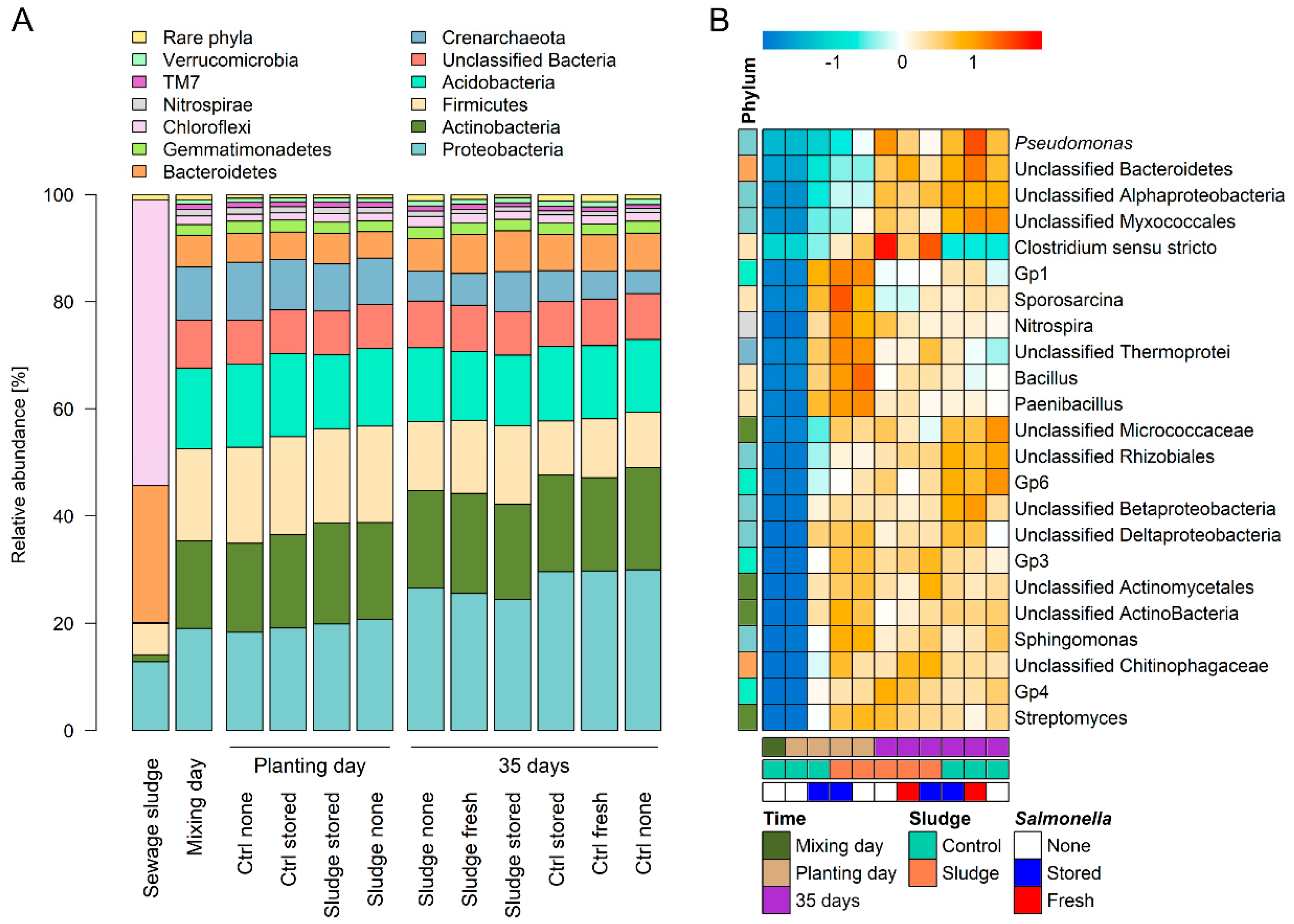
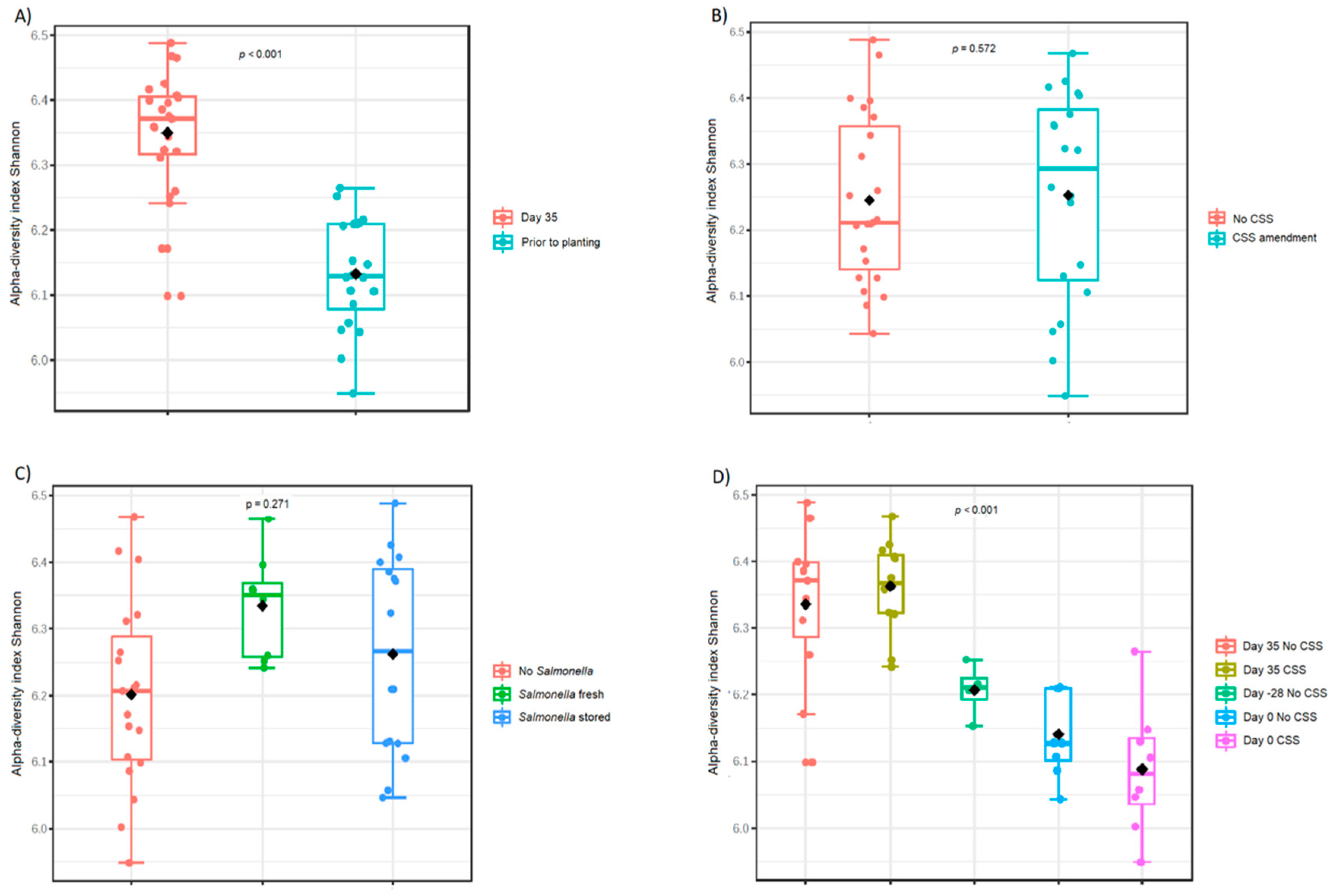
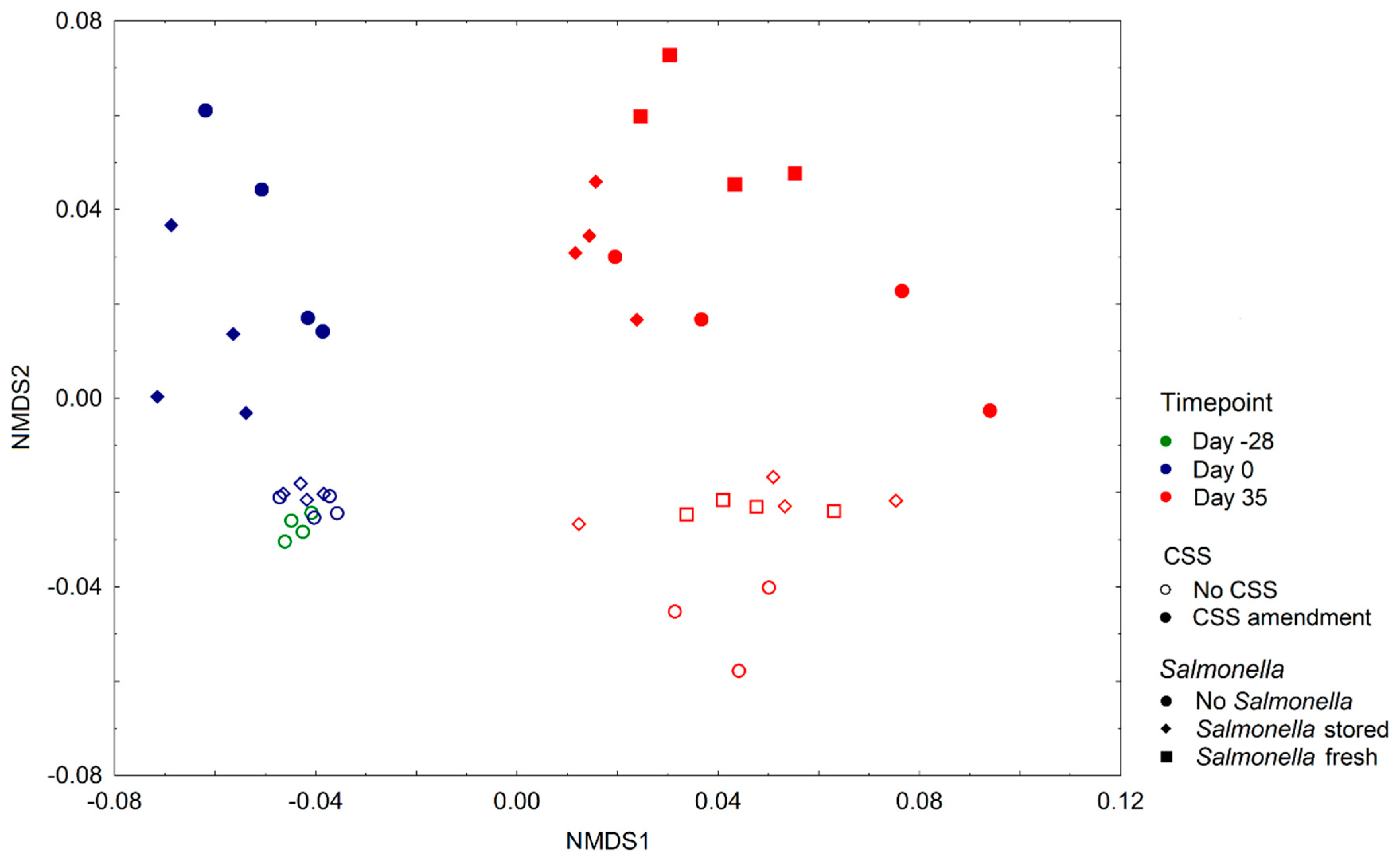
3.1. Composition of Prokaryotic Community Changes upon Time, Sludge Compost Amendment and S. enterica Presence
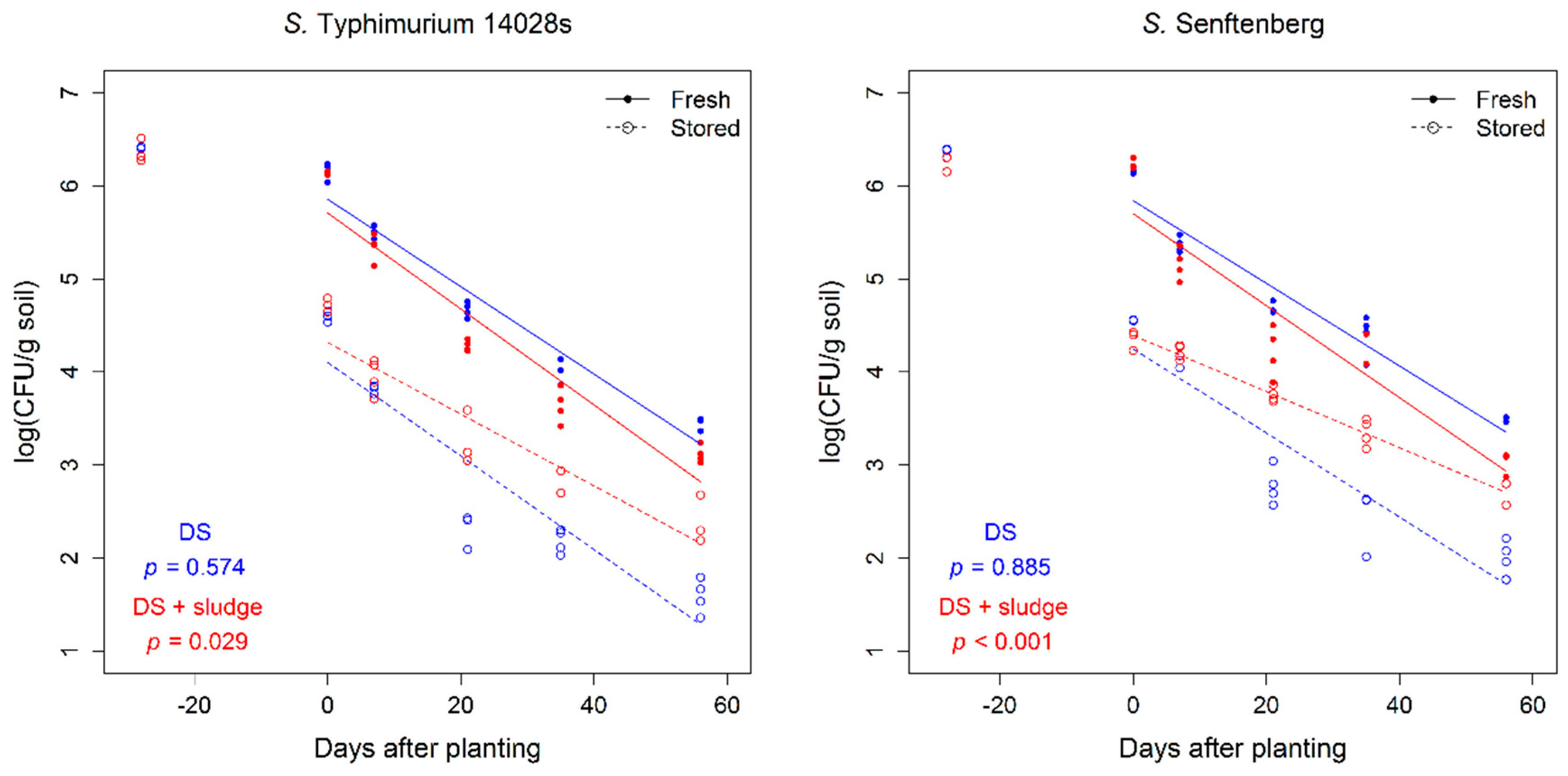
3.2. Amendment with Composted Sewage Sludge Increased the Persistence of Salmonella in Soil
3.3. Amendment with Composted Sewage Sludge Has No Impact on Colonization of B. rapa by Salmonella
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jacobsen, C.S.; Hjelmsø, M.H. Agricultural soils, pesticides and microbial diversity. Curr. Opin. Biotechnol. 2014, 27, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Jansson, J.K.; Hofmockel, K.S. The soil microbiome—From metagenomics to metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Finkel, O.M.; Castrillo, G.; Herrera Paredes, S.; Salas González, I.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Tkacz, A.; Cheema, J.; Chandra, G.; Grant, A.; Poole, P.S. Stability and succession of the rhizosphere microbiota depends upon plant type and soil composition. ISME J. 2015, 9, 2349–2359. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Steele, M.; Odumeru, J. Irrigation Water as Source of Foodborne Pathogens on Fruit and Vegetables. J. Food Prot. 2004, 67, 2839–2849. [Google Scholar] [CrossRef]
- Barak, J.D.; Liang, A.S. Role of soil, crop debris, and a plant pathogen in Salmonella enterica contamination of tomato plants. PLoS ONE 2008, 3, e1657. [Google Scholar] [CrossRef]
- Schikora, A.; Garcia, A.V.; Hirt, H. Plants as alternative hosts for Salmonella. Trends Plant Sci. 2012, 17, 245–249. [Google Scholar] [CrossRef]
- Urra, J.; Alkorta, I.; Mijangos, I.; Epelde, L.; Garbisu, C. Application of sewage sludge to agricultural soil increases the abundance of antibiotic resistance genes without altering the composition of prokaryotic communities. Sci. Total Environ. 2019, 647, 1410–1420. [Google Scholar] [CrossRef]
- Goberna, M.; Simón, P.; Hernández, M.T.; García, C. Prokaryotic communities and potential pathogens in sewage sludge: Response to wastewaster origin, loading rate and treatment technology. Sci. Total Environ. 2018, 615, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.J.; Farber, J.N.; Beuchat, L.R.; Parish, M.E.; Suslow, T.V.; Garrett, E.H.; Busta, F.F. Outbreaks Associated with Fresh Produce: Incidence, Growth, and Survival of Pathogens in Fresh and Fresh-Cut Produce. Compr. Rev. Food Sci. Food Saf. 2003, 2, 78–141. [Google Scholar] [CrossRef]
- European Commision. European Commission Council Directive 86/278/EEC of 12 June 1986 on the protection of the environment, and in particular of the soil, when sewage sludge is used in agriculture. Off. J. 1986, 181, 6–12. [Google Scholar]
- Černe, M.; Palčić, I.; Pasković, I.; Major, N.; Romić, M.; Filipović, V.; Igrc, M.D.; Perčin, A.; Goreta Ban, S.; Zorko, B.; et al. The effect of stabilization on the utilization of municipal sewage sludge as a soil amendment. Waste Manag. 2019, 94, 27–38. [Google Scholar] [CrossRef]
- Bożym, M.; Siemiątkowski, G. Characterization of composted sewage sludge during the maturation process: A pilot scale study. Environ. Sci. Pollut. Res. 2018, 25, 34332–34342. [Google Scholar] [CrossRef] [Green Version]
- Uçaroğlu, S.; Alkan, U. Composting of wastewater treatment sludge with different bulking agents. J. Air Waste Manag. Assoc. 2016, 66, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Francisco, M.; Moreno, D.A.; Cartea, M.E.; Ferreres, F.; García-Viguera, C.; Velasco, P. Simultaneous identification of glucosinolates and phenolic compounds in a representative collection of vegetable Brassica rapa. J. Chromatogr. A 2009, 1216, 6611–6619. [Google Scholar] [CrossRef]
- Jahangir, M.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Metabolomic response of Brassica rapa submitted to pre-harvest bacterial contamination. Food Chem. 2008, 107, 362–368. [Google Scholar] [CrossRef]
- Heaney, R.K.; Fenwick, G.R. Natural toxins and protective factors in brassica species, including rapeseed. Nat. Toxins 1995, 3, 233–237. [Google Scholar] [CrossRef]
- Francisco, M.; Velasco, P.; Moreno, D.A.; García-Viguera, C.; Cartea, M.E. Cooking methods of Brassica rapa affect the preservation of glucosinolates, phenolics and vitamin C. Food Res. Int. 2010, 43, 1455–1463. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.S.; Choi, H. Antidiabetic effect of Korean traditional Baechu (Chinese Cabbage) kimchi in a type 2 diabetes model of rats. J. Med. Food 2009, 12, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.; Kim, G.-H. Anticancer and Antimicrobial Activities of BETA-Phenylethyl Isothiocyanate in Brassica rapa L. Food Sci. Technol. Res. 2008, 14, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Garrett, K.A.; Dendy, S.P.; Frank, E.E.; Rouse, M.N.; Travers, S.E. Climate Change Effects on Plant Disease: Genomes to Ecosystems. Annu. Rev. Phytopathol. 2006, 44, 489–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alegbeleye, O.O.; Singleton, I.; Sant’Ana, A.S. Sources and contamination routes of microbial pathogens to fresh produce during field cultivation: A review. Food Microbiol. 2018, 73, 177–208. [Google Scholar] [CrossRef]
- Madden, J.M. Microbial pathogens in fresh producethe regulatory perspective. J. Food Prot. 1992, 55, 821–823. [Google Scholar] [CrossRef]
- Lynch, M.F.; Tauxe, R.V.; Hedberg, C.W. The growing burden of foodborne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307–315. [Google Scholar] [CrossRef]
- Deering, A.J.; Mauer, L.J.; Pruitt, R.E. Internalization of E. coli O157:H7 and Salmonella spp. in plants: A review. Food Res. Int. 2012, 45, 567–575. [Google Scholar] [CrossRef]
- Schierstaedt, J.; Jechalke, S.; Nesme, J.; Neuhaus, K.; Sørensen, S.J.; Grosch, R.; Smalla, K.; Schikora, A. Salmonella persistence in soil depends on reciprocal interactions with indigenous microorganisms. Environ. Microbiol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Jechalke, S.; Schierstaedt, J.; Becker, M.; Flemer, B.; Grosch, R.; Smalla, K.; Schikora, A. Salmonella Establishment in Agricultural Soil and Colonization of Crop Plants Depend on Soil Type and Plant Species. Front. Microbiol. 2019, 10, 967. [Google Scholar] [CrossRef] [Green Version]
- Canet, R.; Pomares, F.; Cabot, B.; Chaves, C.; Ferrer, E.; Ribó, M.; Albiach, M.R. Composting olive mill pomace and other residues from rural southeastern Spain. Waste Manag. 2008, 28, 2585–2592. [Google Scholar] [CrossRef]
- US Environmental Protection Agency. EPA Method 1684: Total, Fixed, and Volatile Solids in Water, Solid, and Biosolids; US Environtmental Protection Agency: Washington, DC, USA, 2001.
- Rühlmann, J.; Ruppel, S. Effects of organic amendments on soil carbon content and microbial biomass—Results of the long-term box plot experiment in Grossbeeren. Arch. Agron. Soil Sci. 2005, 51, 163–170. [Google Scholar] [CrossRef]
- Schreiter, S.; Ding, G.-C.; Heuer, H.; Neumann, G.; Sandmann, M.; Grosch, R.; Kropf, S.; Smalla, K. Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce. Front. Microbiol. 2014, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- European Commission. European Commision Council Directive of 12 December 1991 concerning the protection of waters against pollution caused by nitrates from agricultural sources. Off. J. 1991, 375, 1–8. [Google Scholar]
- Yu, Y.; Lee, C.; Kim, J.; Hwang, S. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol. Bioeng. 2005, 89, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, C.; Al-Soud, W.A.; Larsson, M.; Alm, E.; Yekta, S.S.; Svensson, B.H.; Sørensen, S.J.; Karlsson, A. 454 pyrosequencing analyses of bacterial and archaeal richness in 21 full-scale biogas digesters. FEMS Microbiol. Ecol. 2013, 85, 612–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, d633–d642. [Google Scholar] [CrossRef] [Green Version]
- R Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 9 July 2020).
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Spain, A.M.; Krumholz, L.R.; Elshahed, M.S. Abundance, composition, diversity and novelty of soil Proteobacteria. ISME J. 2009, 3, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Schrempf, H. Actinobacteria within soils: Capacities for mutualism, symbiosis and pathogenesis. FEMS Microbiol. Lett. 2013, 342, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathya, A.; Vijayabharathi, R.; Gopalakrishnan, S. Plant growth-promoting actinobacteria: A new strategy for enhancing sustainable production and protection of grain legumes. 3 Biotech 2017, 7, 102. [Google Scholar] [CrossRef] [Green Version]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A.; Błaszczyk, M. Bacteroidetes as a sensitive biological indicator of agricultural soil usage revealed by a culture-independent approach. Appl. Soil Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Yao, L.; Wang, D.; Kang, L.; Wang, D.; Zhang, Y.; Hou, X.; Guo, Y. Effects of fertilizations on soil bacteria and fungi communities in a degraded arid steppe revealed by high through-put sequencing. PeerJ 2018, 6, e4623. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Q.; Han, X. Soil Bacterial Communities Respond to Mowing and Nutrient Addition in a Steppe Ecosystem. PLoS ONE 2013, 8, e84210. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Loria, R. Virulence mechanisms of Gram-Positive plant pathogenic bacteria. Curr. Opin. Plant Biol. 2008, 11, 449–456. [Google Scholar] [CrossRef]
- Su, J.-Q.; Wei, B.; Ou-Yang, W.-Y.; Huang, F.-Y.; Zhao, Y.; Xu, H.-J.; Zhu, Y.-G. Antibiotic Resistome and Its Association with Bacterial Communities during Sewage Sludge Composting. Environ. Sci. Technol. 2015, 49, 7356–7363. [Google Scholar] [CrossRef]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The Ecology of Acidobacteria: Moving beyond Genes and Genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, J.; et al. Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kant, S. Understanding nitrate uptake, signaling and remobilisation for improving plant nitrogen use efficiency. Semin. Cell Dev. Biol. 2018, 74, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Fornefeld, E.; Baklawa, M.; Hallmann, J.; Schikora, A.; Smalla, K. Sewage sludge amendment and inoculation with plant-parasitic nematodes do not facilitate the internalization of Salmonella Typhimurium LT2 in lettuce plants. Food Microbiol. 2018, 71, 111–119. [Google Scholar] [CrossRef]
- Shah, M.K.; Bradshaw, R.; Nyarko, E.; Handy, E.T.; East, C.; Millner, P.D.; Bergholz, T.M.; Sharma, M. Salmonella enterica in soils amended with heat-treated poultry pellets survived longer than bacteria in unamended soils and more readily transferred to and persisted on spinach. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Koukkidis, G.; Haigh, R.; Allcock, N.; Jordan, S.; Freestone, P. Salad leaf juices enhance Salmonella growth, colonization of fresh produce, and virulence. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Weese, J.S.; Flemming, C.; Odumeru, J.; Warriner, K. Fate of Clostridium difficile during wastewater treatment and incidence in Southern Ontario watersheds. J. Appl. Microbiol. 2014, 117, 891–904. [Google Scholar] [CrossRef]
- Chen, Q.; An, X.; Li, H.; Su, J.; Ma, Y.; Zhu, Y.G. Long-term field application of sewage sludge increases the abundance of antibiotic resistance genes in soil. Environ. Int. 2016, 92–93, 1–10. [Google Scholar] [CrossRef]
- Zhang, J.; Sui, Q.; Tong, J.; Zhong, H.; Wang, Y.; Chen, M.; Wei, Y. Soil types influence the fate of antibiotic-resistant bacteria and antibiotic resistance genes following the land application of sludge composts. Environ. Int. 2018, 118, 34–43. [Google Scholar] [CrossRef]
- Bondarczuk, K.; Markowicz, A.; Piotrowska-Seget, Z. The urgent need for risk assessment on the antibiotic resistance spread via sewage sludge land application. Environ. Int. 2016, 87, 49–55. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Sludge | Salmonella | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Planting Day n = 16 | 35 dpi n = 23 | p-Value | Control n = 23 | Sludge n = 20 | p-Value | Control n = 19 | 14028s n = 24 | p-Value | |
| Proteobacteria | 20.1 ± 1.5 | 30.5 ± 3.8 | ˂0.001 | 26.3 ± 6.2 | 25.0 ± 5.9 | 0.946 | 24.1 ± 6.0 | 26.9 ± 5.9 | 0.731 |
| Actinobacteria | 18.8 ± 1.3 | 20.3 ± 1.8 | 0.003 | 19.6 ± 2.1 | 19.7 ± 1.2 | 0.918 | 19.6 ± 2.0 | 19.7 ± 1.5 | 0.606 |
| Firmicutes | 20.7 ± 2.7 | 13.2 ± 3.3 | ˂0.001 | 15.6 ± 4.0 | 17.9 ± 5.4 | ˂0.001 | 17.9 ± 5.4 | 15.8 ± 4.2 | 0.924 |
| Acidobacteria | 16.3 ± 4.0 | 14.8 ± 1.0 | ˂0.001 | 16.1 ± 1.5 | 14.8 ± 1.0 | ˂0.001 | 15.6 ± 1.3 | 15.4 ± 1.6 | 0.909 |
| Crenarchaeota | 10.9 ± 1.2 | 6.2 ± 1.4 | ˂0.001 | 8.6 ± 2.9 | 8.2 ± 2.5 | 0.869 | 8.9 ± 2.8 | 8.0 ± 2.6 | 0.452 |
| Bacteroidetes | 5.8 ± 0.7 | 7.6 ± 1.3 | ˂0.001 | 6.7 ± 1.1 | 6.9 ± 1.7 | 0.848 | 6.5 ± 1.0 | 7.0 ± 1.7 | 0.919 |
| Gemmatimonadetes | 2.3 ± 0.3 | 2.3 ± 0.2 | 0.507 | 2.3 ± 0.2 | 2.3 ± 0.3 | 1.000 | 2.3 ± 0.3 | 2.4 ± 0.2 | 0.522 |
| Chloroflexi | 1.6 ± 0.3 | 1.8 ± 0.3 | 0.035 | 1.6 ± 0.2 | 1.9 ± 0.3 | ˂0.001 | 1.7 ± 0.3 | 1.7 ± 0.3 | 0.517 |
| Nitrospirae | 1.3 ± 0.1 | 0.9 ± 0.2 | ˂0.001 | 1.1 ± 0.3 | 1.1 ± 0.2 | 0.961 | 1.2 ± 0.2 | 1.1 ± 0.3 | 0.566 |
| TM7 | 1.0 ± 0.2 | 0.9 ± 0.3 | 0.729 | 1.0 ± 0.2 | 0.9 ± 0.3 | 1.000 | 1.0 ± 0.2 | 0.9 ± 0.2 | 0.481 |
| Verrucomicrobia | 0.8 ± 0.1 | 1.1 ± 0.1 | ˂0.001 | 0.9 ± 0.2 | 1.0 ± 0.2 | 0.888 | 0.9 ± 0.2 | 1.0 ± 0.1 | 0.616 |
| Soil | Serovar | Scenario | Days after Planting | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DS soil | DS soil + CSS | 14028s | Senftenberg | Fresh | Stored | 7 | 21 | 35 | 56 | ||
| Min (%) | 1.9 | 1.9 | 1.9 | 1.9 | 2.8 | 0.9 | 1.9 | 5.6 | 0.0 | 0.0 | 1.9 |
| Max (%) | 3.8 | 3.8 | 3.8 | 3.8 | 5.6 | 1.9 | 3.8 | 11.3 | 0.0 | 0.0 | 3.8 |
| p-value | 1.000 | 1.000 | 0.066 | 0.001 | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Major, N.; Schierstaedt, J.; Jechalke, S.; Nesme, J.; Ban, S.G.; Černe, M.; Sørensen, S.J.; Ban, D.; Schikora, A. Composted Sewage Sludge Influences the Microbiome and Persistence of Human Pathogens in Soil. Microorganisms 2020, 8, 1020. https://doi.org/10.3390/microorganisms8071020
Major N, Schierstaedt J, Jechalke S, Nesme J, Ban SG, Černe M, Sørensen SJ, Ban D, Schikora A. Composted Sewage Sludge Influences the Microbiome and Persistence of Human Pathogens in Soil. Microorganisms. 2020; 8(7):1020. https://doi.org/10.3390/microorganisms8071020
Chicago/Turabian StyleMajor, Nikola, Jasper Schierstaedt, Sven Jechalke, Joseph Nesme, Smiljana Goreta Ban, Marko Černe, Søren J. Sørensen, Dean Ban, and Adam Schikora. 2020. "Composted Sewage Sludge Influences the Microbiome and Persistence of Human Pathogens in Soil" Microorganisms 8, no. 7: 1020. https://doi.org/10.3390/microorganisms8071020
APA StyleMajor, N., Schierstaedt, J., Jechalke, S., Nesme, J., Ban, S. G., Černe, M., Sørensen, S. J., Ban, D., & Schikora, A. (2020). Composted Sewage Sludge Influences the Microbiome and Persistence of Human Pathogens in Soil. Microorganisms, 8(7), 1020. https://doi.org/10.3390/microorganisms8071020