
Catalysis of Chlorovirus Production by the Foraging of Bursaria truncatella on Paramecia bursaria Containing Endosymbiotic Algae

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
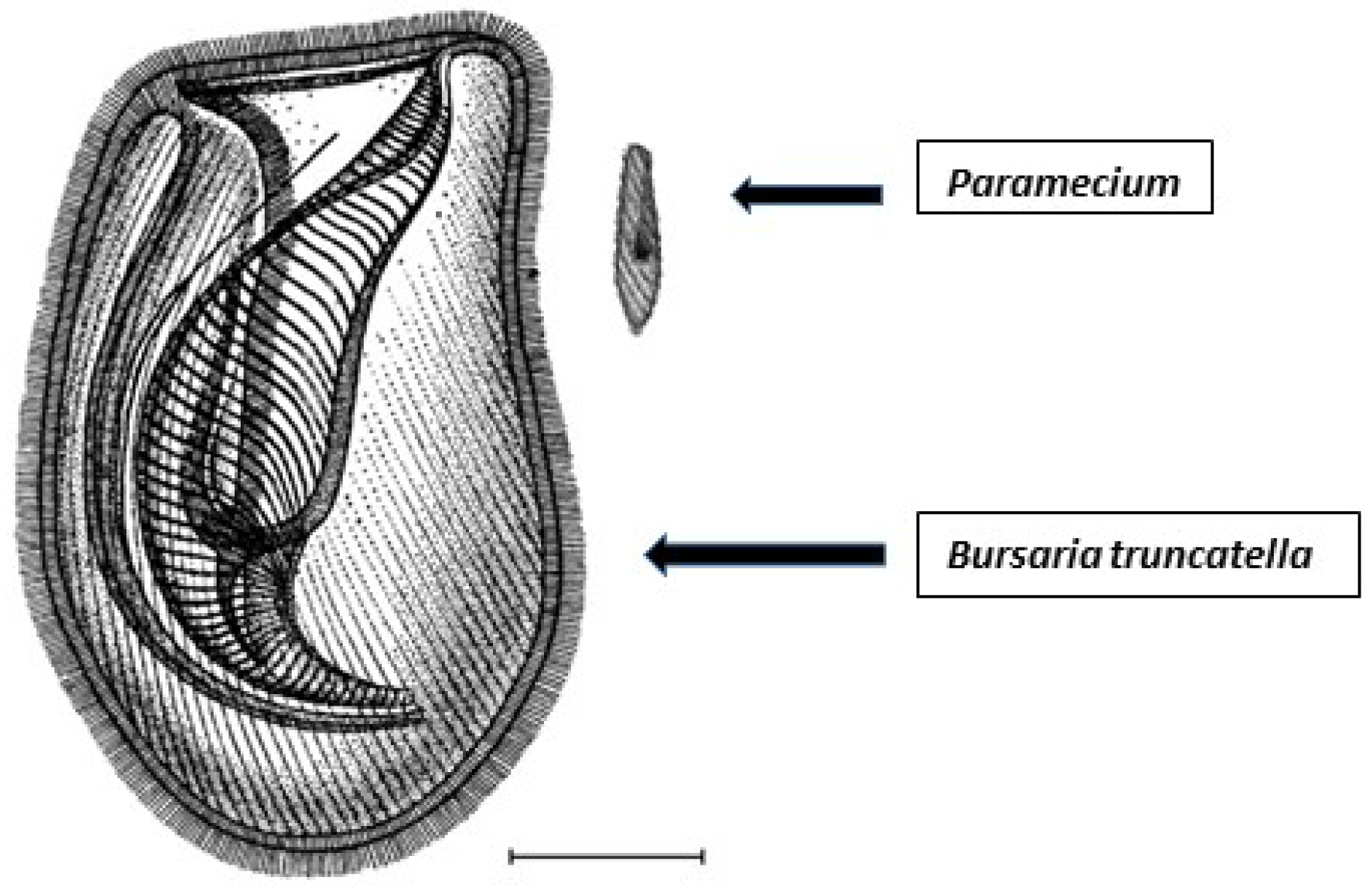
2.1. Organisms and Culturing
2.2. Long-Term Foraging Experiments
2.3. Short-Term Foraging Experiments
2.4. Monitoring Chloroviruses Inside B. truncatella
3. Results
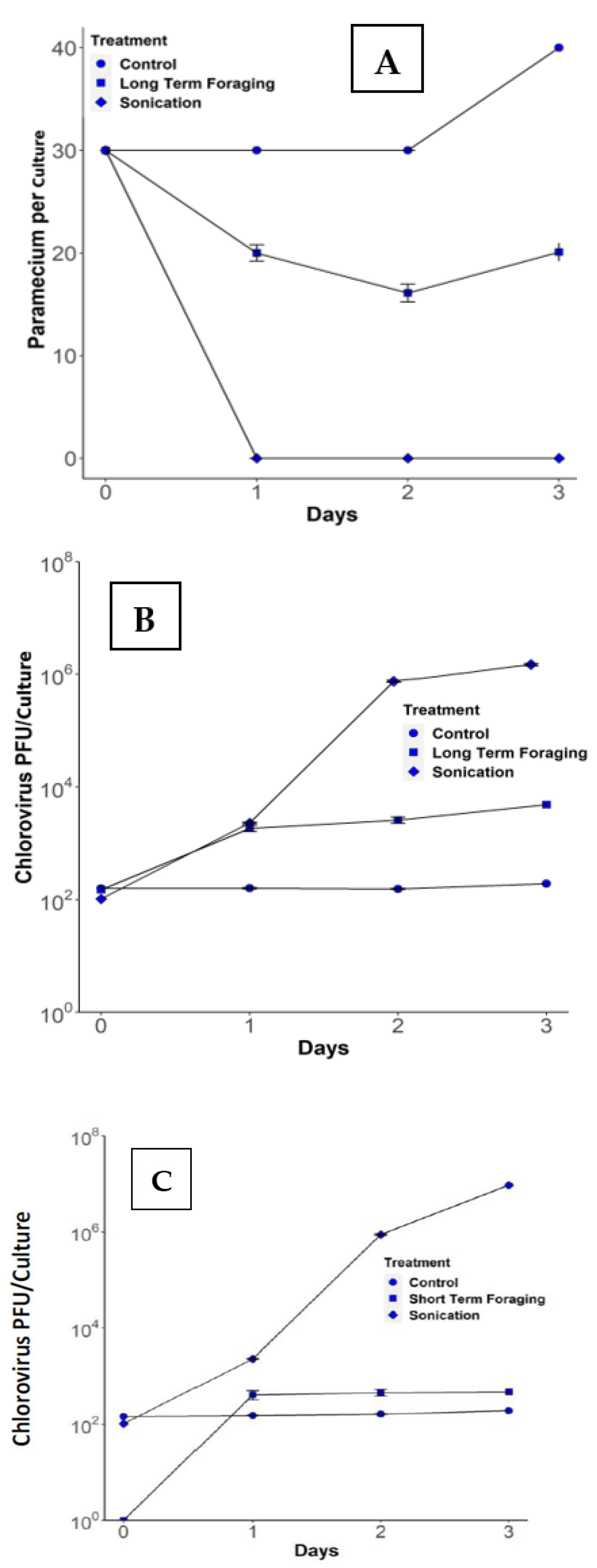
3.1. Long-Term Foraging Experiments
3.2. Short-Term Foraging Experiments
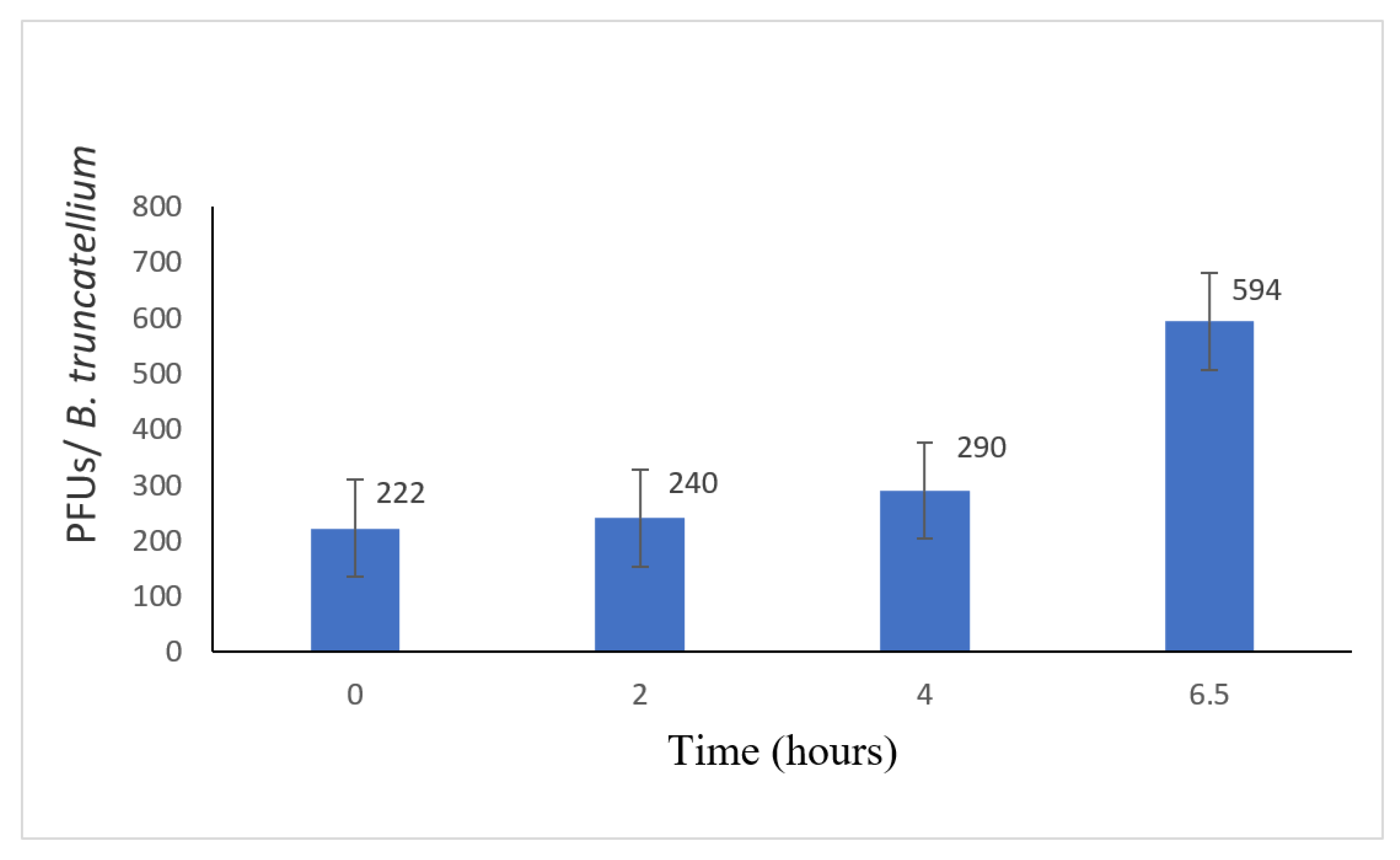
3.3. Disruption of the B. truncatella
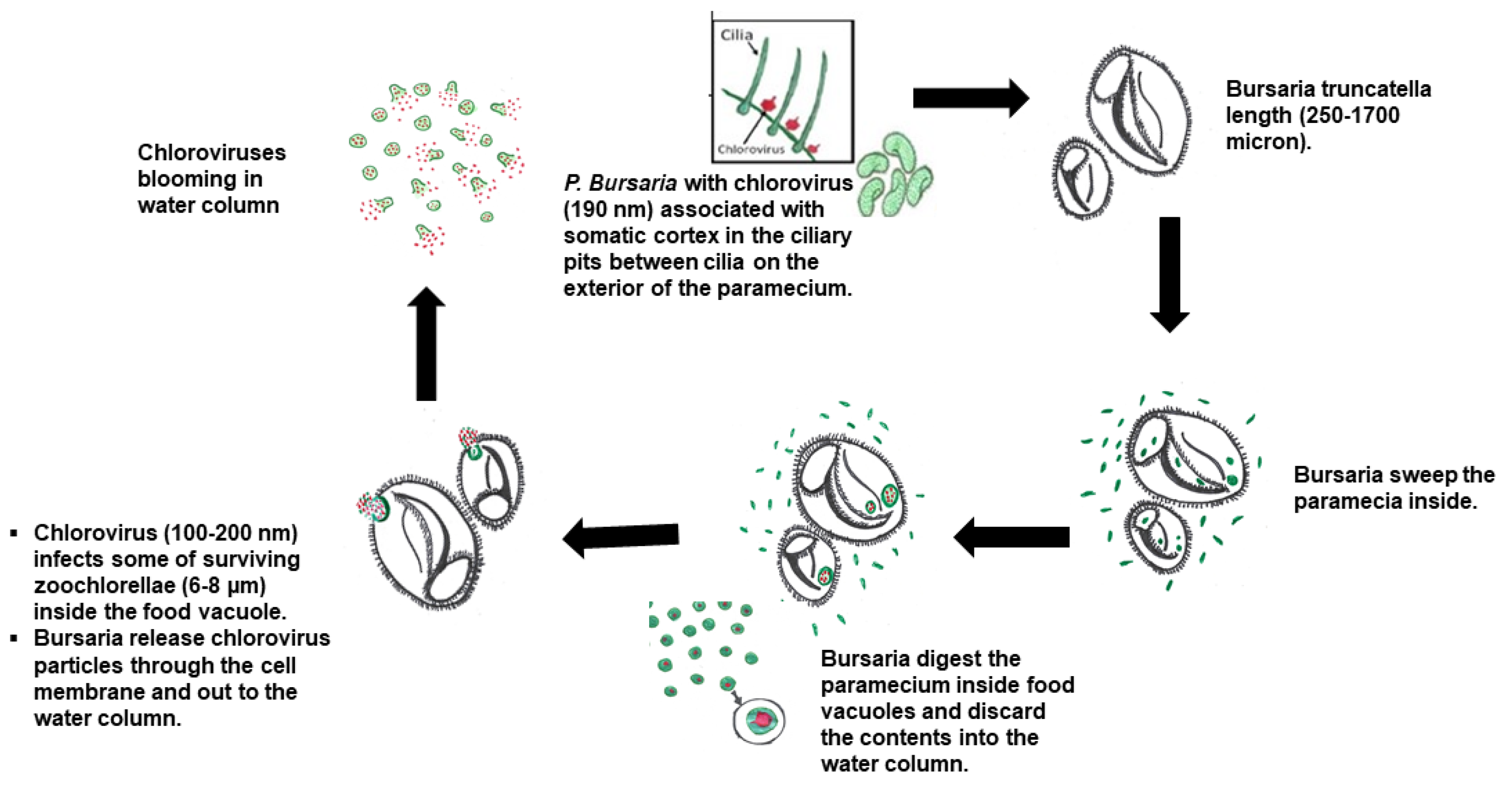
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuhrman, J.A. Marine viruses and their biogeochemical and ecological effects. Nature 1999, 399, 541–548. [Google Scholar] [CrossRef]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef]
- Bergh, O.; Børsheim, K.Y.; Bratbak, G.; Heldal, M. High abundance of viruses found in aquatic environments. Nature 1989, 340, 467–468. [Google Scholar] [CrossRef]
- Suttle, C.A. Marine viruses—Major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef]
- Middelbe, M.; Brussaard, C. Marine viruses: Key players in marine ecosystems. Viruses 2017, 9, 302. [Google Scholar] [CrossRef] [Green Version]
- Karakashian, S.J.; Karakashian, M.W. Evolution and symbiosis in the genus Chlorella and related algae. Evolution 1965, 19, 368–377. [Google Scholar] [CrossRef]
- Kodama, Y.; Suzuki, H.; Dohra, H.; Sugii, M.; Kitazume, T.; Yamaguchi, K.; Shigenobu, S.; Fujishima, M. Comparison of gene expression of Paramecium bursaria with and without Chlorella variabilis symbionts. BMC Genom. 2014, 15, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Etten, J.L.; Dunigan, D.D. Chloroviruses: Not your everyday plant virus. Trends Plant Sci. 2012, 17, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Van Etten, J.L.; Dunigan, D.D. Giant chloroviruses: Five easy questions. PLoS Pathog. 2016, 12, e1005751. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, J.L.; Agarkova, I.V.; Dunigan, D.D. Chloroviruses. Viruses 2020, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Van Etten, J.L.; van Etten, C.H.; Johnson, J.K.; Burbank, D.E. A survey for viruses from fresh water that infect a eukaryotic chlorella-like green alga. Appl. Environ. Microbiol. 1985, 49, 1326–1328. [Google Scholar] [CrossRef] [Green Version]
- Quispe, C.F.; Sonderman, O.; Seng, A.; Rasmussen, B.; Weber, G.; Mueller, C.; Dunigan, D.D.; van Etten, J.L. Three-year survey of abundance, prevalence and genetic diversity of chlorovirus population in a small urban lake. Arch. Virol. 2016, 161, 1839–1847. [Google Scholar] [CrossRef] [Green Version]
- Sommaruga, R.; Sonntag, B. Photobiological aspects of the mutualistic association between Paramecium bursaria and Chlorella Endosymbionts in Paramecium. Microbiol. Monogr. 2009, 12, 111–130. [Google Scholar] [CrossRef]
- Tonooka, Y.; Watanabe, T. A natural strain of Paramecium bursaria lacking symbiotic algae. Eur. J. Protistol. 2002, 38, 55–58. [Google Scholar] [CrossRef]
- Kadono, T.; Kawano, T.; Hosoya, H.; Kosaka, T. Flow cytometric studies of the host-regulated cell cycle in algae symbiotic with green paramecium. Protoplasma 2004, 223, 133–141. [Google Scholar] [CrossRef]
- Kadono, T.; Kawano, T. Natural historical views on the controlled cell growth and oxidative stress responses in symbiotic associations between ciliated protozoa and green algae in green paramecia. New Technol. Med. 2007, 8, 439–445. [Google Scholar]
- Kawakami, H.; Kawakami, N. Behavior of a virus in a symbiotic system Paramecium bursaria-zoochlorella. J. Protozool. 1978, 25, 217–225. [Google Scholar] [CrossRef]
- Yashchenko, V.V.; Gavrilova, O.V.; Rautian, M.S.; Jakobsen, K.S. Association of Paramecium bursaria chlorella viruses with Paramecium bursaria cells: Ultrastructural studies. Eur. J. Protistol. 2012, 48, 149–159. [Google Scholar] [CrossRef]
- Mitra, A.; Flynn, K.J.; Tillmann, U.; Raven, J.A.; Caron, D.; Stoecker, D.K.; Not, F.; Hansen, P.J.; Hallegraeff, G.; Sanders, R.; et al. Defining planktonic protist functional groups on mechanisms for energy and nutrient acquisition: Incorporation of diverse mixotrophic strategies. Protist 2016, 167, 106–120. [Google Scholar] [CrossRef]
- DeLong, J.P.; Al-Ameeli, Z.T.; Duncan, G.; van Etten, J.L.; Dunigan, D.D. Predators catalyze an increase in chloroviruses by foraging on the symbiotic hosts of zoochlorellae. Proc. Natl. Acad. Sci. USA 2016, 113, 13780–13784. [Google Scholar] [CrossRef] [Green Version]
- DeLong, J.P.; Al-Ameeli, Z.T.; Lyon, S.; van Etten, J.L.; Dunigan, D.D. Size-dependent catalysis of Chlorovirus population growth by a messy feeding predator. Microbial. Ecol. 2016, 75, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Beers, C.D. Excystment in the ciliate Bursaria truncatella. Biol. Bull. 1948, 94, 86–98. [Google Scholar] [CrossRef]
- Bary, B.M. Studies on the freshwater ciliates of New Zealand. Part 1: A general morphology of Bursaria truncatella Muller. Trans. Proc. R. Soc. N. Z. 1950, 78, 301–310. [Google Scholar]
- Krause, R.; Bräucker, M. Gravitaxis of Bursaria truncatella: Electrophysiological and behavioural analyses of a large ciliate cell. Eur. J. Protistol. 2009, 45, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Novich, R.A.; Erickson, E.K.; Kalinoski, R.M.; DeLong, J.P. The temperature independence of interaction strength in a sit-and-wait predator. Ecosphere 2014, 5, 1–9. [Google Scholar] [CrossRef]
- Van Etten, J.L.; Burbank, D.E.; Kuczmarski, D.; Meints, R.H. Virus infection of culturable chlorella-like algae and development of a plaque assay. Science 1983, 219, 994–996. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, J.L.; Burbank, D.E.; Xia, Y.; Meints, R.H. Growth cycle of a virus, PBCV-1, that infects Chlorella-like algae. Virology 1983, 126, 117–125. [Google Scholar] [CrossRef]
- Foissner, W.; Chao, A.; Katz, L.A. Diversity and geographic distribution of ciliates (Protista: Ciliophora). Biodivers. Conserv. 2008, 17, 345–363. [Google Scholar] [CrossRef]
- Hechinger, R.F.; Lafferty, K.D.; Dobson, A.P.; Brown, J.H.; Kuris, A.M. A common scaling rule for abundance, energetics, and production of parasitic and free-living species. Science 2011, 333, 445–448. [Google Scholar] [CrossRef] [Green Version]
- Caceres, C.E.; Knight, C.J.; Hakk, S.R. Predator-spreaders: Predation can enhance parasite success in a planktonic host-parasite system. Ecology 2009, 90, 2850–2858. [Google Scholar] [CrossRef] [Green Version]
- Frada, M.J.; Schatz, D.; Farstey, V.; Ossiolinski, J.E.; Sabanay, H.; Ben-Dor, S.; Koren, I.; Vardi, A. Zooplankton may serve as transmission vectors for viruses infecting algal blooms in the ocean. Curr. Biol. 2014, 24, 2592–2597. [Google Scholar] [CrossRef] [Green Version]
- Vermont, A.I.; Martinez, M.J.; Waller, J.D.; Gilg, I.C.; Leavitt, A.H.; Floge, S.A.; Archer, S.D.; Wilson, W.H.; Fields, D.M. Virus infection of Emiliania huxleyideters grazing by the copepod Acartia tonsa. J. Plankton Res. 2016, 38, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Berger, J. Feeding behavior of Didinium nasutum on Paramecium bursaria with normal or apochlorotic zoochlorellae. J. Gen. Microbiol. 1980, 118, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Bevington, D.J.; White, C.; Wallace, R.L. Predatory behavior of Cupelopagis vorax (Rotifera; Collothecacea; Atrochidae) on protozoan prey. Hydrobiologia 1995, 313, 213–217. [Google Scholar] [CrossRef]
- DeLong, J.P. Experimental demonstration of a ‘rate-size’ trade-off governing body size optimization. Evol. Ecol. Res. 2012, 14, 343–352. [Google Scholar]
- Weitz, J.S.; Stock, C.A.; Wilhelm, S.W.; Bourouiba, L.; Coleman, M.L.; Buchan, A.; Follows, M.J.; Fuhrman, J.A.; Jover, L.F.; Lennon, J.T.; et al. A multitrophic model to quantify the effects of marine viruses on microbial food webs and ecosystem processes. ISME J. 2015, 9, 1352–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Ameeli, Z.T.; Al-Sammak, M.A.; DeLong, J.P.; Dunigan, D.D.; Van Etten, J.L. Catalysis of Chlorovirus Production by the Foraging of Bursaria truncatella on Paramecia bursaria Containing Endosymbiotic Algae. Microorganisms 2021, 9, 2170. https://doi.org/10.3390/microorganisms9102170
Al-Ameeli ZT, Al-Sammak MA, DeLong JP, Dunigan DD, Van Etten JL. Catalysis of Chlorovirus Production by the Foraging of Bursaria truncatella on Paramecia bursaria Containing Endosymbiotic Algae. Microorganisms. 2021; 9(10):2170. https://doi.org/10.3390/microorganisms9102170
Chicago/Turabian StyleAl-Ameeli, Zeina T., Maitham A. Al-Sammak, John P. DeLong, David D. Dunigan, and James L. Van Etten. 2021. "Catalysis of Chlorovirus Production by the Foraging of Bursaria truncatella on Paramecia bursaria Containing Endosymbiotic Algae" Microorganisms 9, no. 10: 2170. https://doi.org/10.3390/microorganisms9102170
APA StyleAl-Ameeli, Z. T., Al-Sammak, M. A., DeLong, J. P., Dunigan, D. D., & Van Etten, J. L. (2021). Catalysis of Chlorovirus Production by the Foraging of Bursaria truncatella on Paramecia bursaria Containing Endosymbiotic Algae. Microorganisms, 9(10), 2170. https://doi.org/10.3390/microorganisms9102170