
Immunostimulatory Activity of Synbiotics Using Lactococcus lactis SG-030 and Glucooligosaccharides from Weissella cibaria YRK005


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Lactic Acid Bacteria Strains and Growth Conditions
2.2. Determination of Antioxidant Activity of Lactic Acid Bacteria
Asample: Absorbance of sample
Acontrol: Absorbance of control
2.3. Determination of Acid Tolerance of Lactic Acid Bacteria
2.4. Determination of Bile Salt Tolerance of Lactic Acid Bacteria
2.5. Adhesion Ability to Intestinal Cells
2.6. Identification of Lactic Acid Bacteria
2.7. Production and Purification of Glucooligosaccharides from Weissella cibaria YRK005
2.8. Immunostimulatory Activity of Synbiotics
2.8.1. Application of Synbiotics to RAW 264.7 Cells
2.8.2. Measurement of Nitric Oxide Production
2.8.3. cDNA Synthesis and Quantitative Real-Time PCR
2.8.4. Western Blotting
2.8.5. Statistical Analysis
3. Results and Discussion
3.1. Selection of Lactic Acid Bacteria Which Have Probiotic Properties
3.2. Identification of Lactic Acid Bacteria
3.3. Cytotoxicity and Immunostimulatory Activity of L. lactis SG-030 and Glucooligosaccharides from Weissella cibaria YRK005
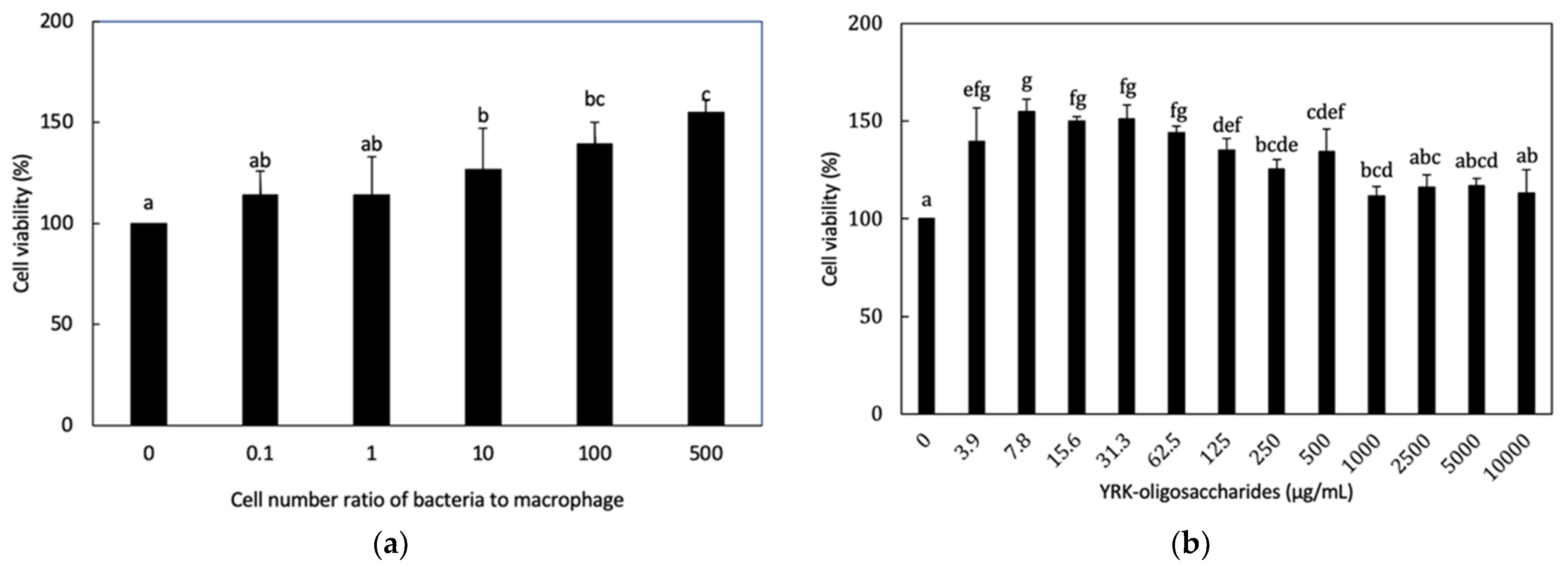
3.3.1. Cytotoxicity of L. lactis SG-030 and Glucooligosaccharides from Weissella cibaria YRK005 to RAW 264.7 Cells
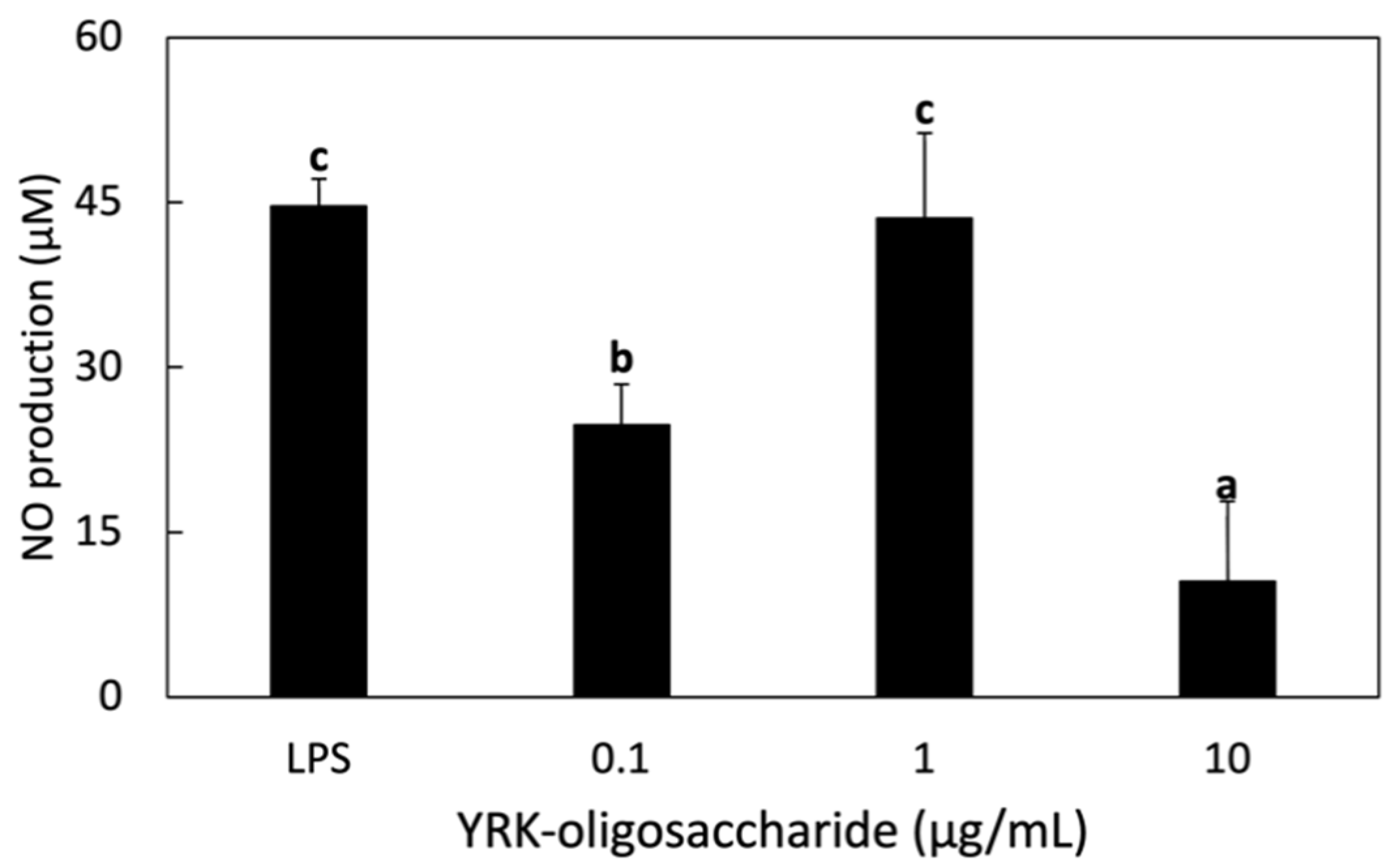
3.3.2. NO Production in RAW 264.7 Cells Treated with Glucooligosaccharides
3.4. Immunostimulatory Activity of Synbiotics
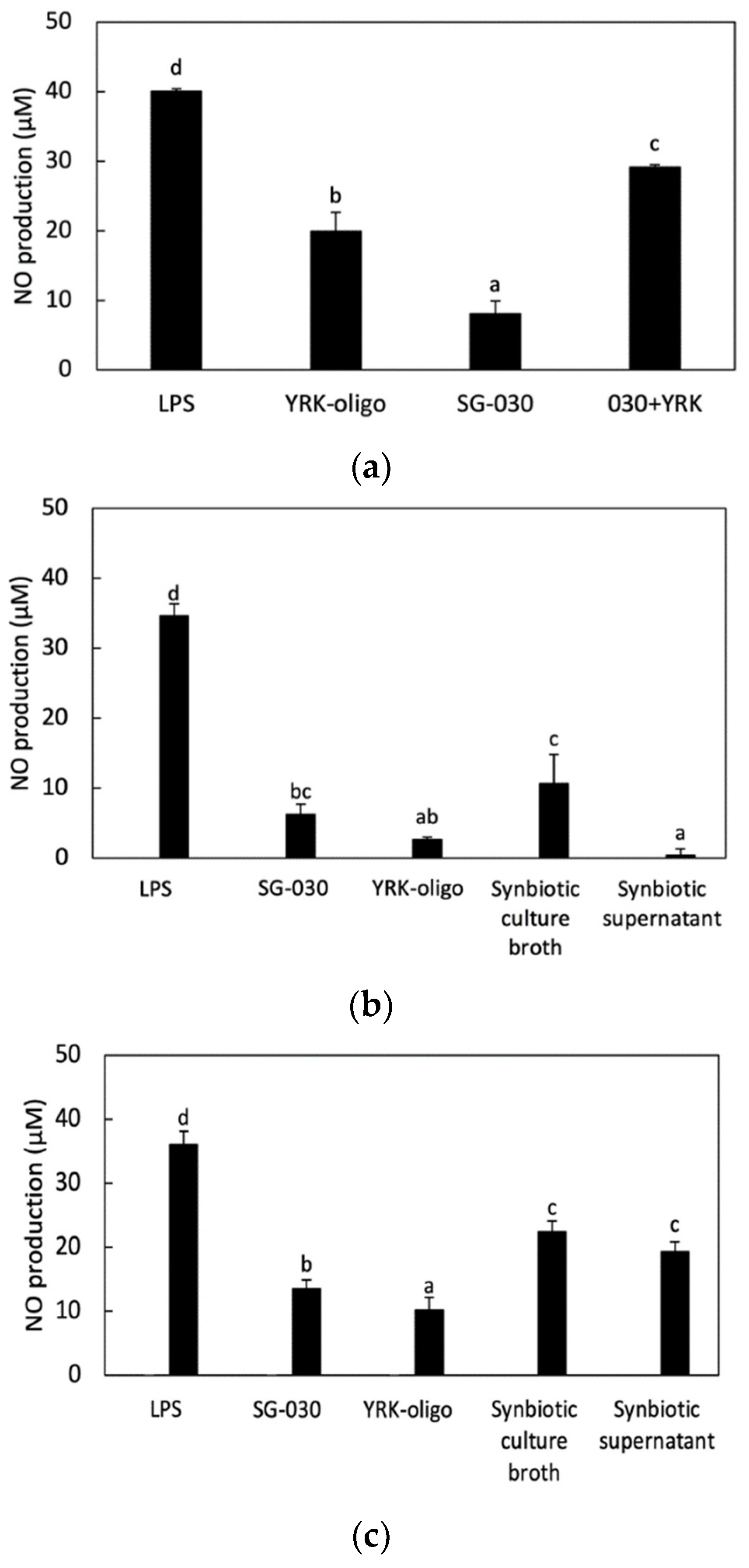
3.4.1. Effect of Synbiotics Treatment on NO Production in RAW 264.7 Cells
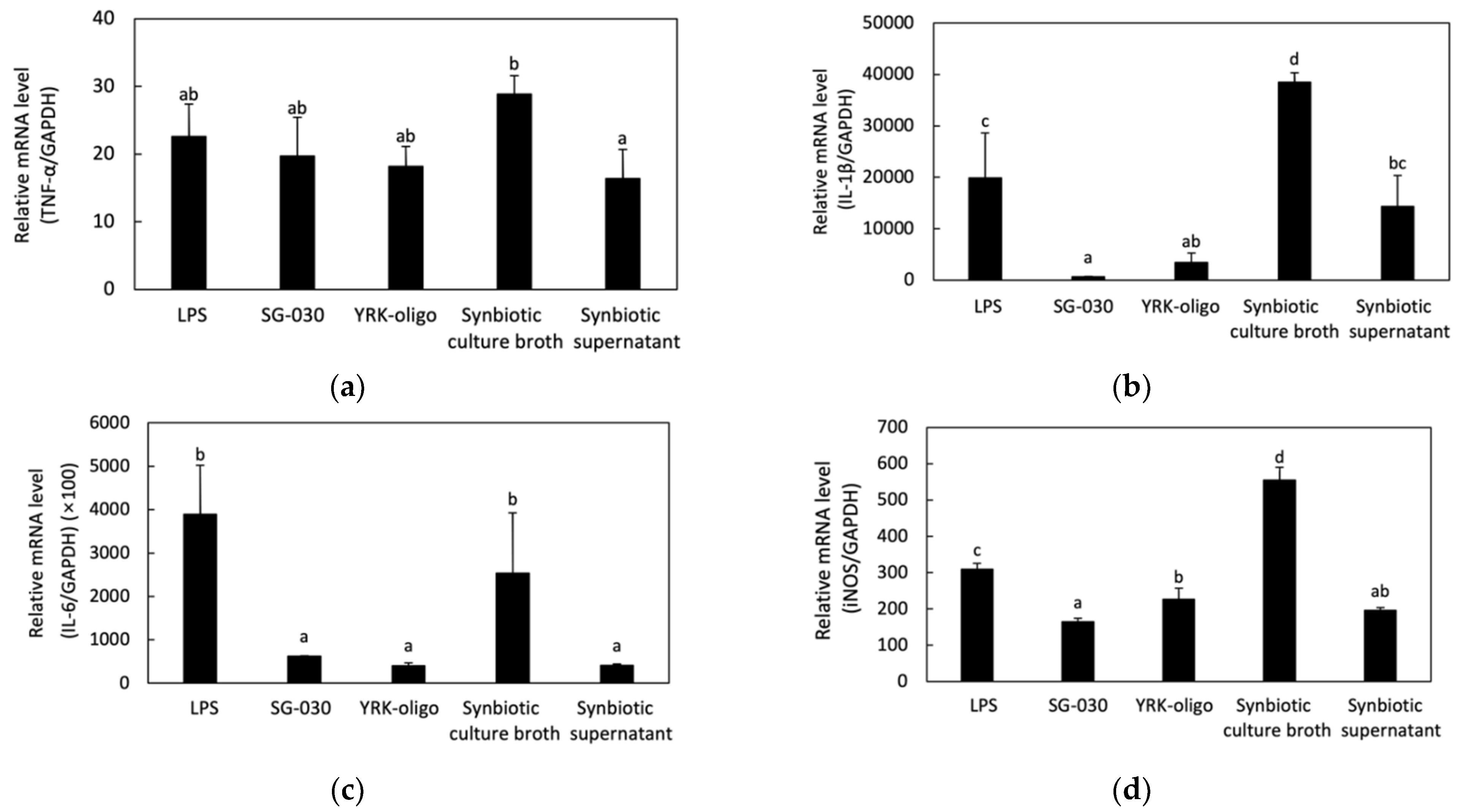
3.4.2. Effect of Synbiotics on Cytokines and INOS Secretions in RAW 264.7 Cells
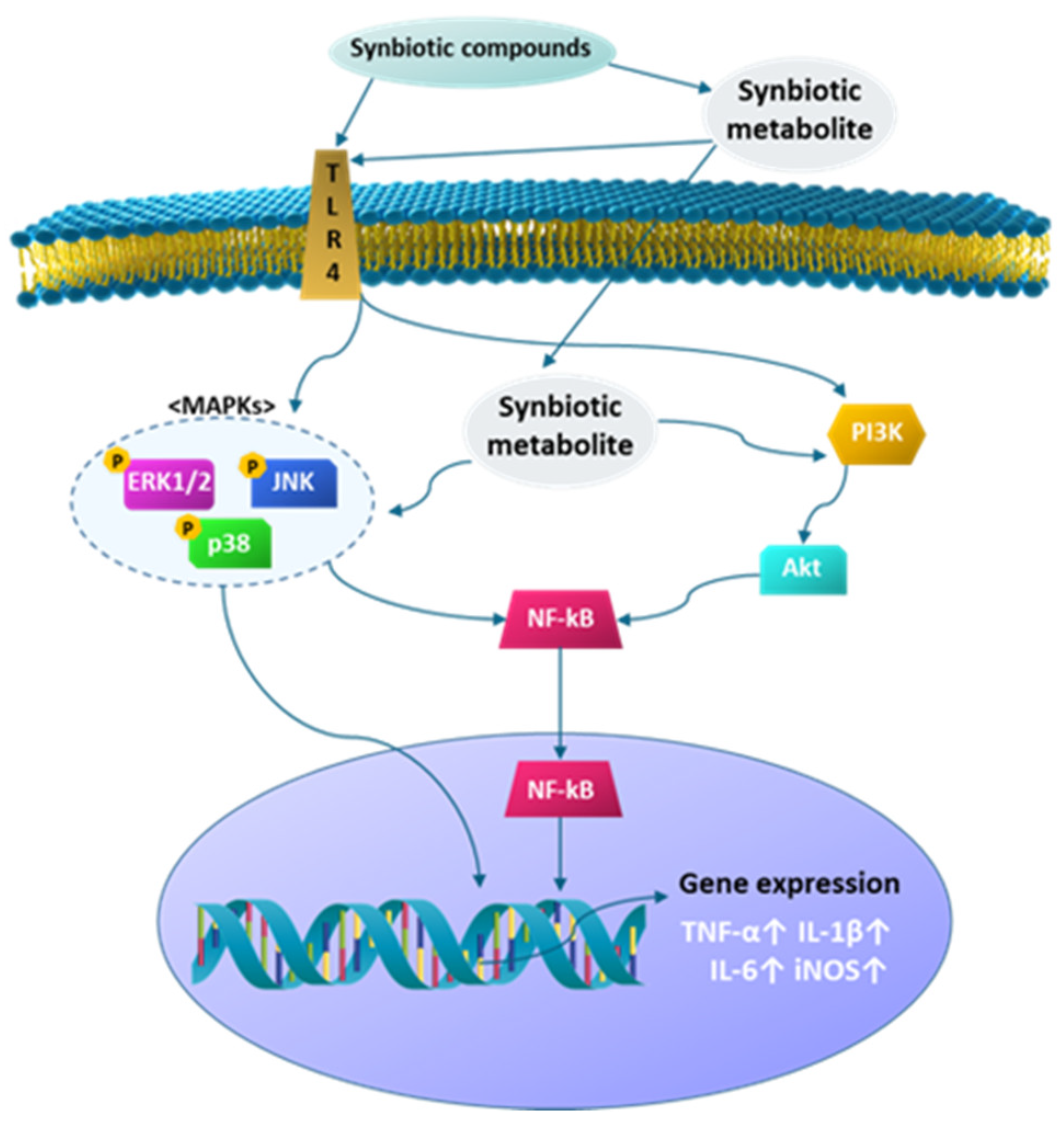
3.4.3. Effect of Synbiotics on the MAPK and PI3K Signaling Pathway in RAW 264.7 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO/WHO. Report of a Joint FAO/WHO Expert Consultation on Evaluation of Health And Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; World Health Organization and Food and Agriculture Organization of the United Nations: London, ON, Canada, 2001. [Google Scholar]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiol. Lett. 2012, 334, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, N.B.; Bryrup, T.; Allin, K.H.; Nielsen, T.; Hansen, T.H.; Pedersen, O. Alterations in fecal microbiota composition by probiotic supplementation in healthy adults: A systematic review of randomized controlled trials. Genome Med. 2016, 8, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, C.P.; Guimarães, J.T.; Esmerino, E.A.; Duarte, M.C.K.; Silva, M.C.; Silva, R.; Ferreira, B.M.; Sant’Ana, A.S.; Freitas, M.Q.; Cruz, A.G. Paraprobiotics and postbiotics: Concepts and potential applications in dairy products. Curr. Opin. Food Sci. 2020, 32, 1–8. [Google Scholar] [CrossRef]
- Cummings, J.H.; Macfarlane, G.T.; Englyst, H.N. Prebiotic digestion and fermentation. Am. J. Clin. Nutr. 2001, 73, 415s–420s. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Szkaradkiewicz, A.K.; Karpinski, T. Probiotics and prebiotics. J. Biol. Earth Sci. 2013, 3, M42–M47. [Google Scholar]
- Kerry, R.G.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Lauzon, H.L.; Dimitroglou, A.; Merrifield, D.L.; Ringø, E.; Davies, S.J. Probiotics and prebiotics: Concepts, definitions and history. In Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics; Merrifield, D.L., Ringø, E., Eds.; WILEY Blackwell: Hoboken, NJ, USA, 2014; pp. 169–184. [Google Scholar]
- Gourbeyre, P.; Denery, S.; Bodinier, M. Probiotics, prebiotics, and synbiotics: Impact on the gut immune system and allergic reactions. J. Leukoc. Biol. 2011, 89, 685–695. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Batista, V.L.; Da Silva, T.F.; De Jesus, L.C.L.; Dias Coelho-Rocha, N.; Barroso, F.A.L.; Tavares, L.M.; Azevedo, V.A.D.C.; Mancha-Agresti, P.D.C.; Drumond, M.M. Probiotics, prebiotics, synbiotics, and paraprobiotics as a therapeutic alternative for intestinal mucositis running head: Alternative treatment for intestinal mucositis. Front. Microbiol. 2020, 11, 2246. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef] [PubMed]
- Shinde, T.; Perera, A.P.; Vemuri, R.; Gondalia, S.V.; Karpe, A.V.; Beale, D.J.; Shastri, S.; Southam, B.; Eri, R.; Stanley, R. Synbiotic supplementation containing whole plant sugar cane fibre and probiotic spores potentiates protective synergistic effects in mouse model of IBD. Nutrients 2019, 11, 818. [Google Scholar] [CrossRef] [Green Version]
- Sarfraz, F.; Farooq, U.; Shafi, A.; Hayat, Z.; Akram, K.; Rehman, H.-U. Hypolipidaemic effects of synbiotic yoghurt in rabbits. Int. J. Dairy Technol. 2019, 72, 545–550. [Google Scholar] [CrossRef]
- Park, J. Gluco-oligosaccharides produced from Weissella cibaria as potential prebiotics. Master’s Thesis, Gachon University, Seongnam, Korea, February 2021. [Google Scholar]
- Kaur, J.; Lee, S.; Park, Y.-S.; Sharma, A. RAPD analysis of Leuconostoc mesenteroides strains associated with vegetables and food products from Korea. LWT Food Sci. Technol. 2017, 77, 383–388. [Google Scholar] [CrossRef]
- Sharma, A.; Kaur, J.; Lee, S.; Park, Y.-S. Tracking of deliberately inoculated Leuconostoc mesenteroides and Lactobacillus brevis in kimchi. Food Sci. Biotechnol. 2020, 29, 817–824. [Google Scholar] [CrossRef]
- Kim, S. Optimization of semi-continuous fermentation for the production of gluco-oligosaccharides using Weissella cibaria. Master’s Thesis, Gachon University, Seongnam, Korea, February 2021. [Google Scholar]
- Kim, M.; Jang, J.-K.; Park, Y.-S. Production optimization, structural analysis, and prebiotic- and anti-inflammatory effects of gluco-oligosaccharides produced by Leuconostoc lactis SBC001. Microorganisms 2021, 9, 200. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, S.; Wang, M.; Zhao, B.; Yang, N.; Li, J.; Chen, Q.; Liu, M.; Zhou, J.; Bao, G. Characterization and establishment of an immortalized rabbit melanocyte cell line using the SV40 large T antigen. Int. J. Mol. Sci. 2019, 20, 4874. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef]
- Yu, H.-S.; Jang, H.J.; Lee, N.-K.; Paik, H.-D. Evaluation of the probiotic characteristics and prophylactic potential of Weissella cibaria strains isolated from kimchi. LWT Food Sci. Technol. 2019, 112, 108229. [Google Scholar] [CrossRef]
- Tang, W.; Xing, Z.; Li, C.; Wang, J.; Wang, Y. Molecular mechanisms and in vitro antioxidant effects of Lactobacillus plantarum MA2. Food Chem. 2017, 221, 1642–1649. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Lin, W.-H.; Hwang, C.-F.; Chen, L.-W.; Tsen, H.-Y. Viable counts, characteristic evaluation for commercial lactic acid bacteria products. Food Microbiol. 2006, 23, 74–81. [Google Scholar] [CrossRef]
- Chang, C.-K.; Wang, S.-C.; Chiu, C.-K.; Chen, S.-Y.; Chen, Z.-T.; Duh, P.-D. Effect of lactic acid bacteria isolated from fermented mustard on immunopotentiating activity. Asian Pac. J. Trop. Biomed. 2015, 5, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.W.; Cho, S.B.; Yun, C.H.; Jeong, H.Y.; Chung, W.T.; Choi, C.W.; Lee, H.J.; Nam, I.S.; Suh, G.H.; Lee, S.S. Induction of cytokines and nitric oxide in murine macrophages stimulated with enzymatically digested lactobacillus strains. J. Microbiol. 2007, 45, 373–378. [Google Scholar] [PubMed]
- Sarkar, D.; Fisher, P.B. Molecular mechanisms of aging-associated inflammation. Cancer Lett. 2006, 236, 13–23. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Ayad, E.H.; Verheul, A.; de Jong, C.; Wouters, J.T.; Smit, G. Flavour forming abilities and amino acid requirements of Lactococcus lactis strains isolated from artisanal and non-dairy origin. Int. Dairy J. 1999, 9, 725–735. [Google Scholar] [CrossRef]
- Gutiérrez-Méndez, N.; Rodríguez-Figueroa, J.C.; González-Córdova, A.F.; Nevárez-Moorillón, G.V.; Rivera-Chavira, B.; Vallejo-Cordoba, B. Phenotypic and genotypic characteristics of Lactococcus lactis strains isolated from different ecosystems. Can. J. Microbiol. 2010, 56, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, G.-G.; Jang, J.-K.; Park, Y.-S. Optimization of oligosaccharide production from Leuconostoc lactis using a response surface methodology and the immunostimulating effects of these oligosaccharides on macrophage cells. Molecules 2018, 23, 2118. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Song, I.H.; Park, Y.-S. In vivo and In vitro study of immunostimulation by Leuconostoc lactis-produced gluco-oligosaccharides. Molecules 2019, 24, 3994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Park, J.; Jang, J.-K.; Lee, B.-H.; Park, Y.-S. Structural analysis of gluco-oligosaccharides produced by Leuconostoc lactis and their prebiotic effect. Molecules 2019, 24, 3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, K.-L.; Meng, L.-Z.; Chen, X.-Q.; Wang, L.-Y.; Wu, D.-T.; Zhao, J.; Li, S.-P. Structural elucidation, chain conformation and immuno-modulatory activity of glucogalactomannan from cultured Cordyceps sinensis fungus UM01. J. Func. Foods 2016, 25, 174–185. [Google Scholar] [CrossRef]
- Li, Q.Z.; Chang, Y.Z.; He, Z.M.; Chen, L.; Zhou, X.W. Immunomodulatory activity of Ganoderma lucidum immunomodulatory protein via PI3K/Akt and MAPK signaling pathways in RAW264. 7 cells. J. Cell. Physiol. 2019, 234, 23337–23348. [Google Scholar] [CrossRef]
- Yu, Y.; Shen, M.; Wang, Z.; Wang, Y.; Xie, M.; Xie, J. Sulfated polysaccharide from Cyclocarya paliurus enhances the immunomodulatory activity of macrophages. Carbohydr. Polym. 2017, 174, 669–676. [Google Scholar] [CrossRef]
- Xu, X.; Yan, H.; Zhang, X. Structure and immuno-stimulating activities of a new heteropolysaccharide from Lentinula edodes. J. Agric. Food Chem. 2012, 60, 11560–11566. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.T.; Jang, W.J.; Kim, H.; Lee, B.-J.; Kim, K.W.; Hur, S.W.; Lim, S.G.; Bai, S.C.; Kong, I.-S. Synergistic effects of dietary Bacillus sp. SJ-10 plus β-glucooligosaccharides as a synbiotic on growth performance, innate immunity and streptococcosis resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2018, 82, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.T.; Jang, W.J.; Tak, J.Y.; Lee, B.-J.; Kim, K.W.; Hur, S.W.; Han, H.-S.; Kim, B.-S.; Huh, M.-D.; Kim, S.-K. Effects of Lactococcus lactis subsp. lactis I2 with β-glucooligosaccharides on growth, innate immunity and streptococcosis resistance in olive flounder (Paralichthys olivaceus). J. Microbiol. Biotechnol. 2018, 28, 1433–1442. [Google Scholar] [CrossRef]
- Liu, Q.; Xiao, X.-H.; Hu, L.-B.; Jie, H.-Y.; Wang, Y.; Ye, W.-C.; Li, M.-M.; Liu, Z. Anhuienoside C ameliorates collagen-induced arthritis through inhibition of MAPK and NF-κB signaling pathways. Front. Pharmacol. 2017, 8, 299. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.J.; Cobb, M.H. Mitogen-activated protein kinase pathways. Curr. Opin. Cell Biol. 1997, 9, 180–186. [Google Scholar] [CrossRef]
- Song, M.; Han, L.; Chen, F.-F.; Wang, D.; Wang, F.; Zhang, L.; Wang, Z.-H.; Zhong, M.; Tang, M.-X.; Zhang, W. Adipocyte-derived exosomes carrying sonic hedgehog mediate M1 macrophage polarization-induced insulin resistance via Patch and PI3K pathways. Cell. Physiol. Biochem. 2018, 48, 1416–1432. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xing, R.; Liu, S.; Qin, Y.; Li, K.; Yu, H.; Li, P. Hydroxypropyltrimethyl ammonium chloride chitosan activates RAW 264.7 macrophages through the MAPK and JAK-STAT signaling pathways. Carbohydr. Polym. 2019, 205, 401–440. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xing, R.; Liu, S.; Qin, Y.; Li, K.; Yu, H.; Li, P. Immunostimulatory effects of sulfated chitosans on RAW 264.7 mouse macrophages via the activation of PI3 K/Akt signaling pathway. Int. J. Biol. Macromol. 2018, 108, 1310–1321. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, A.; Park, Y.-S. Immunostimulatory Activity of Synbiotics Using Lactococcus lactis SG-030 and Glucooligosaccharides from Weissella cibaria YRK005. Microorganisms 2021, 9, 2437. https://doi.org/10.3390/microorganisms9122437
Kwon A, Park Y-S. Immunostimulatory Activity of Synbiotics Using Lactococcus lactis SG-030 and Glucooligosaccharides from Weissella cibaria YRK005. Microorganisms. 2021; 9(12):2437. https://doi.org/10.3390/microorganisms9122437
Chicago/Turabian StyleKwon, Ayeon, and Young-Seo Park. 2021. "Immunostimulatory Activity of Synbiotics Using Lactococcus lactis SG-030 and Glucooligosaccharides from Weissella cibaria YRK005" Microorganisms 9, no. 12: 2437. https://doi.org/10.3390/microorganisms9122437
APA StyleKwon, A., & Park, Y. -S. (2021). Immunostimulatory Activity of Synbiotics Using Lactococcus lactis SG-030 and Glucooligosaccharides from Weissella cibaria YRK005. Microorganisms, 9(12), 2437. https://doi.org/10.3390/microorganisms9122437