
Drought and Plant Community Composition Affect the Metabolic and Genotypic Diversity of Pseudomonas Strains in Grassland Soils


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Soil Sampling
2.2. Soil Parameters
2.3. Selective Isolation and Cultivation of Pseudomonas Bacteria
2.4. DNA Extraction and Partial 16S rRNA Gene Sequencing
2.5. Quantification of P- and K- Solubilizing Activity and Siderophore Production
2.6. Statistical Analysis
3. Results
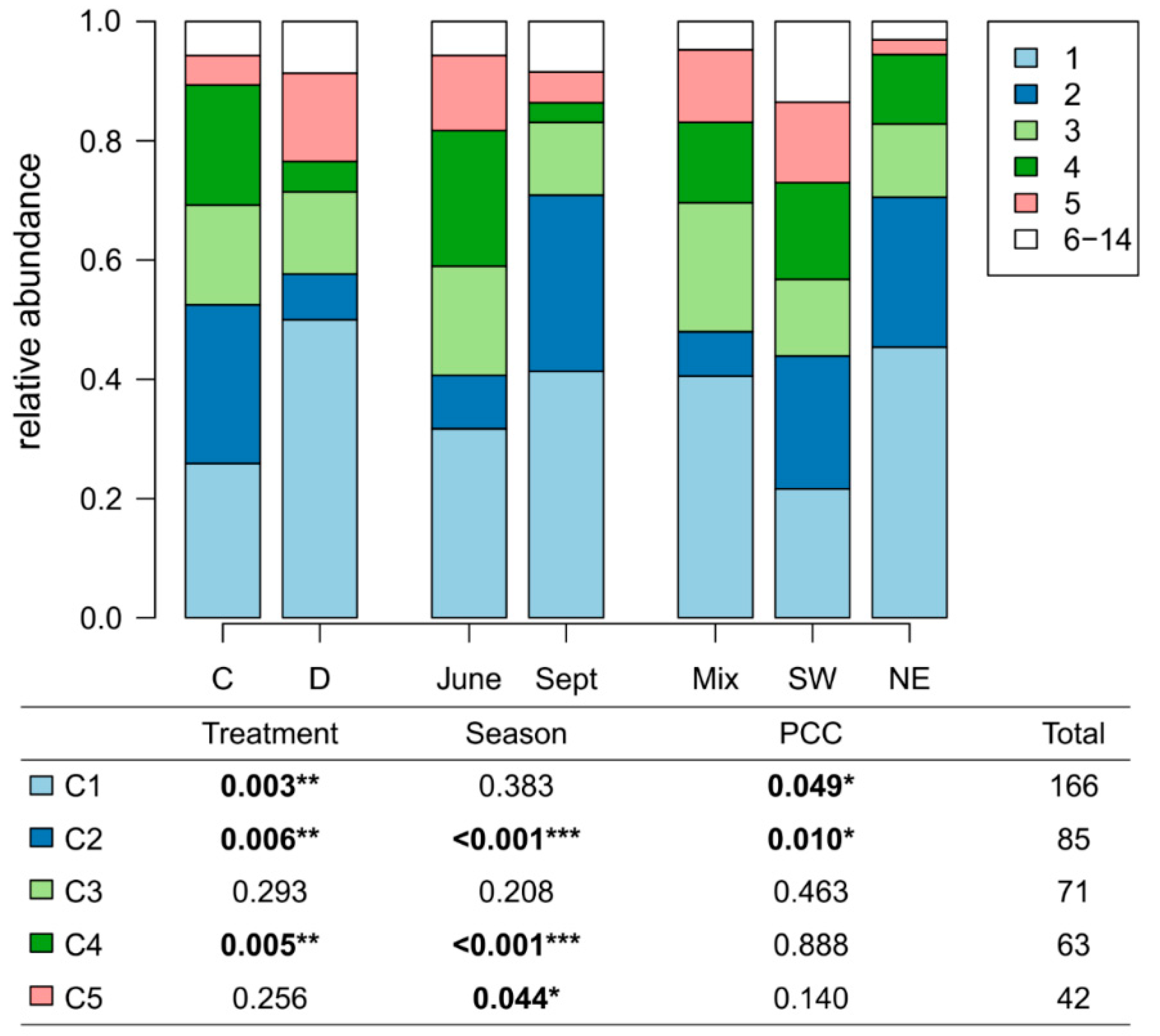
3.1. Isolation of Pseudomonas Strains
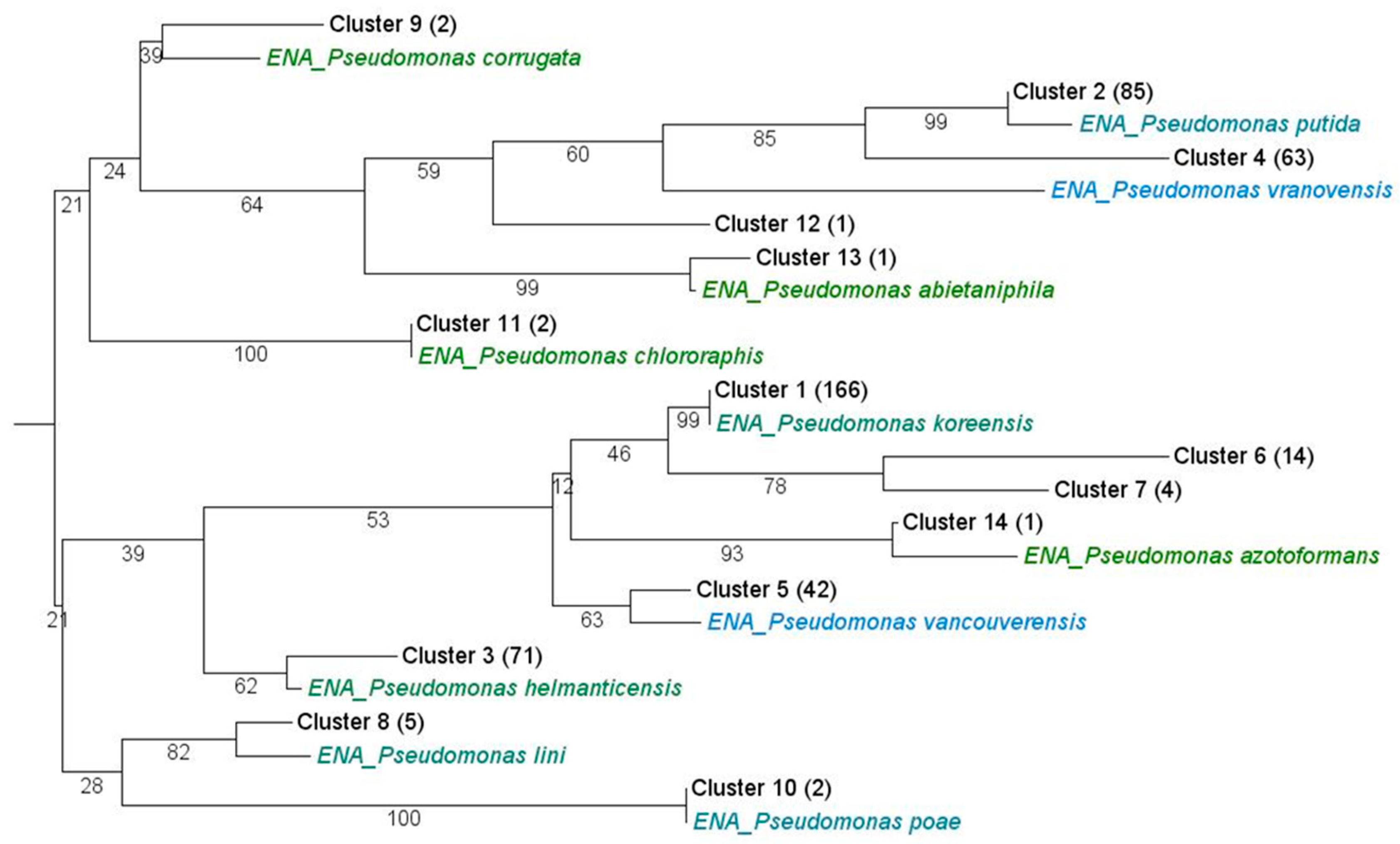
3.2. Phylogenetic Classification and Distribution of the Isolates
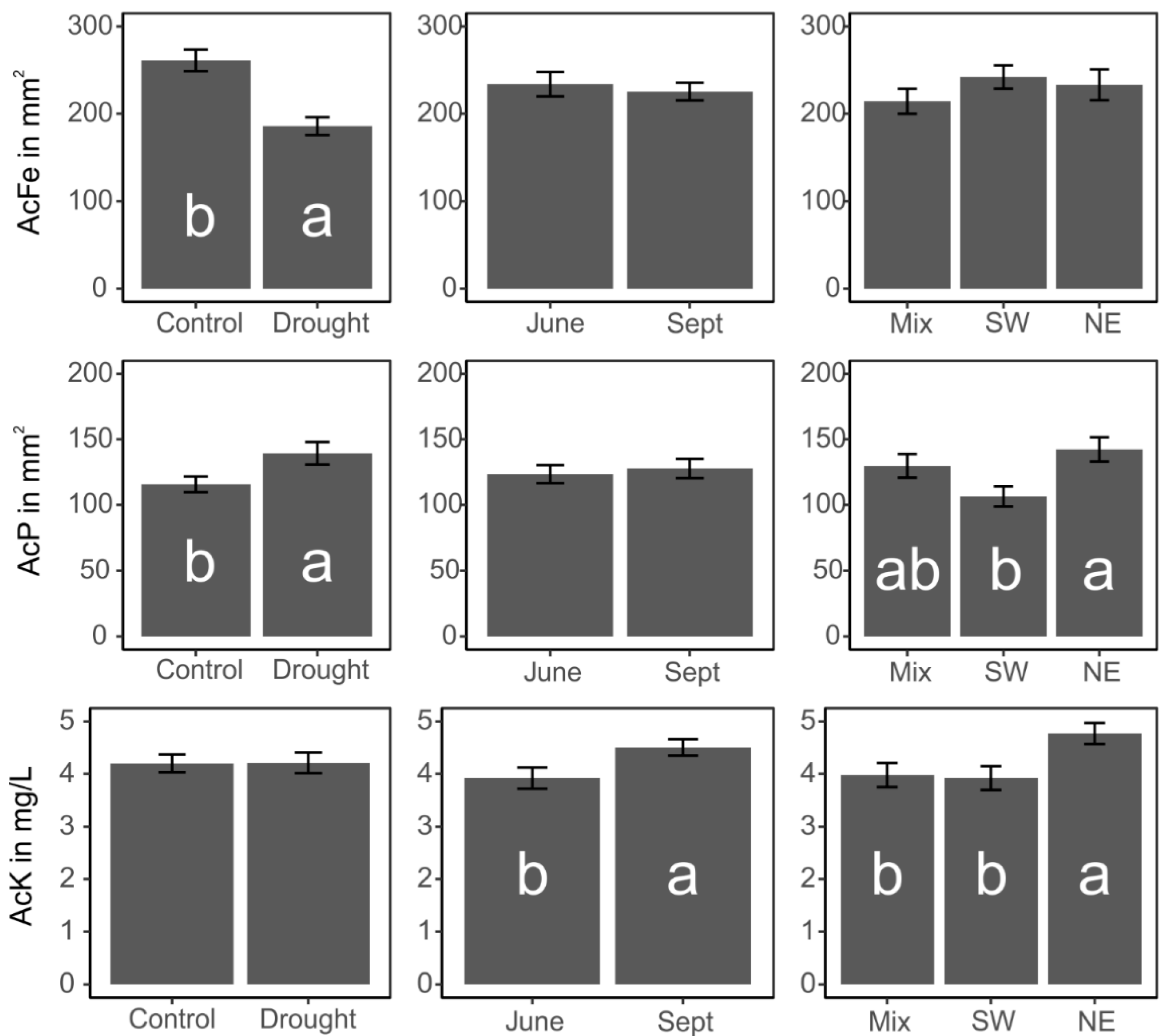
3.3. Impact on Functionality of the Isolated Pseudomonas Strains
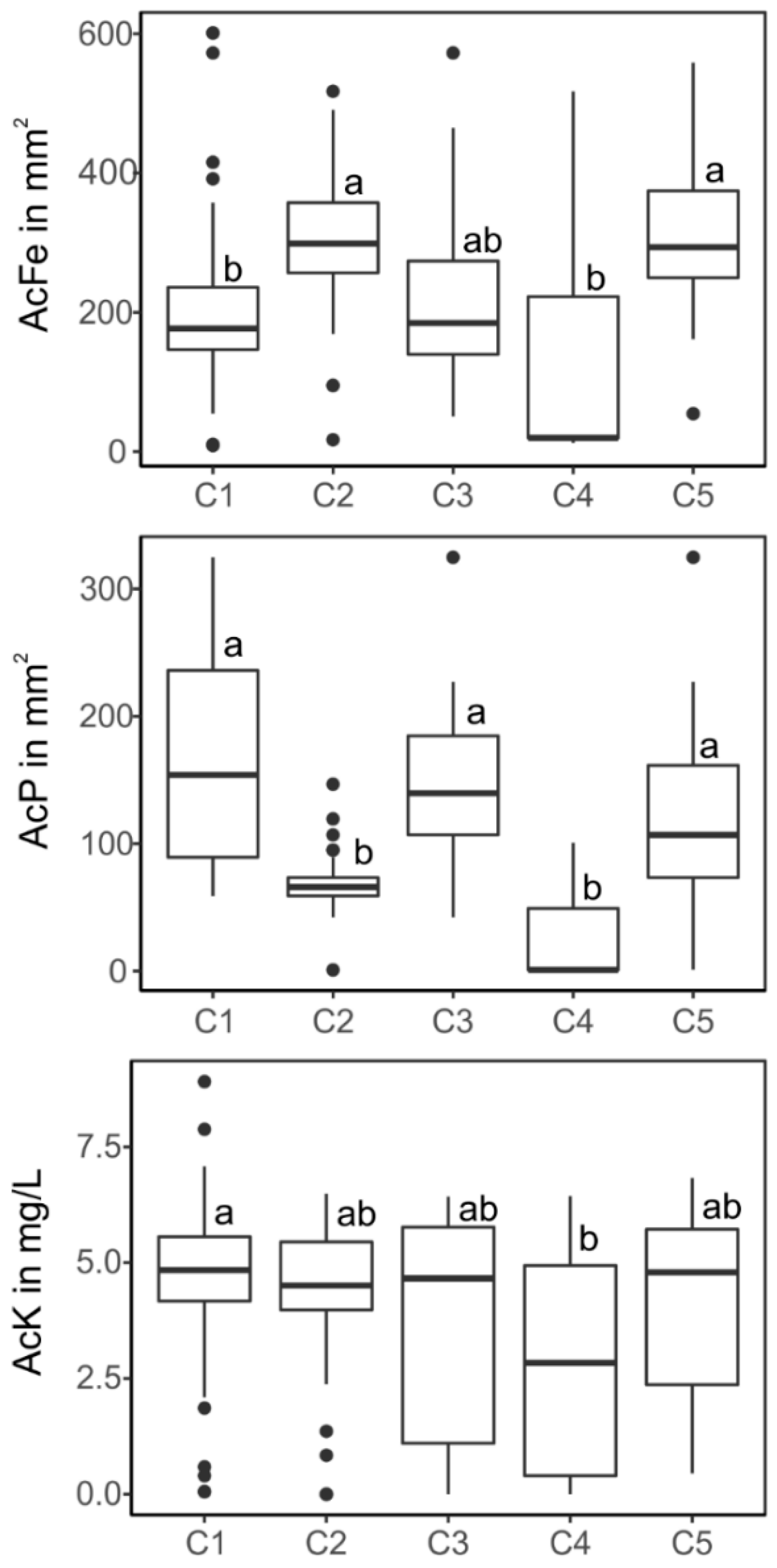
3.4. Relationships between Phylogenetic Classification and Functional Traits of the Isolates
4. Discussion
4.1. Phylogenetic Identity of Pseudomonads
4.2. Effects of the Experimental Treatments on Functional Properties of the Pseudomonas Community
4.3. Links between Phylogeny and Function
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in bacterial taxonomy: Impact on the genus Pseudomonas. Genes 2020, 11, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugtenberg, B.J.J.; Bloemberg, G.V. Life in the Rhizosphere. In Pseudomonas: Volume 1 Genomics, Life Style and Molecular Architecture; Ramos, J.-L., Ed.; Springer: Boston, MA, USA, 2004; Volume 1, pp. 403–430. [Google Scholar]
- Haas, D.; Defago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Panjiar, N.; Kumar, S.; Saxena, A.K.; Suman, A. Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Ann. Microbiol. 2015, 65, 1885–1899. [Google Scholar] [CrossRef]
- Gebauer, L.; Bouffaud, M.L.; Ganther, M.; Yim, B.; Vetterlein, D.; Smalla, K.; Buscot, F.; Heintz-Buschart, A.; Tarkka, M.T. Soil texture, sampling depth and root hairs shape the structure of ACC deaminase bacterial community composition in maize rhizosphere. Front. Microbiol. 2021, 12, 616828. [Google Scholar] [CrossRef]
- Rainey, P.B. Adaptation of Pseudomonas fluorescens to the plant rhizosphere. Environ. Microbiol. 1999, 1, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Binkley, D.; Doxtader, K.G. A new method for estimating gross phosphorus mineralization and immobilization rates in soils. Plant Soil 1992, 147, 243–250. [Google Scholar] [CrossRef]
- Goswami, D.; Vaghela, H.; Parmar, S.; Dhandhukia, P.; Thakker, J.N. Plant growth promoting potentials of Pseudomonas spp. strain OG isolated from marine water. J. Plant Interact. 2013, 8, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Peix, A.; Rivas, R.; Mateos, P.F.; Martinez-Molina, E.; Rodriguez-Barrueco, C.; Velazquez, E. Pseudomonas rhizosphaerae sp nov., a novel species that actively solubilizes phosphate in vitro. Int. J. Syst. Evol. Microbiol. 2003, 53, 2067–2072. [Google Scholar] [CrossRef] [Green Version]
- Gulati, A.; Rahi, P.; Vyas, P. Characterization of phosphate-solubilizing fluorescent pseudomonads from the rhizosphere of seabuckthorn growing in the cold deserts of Himalayas. Curr. Microbiol. 2008, 56, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Rekha, P.D.; Arun, A.B.; Shen, F.T.; Lai, W.A.; Young, C.C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Gyaneshwar, P.; Kumar, G.N.; Parekh, L.J.; Poole, P.S. Role of soil microorganisms in improving P nutrition of plants. Plant Soil 2002, 245, 83–93. [Google Scholar] [CrossRef]
- Sharma, A.; Johri, B.N. Growth promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9 in maize (Zea mays L.) under iron limiting conditions. Microbiol. Res. 2003, 158, 243–248. [Google Scholar] [CrossRef]
- Barness, E.; Chen, Y.; Hadar, Y.; Marschner, H.; Romheld, V. Siderophores of Pseudomonas putida as an iron source for dicot and monocot plants. Plant Soil 1991, 130, 231–241. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Pseudomonas siderophores – a mechanism explaining disease-suppressive soils. Curr. Microbiol. 1980, 4, 317–320. [Google Scholar] [CrossRef]
- Castro, H.F.; Classen, A.T.; Austin, E.E.; Norby, R.J.; Schadt, C.W. Soil Microbial Community Responses to Multiple Experimental Climate Change Drivers. Appl. Environ. Microbiol. 2010, 76, 999–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Wang, C.; Luo, Y. Response of soil microbial communities to altered precipitation: A global synthesis. Glob. Ecol. Biogeogr. 2018, 27, 1121–1136. [Google Scholar] [CrossRef]
- Pritchard, S.G. Soil organisms and global climate change. Plant Pathol. 2011, 60, 82–99. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Fry, E.L.; Eldridge, D.J.; de Vries, F.T.; Manning, P.; Hamonts, K.; Kattge, J.; Boenisch, G.; Singh, B.K.; Bardgett, R.D. Plant attributes explain the distribution of soil microbial communities in two contrasting regions of the globe. New Phytol. 2018, 219, 574–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gornish, E.S.; Tylianakis, J.M. Community shifts under climate change: Mechanisms at multiple scales. Am. J. Bot. 2013, 100, 1422–1434. [Google Scholar] [CrossRef]
- Sriswasdi, S.; Yang, C.C.; Iwasaki, W. Generalist species drive microbial dispersion and evolution. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macleod, K.; Rumbold, K.; Padayachee, K. A systems approach to uncover the effects of the PGPR Pseudomonas koreensis on the level of drought stress tolerance in Helianthus annuus. Procedia. Environ. Sci. 2015, 29, 262–263. [Google Scholar] [CrossRef] [Green Version]
- Sandhya, V.Z.A.S.; Grover, M.; Reddy, G.; Venkateswarlu, B. Alleviation of drought stress effects in sunflower seedlings by the exopolysaccharides producing Pseudomonas putida strain GAP-P45. Biol. Fertil. Soils 2009, 46, 17–26. [Google Scholar] [CrossRef]
- Classen, A.T.; Sundqvist, M.K.; Henning, J.A.; Newman, G.S.; Moore, J.A.M.; Cregger, M.A.; Moorhead, L.C.; Patterson, C.M. Direct and indirect effects of climate change on soil microbial and soil microbial-plant interactions: What lies ahead? Ecosphere 2015, 6, 130. [Google Scholar] [CrossRef]
- Marasco, R.; Rolli, E.; Ettoumi, B.; Vigani, G.; Mapelli, F.; Borin, S.; Abou-Hadid, A.F.; El-Behairy, U.A.; Sorlini, C.; Cherif, A.; et al. A drought resistance-promoting microbiome is selected by root system under desert farming. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Altermann, M.; Rinklebe, J.; Merbach, I.; Körschens, M.; Langer, U.; Hofmann, B. Chernozem – soil of the year 2005. J. Plant Nutr. Soil Sci. 2005, 168, 725–740. [Google Scholar] [CrossRef]
- Riehm, H. Arbeitsvorschrift zur Bestimmung der Phosphorsäure und des Kaliums nach Lactatverfahren. Z. Für Pflanz. Düngung Bodenkd. 1948, 40, 61–74. [Google Scholar]
- Murphy, J.A.M.E.S.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Krueger, C.L.; Sheikh, W. A new selective medium for isolating Pseudomonas spp. from water. Appl. Environ. Microbiol. 1987, 53, 895–897. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.C.; Dong, H.L.; Zhang, G.X.; Yu, B.S.; Chapman, L.R.; Fields, M.W. Microbial diversity in water and sediment of Lake Chaka, an athalassohaline lake in northwestern China. Appl. Environ. Microbiol. 2006, 72, 3832–3845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Niu, B.F.; Gao, Y.; Fu, L.M.; Li, W.Z. CD-HIT Suite: A web server for clustering and comparing biological sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Han, M.V.; Zmasek, C.M. phyloXML: XML for evolutionary biology and comparative genomics. BMC Bioinform. 2009, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pikovskaya, R.I. Mobilization of phosphorus in soil connection with the vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of blue agar CAS assay for siderophore detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmar, P.; Sindhu, S.S. Potassium solubilization by rhizosphere bacteria: Influence of nutritional and environmental conditions. J. Microbiol. Res. 2013, 3, 25–31. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2019. Available online: https://www.R-project.org/ (accessed on 5 January 2021).
- Bloemberg, G.V.; Lugtenberg, B.J.J. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Mahdavikia, H.; Rezaei-Chiyaneh, E.; Rahimi, A.; Mohammadkhani, N. Effects of fertilizer treatments on antioxidant activities and physiological traits of basil (Ocimum basilicum L.) under water limitation conditions. J. Med. Plants By-Prod. 2019, 8, 143–151. [Google Scholar]
- Kwon, S.W.; Kim, J.S.; Park, I.C.; Yoon, S.H.; Park, D.H.; Lim, C.K.; Go, S.J. Pseudomonas koreensis sp nov., Pseudomonas umsongensis sp nov and Pseudomonas jinjuensis sp nov., novel species from farm soils in Korea. Int. J. Syst. Evol. Microbiol. 2003, 53, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, A.; Khan, M.A.; Lee, K.E.; Kang, S.M.; Dhungana, S.K.; Bhusal, N.; Lee, I.J. The halotolerant rhizobacterium—Pseudomonas koreensis MU2 enhances inorganic silicon and phosphorus use efficiency and augments salt stress tolerance in soybean (Glycine max L.). Microorganisms 2020, 8, 1256. [Google Scholar] [CrossRef]
- Mursyida, E.; Mubarik, N.R.; Tjahjoleksono, A. selection and identification of phosphate-potassium solubilizing bacteria from the area around the limestone mining in Cirebon Quarry. Res. J. Microbiol. 2015, 10, 270–279. [Google Scholar]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Saxena, A.K.; Suman, A. Potassium-solubilizing microbes: Diversity, distribution, and role in plant growth promotion. In Microorganisms for Green Revolution; Springer: Singapore, 2017; pp. 125–149. [Google Scholar]
- Scher, F.M.; Baker, R. Effect of Pseudomonas putida and synthetic iron chelator on induction of soil suppressiveness to Fusarium-wilt pathogens. Phytopathology 1982, 72, 1567–1573. [Google Scholar] [CrossRef]
- Mazzola, M.; Gu, Y.H. Impact of wheat cultivation on microbial communities from replant soils and apple growth in greenhouse trials. Phytopathology 2000, 90, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Martínez, K.; Suttle, K.B.; Brodie, E.L.; Power, M.E.; Andersen, G.L.; Banfield, J.F. Despite strong seasonal responses, soil microbial consortia are more resilient to long-term changes in rainfall than overlying grassland. ISME J. 2009, 3, 738–744. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Whiteley, A.S.; O’Donnell, A.G.; Bailey, M.J. Physiological and community responses of established grassland bacterial populations to water stress. Appl. Environ. Microbiol. 2003, 69, 6961–6968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardgett, R.D.; Caruso, T. Soil microbial community responses to climate extremes: Resistance, resilience and transitions to alternative states. Philos. Trans. R. Soc. B 2020, 375, 20190112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach. Wiley Sons 1995. Available online: https://books.google.de/books?id=UKXj6Xpc2PIC (accessed on 25 June 2021).
- Crowley, D.E. Microbial Siderophores in the Plant Rhizosphere. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Barton, L.L., Abadia, J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 169–198. [Google Scholar]
- Buckling, A.; Harrison, F.; Vos, M.; Brockhurst, M.A.; Gardner, A.; West, S.A.; Griffin, A. Siderophore-mediated cooperation and virulence in Pseudomonas aeruginosa. FEMS Microbiol. Ecol. 2007, 62, 135–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirinbayan, S.; Khosravi, H.; Malakouti, M.J. Alleviation of drought stress in maize (Zea mays) by inoculation with Azotobacter strains isolated from semi-arid regions. Appl. Soil Ecol. 2019, 133, 138–145. [Google Scholar] [CrossRef]
- Kumar, M.; Mishra, S.; Dixit, V.; Kumar, M.; Agarwal, L.; Chauhan, P.S.; Nautiyal, C.S. Synergistic effect of Pseudomonas putida and Bacillus amyloliquefaciens ameliorates drought stress in chickpea (Cicer arietinum L.). Plant Signal. Behav. 2016, 11, e1071004. [Google Scholar] [CrossRef] [Green Version]
- Latour, X.; Corberand, T.S.; Laguerre, G.; Allard, F.; Lemanceau, P. The composition of fluorescent pseudomonad populations associated with roots is influenced by plant and soil type. Appl. Environ. Microbiol. 1996, 62, 2449–2456. [Google Scholar] [CrossRef] [Green Version]
- Schreiter, S.; Babin, D.; Smalla, K.; Grosch, R. Rhizosphere competence and biocontrol effect of Pseudomonas sp. RU47 independent from plant species and soil type at the field scale. Front. Microbiol. 2018, 9, 97. [Google Scholar]
- Hobbie, S.E. Effects of plant species on nutrient cycling. Trends Ecol. Evol. 1992, 7, 336–339. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Che, R.X.; Wang, P.; Liu, H.K.; Ji, B.M.; Cui, X.Y. Precipitation shapes communities of arbuscular mycorrhizal fungi in Tibetan alpine steppe. Sci. Rep. 2016, 6, 23488. [Google Scholar] [CrossRef]
- Martiny, J.B.H.; Jones, S.E.; Lennon, J.T.; Martiny, A.C. Microbiomes in light of traits: A phylogenetic perspective. Science 2015, 350, aac9323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clays-Josserand, A.; Ghiglione, J.F.; Philippot, L.; Lemanceau, P.; Lensi, R. Effect of soil type and plant species on the fluorescent pseudomonads nitrate dissimilating community. Plant Soil 1999, 209, 275–282. [Google Scholar] [CrossRef]
- Meyer, A.F.; Lipson, D.A.; Martin, A.P.; Schadt, C.W.; Schmidt, S.K. Molecular and metabolic characterization of cold-tolerant alpine soil Pseudomonas sensu stricto. Appl. Environ. Microbiol. 2004, 70, 483–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aulakh, M.S.; Wassmann, R.; Bueno, C.; Kreuzwieser, J.; Rennenberg, H. Characterization of root exudates at different growth stages of ten rice (Oryza sativa L.) cultivars. Plant Biol. 2001, 3, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Iannucci, A.; Fragasso, M.; Platani, C.; Papa, R. Plant growth and phenolic compounds in the rhizosphere soil of wild oat (Avena fatua L.). Front. Plant Sci. 2013, 4, 509. [Google Scholar] [CrossRef] [Green Version]
- Thomas-Barry, G.; Martin, C.C.S.; Lynch, M.D.; Ramsubhag, A.; Rouse-Miller, J.; Charles, T.C. Driving factors influencing the rhizobacteriome community structure of plants adapted to multiple climatic stressors in edaphic savannas. Sci. Total Environ. 2021, 769, 145214. [Google Scholar] [CrossRef]
- Breitkreuz, C.; Buscot, F.; Tarkka, M.; Reitz, T. Shifts between and among populations of wheat rhizosphere Pseudomonas, Streptomyces and Phyllobacterium suggest consistent phosphate mobilization at different wheat growth stages under abiotic stress. Front. Microbiol. 2020, 10, 3109. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Moisture | Soil P | TOC | TN | PB | RB | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AcFe | −0.22 | 0.10 | 0.34 | 0.01 * | −0.28 | 0.04 * | −0.11 | 0.41 | −0.11 | 0.44 | 0.18 | 0.18 | 0.02 | 0.88 |
| AcP | 0.10 | 0.46 | −0.33 | 0.01 * | 0.23 | 0.09 | 0.05 | 0.70 | 0.11 | 0.41 | −0.18 | 0.20 | −0.03 | 0.83 |
| AcK | −0.16 | 0.23 | −0.09 | 0.52 | 0.15 | 0.29 | 0.08 | 0.58 | 0.11 | 0.42 | 0.21 | 0.13 | 0.14 | 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breitkreuz, C.; Reitz, T.; Schulz, E.; Tarkka, M.T. Drought and Plant Community Composition Affect the Metabolic and Genotypic Diversity of Pseudomonas Strains in Grassland Soils. Microorganisms 2021, 9, 1677. https://doi.org/10.3390/microorganisms9081677
Breitkreuz C, Reitz T, Schulz E, Tarkka MT. Drought and Plant Community Composition Affect the Metabolic and Genotypic Diversity of Pseudomonas Strains in Grassland Soils. Microorganisms. 2021; 9(8):1677. https://doi.org/10.3390/microorganisms9081677
Chicago/Turabian StyleBreitkreuz, Claudia, Thomas Reitz, Elke Schulz, and Mika Tapio Tarkka. 2021. "Drought and Plant Community Composition Affect the Metabolic and Genotypic Diversity of Pseudomonas Strains in Grassland Soils" Microorganisms 9, no. 8: 1677. https://doi.org/10.3390/microorganisms9081677
APA StyleBreitkreuz, C., Reitz, T., Schulz, E., & Tarkka, M. T. (2021). Drought and Plant Community Composition Affect the Metabolic and Genotypic Diversity of Pseudomonas Strains in Grassland Soils. Microorganisms, 9(8), 1677. https://doi.org/10.3390/microorganisms9081677