A Novel SNPs in Alpha-Lactalbumin Gene Effects on Lactation Traits in Chinese Holstein Dairy Cows

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Preparation
2.2. PCR-SSCP Analysis
2.3. PCR Products Sequencing and Alignment
2.4. Bovine α-LA Protein Spatial Structure Prediction Modeling
2.5. SNPs Association Analysis
3. Results
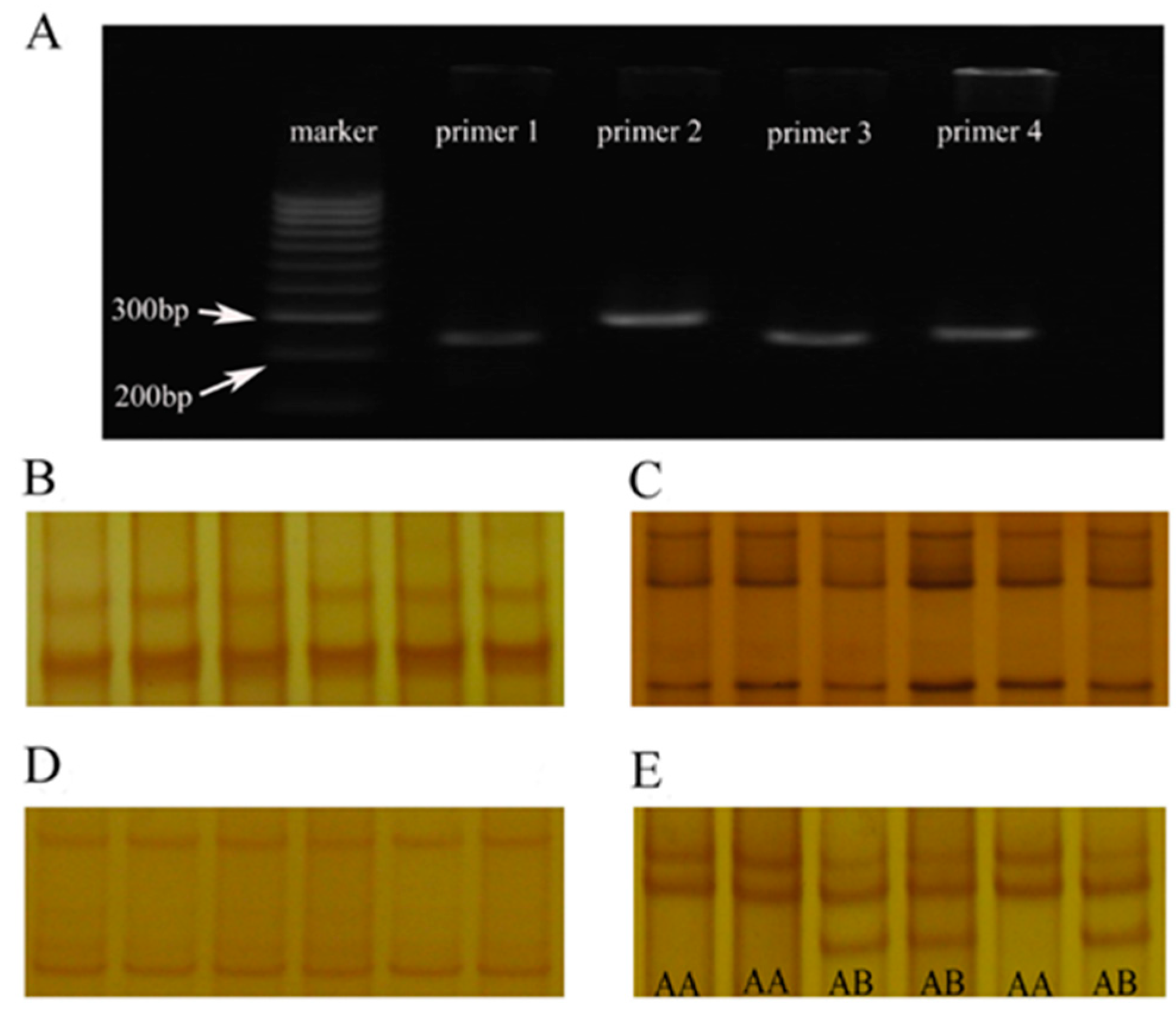
3.1. The PCR Amplification Products of the Primer 4 have SSCP Polymorphism
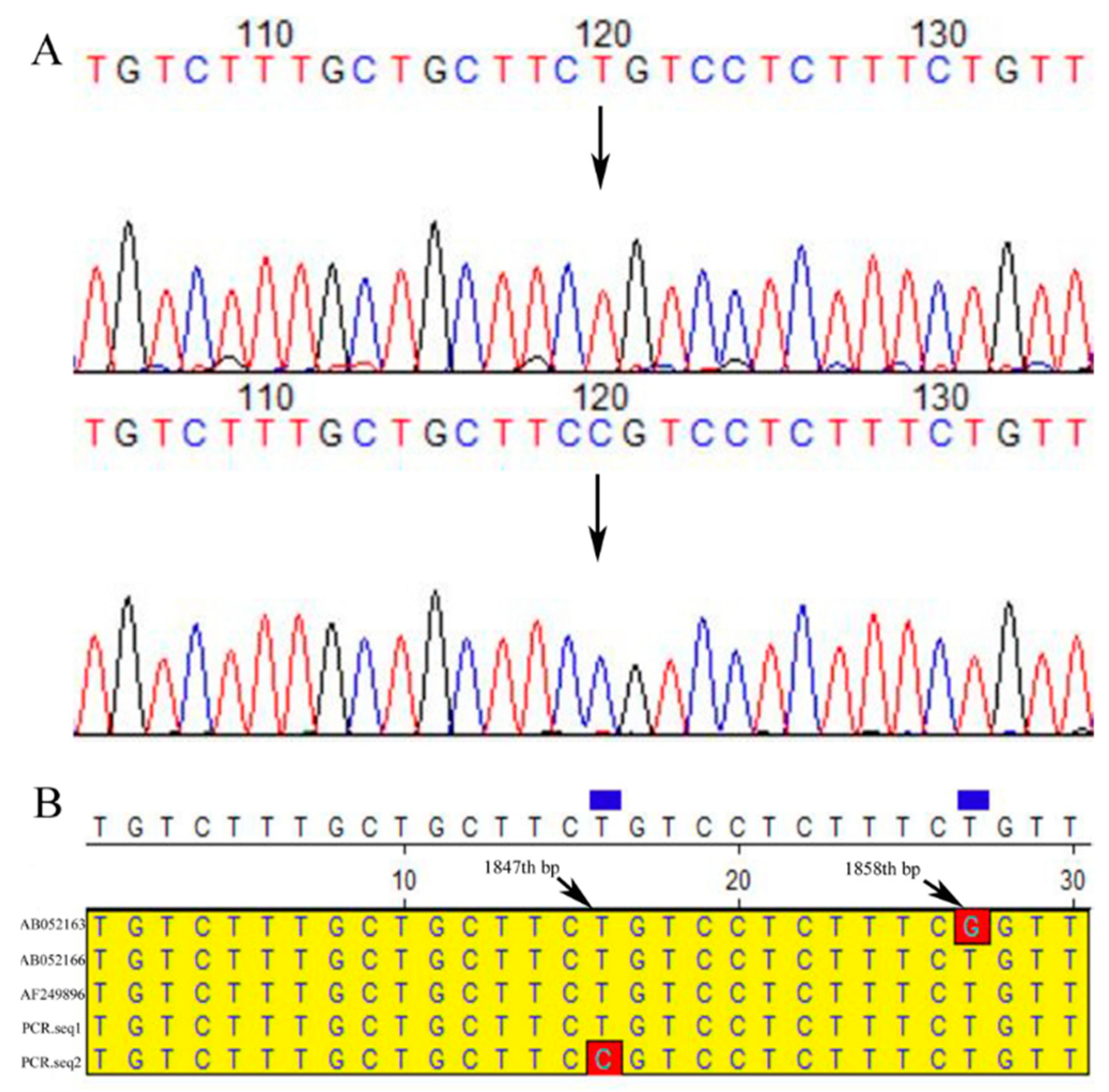
3.2. Alignment Analysis of the Four Primers’ PCR Amplification Products
3.3. SNPs of CDS 2 Region (562th, G/A) in the α-LA Gene Affect the Spatial Structure
3.4. Population Genetic Analysis Parameters of α-LA Gene SNPs (1847th, T/C) in Chinese Holstein Dairy Cows
4. Discussion
4.1. SSCP Typing Analysis Identified Two Genotypes (AA and AB Genotype) of α-LA Gene in Chinese Holstein Dairy Cows
4.2. Genetic Polymorphisms (SNPs) of the α-LA Gene Existing in Four Different Bovine Breeds
4.3. One SNPs (1847th (T/C)) Locus in the Noncoding Region Near CDS4 of α-LA Gene Affected the Milk Lactose Content and Yield in Chinese Holstein Dairy Cows
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bittante, G.; Penasa, M.; Cecchinato, A. Invited review: Genetics and modeling of milk coagulation properties. J. Dairy Sci. 2012, 95, 6843–6870. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.P.; Ferrand, M.; Gelé, M.; Pourchet, D.; Miranda, G.; Martin, P.; Brochard, M.; Boichard, D. Short communication: Genetic parameters for milk protein composition predicted using mid-infrared spectroscopy in the French Montbeliarde, Normande, and Holstein dairy cattle breeds. J. Dairy Sci. 2017, 100, 6371–6375. [Google Scholar] [CrossRef] [PubMed]
- Gebreyesus, G.; Lund, M.S.; Janss, L.; Poulsen, N.A.; Larsen, L.B.; Bovenhuis, H.; Buitenhuis, A.J. Short communication: Multi-trait estimation of genetic parameters for milk protein composition in the Danish Holstein. J. Dairy Sci. 2016, 99, 2863–2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, M.P.; Govignon-Gion, A.; Croiseau, P.; Fritz, S.; Hozé, C.; Miranda, G.; Martin, P.; Barbat-Leterrier, A.; Letaïef, R.; Rocha, D.; et al. Within-breed and multi-breed GWAS on imputed whole-genome sequence variants reveal candidate mutations affecting milk protein composition in dairy cattle. Genet. Sel. Evol. 2017, 49, 68. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.P.; Liu, G.Q.; Wang, C.; Mao, Y.J.; Xiong, Y.Z. Milk trait heritability and correlation with heterozygosity in yak. J. Appl. Genet. 2004, 45, 215–224. [Google Scholar]
- Neville, M.C. Introduction: Alpha-lactalbumin, a multifunctional protein that specifies lactose synthesis in the Golgi. J. Mammary Gland. Biol. Neoplasia 2009, 14, 211–212. [Google Scholar] [CrossRef] [Green Version]
- Layman, D.K.; Lonnerda, B.; Fernstrom, J.D. Applications for alpha-lactalbumin in human nutrition. Nutr. Rev. 2018, 76, 444–460. [Google Scholar] [CrossRef]
- Voelker, G.R.; Bleck, G.T.; Wheeler, M.B. Single-base polymorphisms within the 5′ flanking region of the bovine alpha-lactalbumin gene. J. Dairy Sci. 1997, 80, 194–197. [Google Scholar] [CrossRef]
- Shinozaki, R.; Iwaoka, M. Effects of Metal Ions, Temperature, and a Denaturant on the Oxidative Folding Pathways of Bovine alpha-Lactalbumin. Int. J. Mol. Sci. 2017, 18, 1996. [Google Scholar] [CrossRef] [Green Version]
- Meulenbroek, L.A.; den Hartog Jager, C.F.; Lebens, A.F.; Knulst, A.C.; Bruijnzeel-Koomen, C.A.; Garssen, J.; Knippels, L.M.; Van Hoffen, E. Characterization of T cell epitopes in bovine alpha-lactalbumin. Int. Arch. Allergy Immunol. 2014, 163, 292–296. [Google Scholar] [CrossRef]
- Brinkmann, C.R.; Heegaard, C.W.; Petersen, T.E.; Jensenius, J.C.; Thiel, S. The toxicity of bovine alpha-lactalbumin made lethal to tumor cells is highly dependent on oleic acid and induces killing in cancer cell lines and noncancer-derived primary cells. FEBS J. 2011, 278, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Fukawa, A.; Kobayashi, O.; Yamaguchi, M.; Uchida, M.; Hosono, A. Bovine milk-derived alpha-lactalbumin prevents hepatic fibrosis induced by dimethylnitrosamine via nitric oxide pathway in rats. Biosci. Biotechnol. Biochem. 2017, 81, 1941–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, R.N.; Deng, C.J.; Zhang, X.M.; Yue, X.P.; Lan, X.Y.; Chen, H.; Lei, C.Z. S-novel SNP of alpha-lactalbumin gene in chinese dairy goats. Mol. Biol. 2010, 44, 536–540. [Google Scholar] [CrossRef]
- Bleck, G.T.; Johnson-Roberts, D.L.; Jiménez-Flores, R.; Wheeler, M.B. DNA sequence of the porcine alpha-lactalbumin 5′ flanking region and single-base polymorphisms within this region. Anim. Genet. 1995, 26, 101–103. [Google Scholar] [CrossRef]
- Dayal, S.; Bhattacharya, T.K.; Vohra, V.; Kumar, P.; Sharma, A. Genetic polymorphism of alpha-lactalbumin gene in riverine buffalo. DNA Seq. 2005, 16, 173–179. [Google Scholar] [CrossRef]
- Martins, L.F.; Milazzotto, M.P.; Feitosa, W.B.; Coutinho, A.R.; Sim[otilde]es, R.; Marques, M.G.; Assumpcao, M.E.; Visintin, J.A. Sequence variation of the alpha-lactalbumin gene in Holstein and Nellore cows. Anim. Biotechnol. 2008, 19, 194–198. [Google Scholar] [CrossRef]
- Dettori, M.L.; Pazzola, M.; Paschino, P.; Pira, M.G.; Vacca, G.M. Variability of the caprine whey protein genes and their association with milk yield, composition and renneting properties in the Sarda breed. 1. The LALBA gene. J. Dairy Res. 2015, 82, 434–441. [Google Scholar] [CrossRef]
- Kazmer, G.W.; Zhou, P.; Troyer, J.L.; Strausbaugh, L.D. Haplotype analysis involving a novel polymorphism in bovine alpha-lactalbumin 5′ flanking region. J. Dairy Sci. 2001, 84, 1542–1544. [Google Scholar] [CrossRef]
- Visker, M.H.; Heck, J.M.; van Valenberg, H.J.; van Arendonk, J.A.; Bovenhuis, H. Short communication: A new bovine milk-protein variant: Alpha-lactalbumin variant D. J. Dairy Sci. 2012, 95, 2165–2169. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Bonfatti, V.; Cecchinato, A.; Gallo, L.; Blasco, A.; Carnier, P. Genetic analysis of detailed milk protein composition and coagulation properties in Simmental cattle. J. Dairy Sci. 2011, 94, 5183–5193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopen, G.C.; Heck, J.M.; Bovenhuis, H.; Visker, M.H.; Van Valenberg, H.J.; Van Arendonk, J.A. Genetic parameters for major milk proteins in Dutch Holstein-Friesians. J. Dairy Sci. 2009, 92, 1182–1191. [Google Scholar] [CrossRef]
- Poulsen, N.A.; Jensen, H.B.; Larsen, L.B. Factors influencing degree of glycosylation and phosphorylation of caseins in individual cow milk samples. J. Dairy Sci. 2016, 99, 3325–3333. [Google Scholar] [CrossRef] [Green Version]
- Zidi, A.; Casas, E.; Amills, M.; Jordana, J.; Carrizosa, J.; Urrutia, B.; Serradilla, J.M. Genetic variation at the caprine lactalbumin, alpha (LALBA) gene and its association with milk lactose concentration. Anim. Genet. 2014, 45, 612–613. [Google Scholar] [CrossRef]
- Gustavsson, F.; Buitenhuis, A.J.; Johansson, M.; Bertelsen, H.P.; Glantz, M.; Poulsen, N.A.; Månsson, H.L.; Stålhammar, H.; Larsen, L.B.; Bendixen, C.; et al. Effects of breed and casein genetic variants on protein profile in milk from Swedish Red, Danish Holstein, and Danish Jersey cows. J. Dairy Sci. 2014, 97, 3866–3877. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Primer Name | Sequence (5′–3′) | Base Number (bp) | CDS Region (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|---|
| CDS1 | primer1F primer1R | GAGCAGTGTGGTGACCCC TGGAGGGAAAGAGTGAAGAG | 237 (706–942) | 751–883 | 58.7 |
| CDS2 | primer2F primer2R | GGTCTGGGAATACAGGTCC GGTTATCCCAGGAGTAGGTT | 295 (1129–1423) | 1205–1363 | 58.9 |
| CDS3 | primer3F primer3R | GGCAACAGGCATAAGCCTC GGACTGAGAAGAAAGAGAGG | 240 (1727–1965) | 1837–1912 | 58.7 |
| CDS4 | primer4F primer4R | CCTCAGCCTTCCTGGGGA CAGGGCTCAGAGACGAGTT | 255 (2332–2586) | 2417–2477 | 58.8 |
| Items | Genotypic Frequencies | Allele Frequencies | p < 0.01 | |||
|---|---|---|---|---|---|---|
| AA | AB | A | B | |||
| Group 1 | Number | 115 | 37 | |||
| Rates | 0.7566 | 0.2434 | 0.8783 | 0.1217 | ||
| Group 2 | Number | 281 | 102 | |||
| Rates | 0.7337 | 0.2663 | 0.8468 | 0.1332 | ||
| Total | Number | 396 | 139 | |||
| Rates | 0.7402 | 0.2598 | 0.8701 | 0.1299 | ||
| Groups | Sample Size | Na | Ne | H | PIC | Obs.F | Min F | Max F | L95 | U95 |
|---|---|---|---|---|---|---|---|---|---|---|
| Group 1 | 152 | 2 | 1.600 | 0.3750 | 0.5623 | 0.6250 | 0.50 | 0.9869 | 0.5022 | 0.9869 |
| Group 2 | 383 | 2 | 1.648 | 0.3932 | 0.5822 | 0.6098 | 0.50 | 0.9948 | 0.5052 | 0.9948 |
| Mean | 2 | 1.624 | 0.3841 | 0.5723 | 0.6174 | 0.50 | 0.9908 | 0.5037 | 0.9908 | |
| SD | 0 | 0.024 | 0.0091 | 0.00995 | 0.0076 | 0 | 0.00395 | 0.0015 | 0.00395 |
| Items | Genotype/Base | Mean Fat Content (%) | Mean Protein Content (%) | Lactose (%) | 305D (kg) |
|---|---|---|---|---|---|
| Group 1 | AA/T | 3.545 ± 0.3215 | 3.280 ± 0.3050 | 4.668 ± 0.232 ** | 10058.68 ± 822.96 |
| AB/C | 3.602 ± 0.3426 | 3.125 ± 0.3062 | 4.732 ± 0.233 * | 9723.01 ± 1033.12 | |
| Group 2 | AA/T | 3.574 ± 0.3968 | 3.155 ± 0.3040 | 4.793 ± 0.1821 ** | 10187.31 ± 940.89 |
| AB/C | 3.520 ± 0.4747 | 3.200 ± 0.3008 | 4.837 ± 0.2077 ** | 10051.50 ± 1135.12 | |
| Mean ± SD | 3.560 ± 0.3839 | 3.191 ± 0.3058 | 4.757 ± 0.2139 | 10005.27 ± 983.03 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Zhang, M.; Rong, Y.; Liu, Z.; Yang, S.; Zhang, W.; Li, J.; Cai, Y. A Novel SNPs in Alpha-Lactalbumin Gene Effects on Lactation Traits in Chinese Holstein Dairy Cows. Animals 2020, 10, 60. https://doi.org/10.3390/ani10010060
Yang F, Zhang M, Rong Y, Liu Z, Yang S, Zhang W, Li J, Cai Y. A Novel SNPs in Alpha-Lactalbumin Gene Effects on Lactation Traits in Chinese Holstein Dairy Cows. Animals. 2020; 10(1):60. https://doi.org/10.3390/ani10010060
Chicago/Turabian StyleYang, Fan, Manling Zhang, Yuewen Rong, Zaiqun Liu, Shuai Yang, Wei Zhang, Jun Li, and Yafei Cai. 2020. "A Novel SNPs in Alpha-Lactalbumin Gene Effects on Lactation Traits in Chinese Holstein Dairy Cows" Animals 10, no. 1: 60. https://doi.org/10.3390/ani10010060
APA StyleYang, F., Zhang, M., Rong, Y., Liu, Z., Yang, S., Zhang, W., Li, J., & Cai, Y. (2020). A Novel SNPs in Alpha-Lactalbumin Gene Effects on Lactation Traits in Chinese Holstein Dairy Cows. Animals, 10(1), 60. https://doi.org/10.3390/ani10010060