Variation in the Lipin 1 Gene Is Associated with Birth Weight and Selected Carcass Traits in New Zealand Romney Sheep

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sheep Investigated and Data Collection
2.2. Blood Samples and Polymerase Chain Reaction (PCR) Amplification
2.3. Screening for Sequence Variation
2.4. Sequencing of the Variants and Sequence Analysis
2.5. Statistical Analyses
3. Results
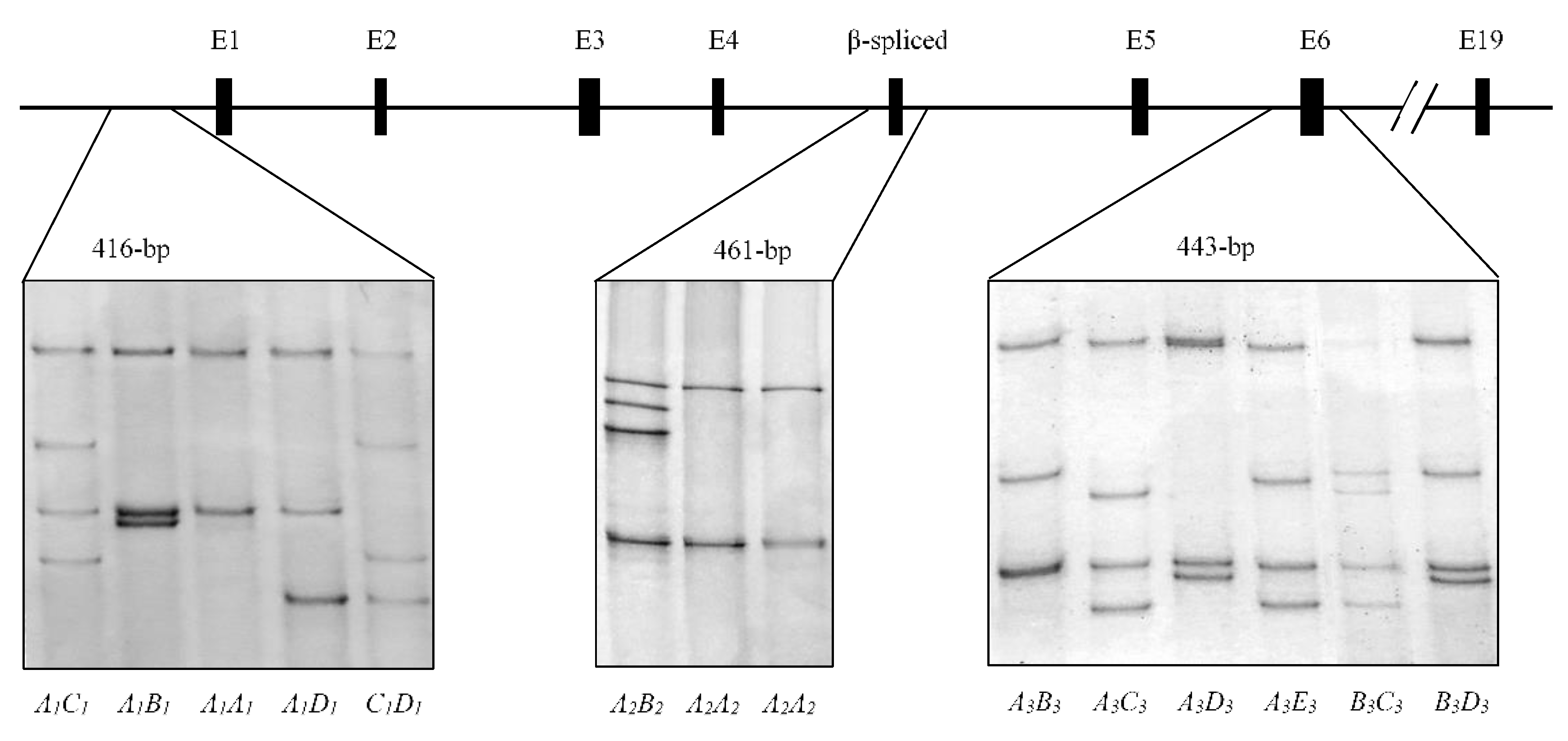
3.1. Variation in Ovine LPIN1
3.2. Effect of LPIN1 Variation on Production Traits
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Péterfy, M.; Phan, J.; Xu, P.; Reue, K. Lipodystrophy in the fld mouse results from mutation of a new gene encoding a nuclear protein, lipin. Nat. Genet. 2001, 27, 121. [Google Scholar] [CrossRef] [PubMed]
- Donkor, J.; Sariahmetoglu, M.; Dewald, J.; Brindley, D.N.; Reue, K. Three mammalian lipins act as phosphatidate phosphatases with distinct tissue expression patterns. J. Biol. Chem. 2007, 282, 3450–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, Y.K.; Lee, M.Y.; Kim, J.W.; Kim, M.; Moon, J.S.; Lee, Y.J.; Ahn, Y.H.; Kim, K.S. Lipin1 is a key factor for the maturation and maintenance of adipocytes in the regulatory network with CCAAT/enhancer-binding protein α and peroxisome proliferator-activated receptor γ2. J. Biol. Chem. 2008, 283, 34896–34906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, J.; Reue, K. Lipin, a lipodystrophy and obesity gene. Cell Metab. 2005, 1, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Péterfy, M.; Phan, J.; Reue, K. Alternatively spliced lipin isoforms exhibit distinct expression pattern, subcellular localization, and role in adipogenesis. J. Biol. Chem. 2005, 280, 32883–32889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeharia, A.; Shaag, A.; Houtkooper, R.H.; Hindi, T.; de Lonlay, P.; Erez, G.; Hubert, L.; Saada, A.; de Keyzer, Y.; Eshel, G.; et al. Mutations in LPIN1 cause recurrent acute myoglobinuria in childhood. Am. J. Hum. Genet. 2008, 83, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Michot, C.; Hubert, L.; Brivet, M.; De Meirleir, L.; Valayannopoulos, V.; Müller-Felber, W.; Venkateswaran, R.; Ogier, H.; Desguerre, I.; Altuzarra, C.; et al. LPIN1 gene mutations: A major cause of severe rhabdomyolysis in early childhood. Hum. Mutat. 2010, 31, E1564–E1573. [Google Scholar] [CrossRef] [Green Version]
- Suviolahti, E.; Reue, K.; Cantor, R.M.; Phan, J.; Gentile, M.; Naukkarinen, J.; Soro-Paavonen, A.; Oksanen, L.; Kaprio, J.; Rissanen, A.; et al. Cross-species analyses implicate Lipin 1 involvement in human glucose metabolism. Hum. Mol. Genet. 2005, 15, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, K.A.; Grimsey, N.; Loos, R.J.F.; Wheeler, E.; Daly, A.; Soos, M.; Semple, R.; Syddall, H.; Cooper, C.; Siniossoglou, S.; et al. Evaluating the role of LPIN1 variation in insulin resistance, body weight, and human lipodystrophy in UK Populations. Diabetes 2008, 57, 2527–2533. [Google Scholar] [CrossRef] [Green Version]
- He, X.P.; Xu, X.W.; Zhao, S.H.; Fan, B.; Yu, M.; Zhu, M.J.; Li, C.C.; Peng, Z.Z.; Liu, B. Investigation of Lpin1 as a candidate gene for fat deposition in pigs. Mol. Biol. Rep. 2009, 36, 1175–1180. [Google Scholar] [CrossRef]
- Cecchinato, A.; Ribeca, C.; Chessa, S.; Cipolat-Gotet, C.; Maretto, F.; Casellas, J.; Bittante, G. Candidate gene association analysis for milk yield, composition, urea nitrogen and somatic cell scores in Brown Swiss cows. Animal 2014, 8, 1062–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Wang, T.; Lu, W.; Zhang, W.; Chen, W.; Kang, X.; Huang, Y. Three indel variants in chicken LPIN1 exon 6/flanking region are associated with performance and carcass traits. Br. Poult. Sci. 2015, 56, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Yuan, Y.; Liang, R.; Li, Y.; Liu, L.; Sun, D. Genetic effects of LPIN1 polymorphisms on milk production traits in dairy cattle. Genes 2019, 10, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, D.L.; Safari, E.; Thompson, J.M.; Smith, C.R. Video image analysis in the Australian meat industry–precision and accuracy of predicting lean meat yield in lamb carcasses. Meat Sci. 2004, 67, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hickford, J.G.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G.H. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Hickford, J.G. Diversity of the glycine/tyrosine-rich keratin-associated protein 6 gene (KAP6) family in sheep. Mol. Biol. Rep. 2011, 38, 31–35. [Google Scholar] [CrossRef]
- Jacobson, G.N.; Clark, P.L. Quality over quantity: Optimizing co-translational protein folding with non-‘optimal’synonymous codons. Curr. Opin. Struct. Biol. 2016, 38, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Wardrop, I.D. Birth weight, liveweight gain in early life, and subsequent gain in sheep and cattle. Aust. J. Agric. Res. 1968, 19, 837–844. [Google Scholar] [CrossRef]
- McSporran, K.D.; Buchanan, R.; Fielden, E.D. Observations on dystocia in a Romney flock. N. Z. Vet. J. 1977, 25, 247–251. [Google Scholar] [CrossRef]
- Dwyer, C.M.; Bünger, L. Factors affecting dystocia and offspring vigour in different sheep genotypes. Prev. Vet. Med. 2012, 103, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Gourt, J.; Hides, S.; Webb-Ware, J. (Eds.) Reproductive management. In Sheep Farming for Meat and Wool, 1st ed.; CSIRO Publishing: Collingwood, Australia, 2010; pp. 143–160. [Google Scholar]



{kind=link}
{kind=link}
| Gene Region | Primer Sequence (5’–3’) | Amplicon Size (Bp) |
|---|---|---|
| 5′ non-coding region | F: ACAAGGAGAGAACATGGGAG | 416 |
| R: CACACCTCAGCACTGGGTC | ||
| LPIN1β-spliced exon | F: AGCAATTCATTATGGGCCTGC | 461 |
| R: CACATAAGTAATTTGGTTAATGG | ||
| Coding exon 6 | F: GATCCAGTCCTCACCACAC | 443 |
| R: CAAGAGAGATGTCCTGTCTC |
| Trait | Variant Assessed 2 | Other Variants Fitted | Mean ± SE 3 | p-Value | |
|---|---|---|---|---|---|
| Absent | Present | ||||
| Birth weight (kg) | A3 | None | 5.8 ± 0.11 | 5.9 ± 0.13 | 0.170 |
| B3 | None | 5.8 ± 0.11 | 6.0 ± 0.14 | 0.061 | |
| C3 | None | 5.9 ± 0.11 | 5.8 ± 0.13 | 0.766 | |
| D3 | None | 5.8 ± 0.10 | 5.8 ± 0.15 | 0.872 | |
| E3 | None | 6.0 ± 0.11 | 5.6 ± 0.13 | 0.001 | |
| B3 | A3, E3 | 5.8 ± 0.10 | 6.0 ± 0.15 | 0.115 | |
| E3 | A3, B3 | 6.1 ± 0.11 | 5.7 ± 0.14 | 0.005 | |
| Pre-weaning growth rate (g/day) | A3 | None | 335.6 ± 6.02 | 333.8 ± 6.48 | 0.760 |
| B3 | None | 330.9 ± 5.77 | 344.5 ± 7.20 | 0.041 | |
| C3 | None | 334.8 ± 5.97 | 334.9 ± 6.59 | 0.984 | |
| D3 | None | 335.6 ± 5.56 | 329.5 ± 8.20 | 0.379 | |
| E3 | None | 334.9 ± 5.99 | 334.8 ± 6.83 | 0.997 | |
| Hot carcass weight (kg) | A3 | None | 17.4 ± 0.28 | 17.5 ± 0.30 | 0.547 |
| B3 | None | 17.4 ± 0.27 | 17.6 ± 0.33 | 0.308 | |
| C3 | None | 17.4 ± 0.28 | 17.4 ± 0.30 | 0.913 | |
| D3 | None | 17.5 ± 0.26 | 17.2 ± 0.37 | 0.329 | |
| E3 | None | 17.2 ± 0.28 | 17.7 ± 0.29 | 0.032 | |
| V-GR (mm) 1 | A3 | None | 7.9 ± 0.40 | 7.7 ± 0.44 | 0.605 |
| B3 | None | 7.8 ± 0.39 | 8.0 ± 0.48 | 0.535 | |
| C3 | None | 7.9 ± 0.40 | 7.8 ± 0.43 | 0.806 | |
| D3 | None | 7.8 ± 0.38 | 7.8 ± 0.54 | 0.948 | |
| E3 | None | 7.5 ± 0.41 | 8.2 ± 0.42 | 0.050 | |
| Leg yield (%) | A3 | None | 22.2 ± 0.18 | 22.2 ± 0.20 | 0.937 |
| B3 | None | 22.2 ± 0.17 | 22.2 ± 0.22 | 0.674 | |
| C3 | None | 22.1 ± 0.18 | 22.3 ± 0.20 | 0.304 | |
| D3 | None | 22.2 ± 0.17 | 21.9 ± 0.24 | 0.110 | |
| E3 | None | 22.2 ± 0.19 | 22.1 ± 0.19 | 0.540 | |
| Loin yield (%) | A3 | None | 15.0 ± 0.13 | 15.1 ± 0.14 | 0.523 |
| B3 | None | 15.0 ± 0.12 | 15.2 ± 0.16 | 0.402 | |
| C3 | None | 15.1 ± 0.13 | 15.1 ± 0.14 | 0.982 | |
| D3 | None | 15.1 ± 0.12 | 14.9 ± 0.17 | 0.257 | |
| E3 | None | 15.0 ± 0.13 | 15.2 ± 0.14 | 0.243 | |
| Shoulder yield (%) | A3 | None | 17.5 ± 0.15 | 17.3 ± 0.15 | 0.137 |
| B3 | None | 17.4 ± 0.14 | 17.3 ± 0.17 | 0.213 | |
| C3 | None | 17.2 ± 0.15 | 17.6 ± 0.15 | 0.005 | |
| D3 | None | 17.4 ± 0.13 | 17.2 ± 0.20 | 0.215 | |
| E3 | None | 17.3 ± 0.15 | 17.5 ± 0.15 | 0.158 | |
| C3 | A3, E3 | 17.3 ± 0.13 | 17.8 ± 0.14 | <0.001 | |
| Total lean meat yield (%) | A3 | None | 54.7 ± 0.33 | 54.6 ± 0.38 | 0.872 |
| B3 | None | 54.6 ± 0.32 | 54.7 ± 0.43 | 0.834 | |
| C3 | None | 54.5 ± 0.34 | 55.0 ± 0.37 | 0.142 | |
| D3 | None | 54.8 ± 0.32 | 54.2 ± 0.45 | 0.129 | |
| E3 | None | 54.6 ± 0.36 | 54.7 ± 0.36 | 0.718 | |
| Proportion of leg yield (%) | A3 | None | 40.6 ± 0.15 | 40.7 ± 0.17 | 0.747 |
| B3 | None | 40.6 ± 0.15 | 40.7 ± 0.18 | 0.586 | |
| C3 | None | 40.7 ± 0.15 | 40.6 ± 0.16 | 0.626 | |
| D3 | None | 40.7 ± 0.14 | 40.6 ± 0.20 | 0.670 | |
| E3 | None | 40.8 ± 0.16 | 40.5 ± 0.16 | 0.045 | |
| Proportion of loin yield (%) | A3 | None | 27.6 ± 0.12 | 27.7 ± 0.13 | 0.167 |
| B3 | None | 27.6 ± 0.12 | 27.7 ± 0.15 | 0.226 | |
| C3 | None | 27.7 ± 0.12 | 27.5 ± 0.13 | 0.029 | |
| D3 | None | 27.6 ± 0.11 | 27.7 ± 0.16 | 0.816 | |
| E3 | None | 27.5 ± 0.13 | 27.7 ± 0.13 | 0.069 | |
| C3 | A3, E3 | 27.5 ± 0.11 | 27.4 ± 0.13 | 0.414 | |
| E3 | A3, C3 | 27.3 ± 0.11 | 27.6 ± 0.13 | 0.014 | |
| Proportion of shoulder yield (%) | A3 | None | 32.0 ± 0.19 | 31.8 ± 0.18 | 0.260 |
| B3 | None | 31.9 ± 0.17 | 31.6 ± 0.21 | 0.087 | |
| C3 | None | 31.8 ± 0.18 | 32.0 ± 0.18 | 0.124 | |
| D3 | None | 31.9 ± 0.17 | 32.0 ± 0.25 | 0.507 | |
| E3 | None | 31.8 ± 0.19 | 31.9 ± 0.18 | 0.438 | |
| B3 | C3 | 32.0 ± 0.17 | 31.9 ± 0.21 | 0.545 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Zhou, H.; Gong, H.; He, J.; Luo, Y.; Hickford, J.G.H.; Hu, J.; Wang, J.; Liu, X.; Li, S. Variation in the Lipin 1 Gene Is Associated with Birth Weight and Selected Carcass Traits in New Zealand Romney Sheep. Animals 2020, 10, 237. https://doi.org/10.3390/ani10020237
Wang G, Zhou H, Gong H, He J, Luo Y, Hickford JGH, Hu J, Wang J, Liu X, Li S. Variation in the Lipin 1 Gene Is Associated with Birth Weight and Selected Carcass Traits in New Zealand Romney Sheep. Animals. 2020; 10(2):237. https://doi.org/10.3390/ani10020237
Chicago/Turabian StyleWang, Guan, Huitong Zhou, Hua Gong, Jianning He, Yuzhu Luo, Jon G. H. Hickford, Jiang Hu, Jiqing Wang, Xiu Liu, and Shaobin Li. 2020. "Variation in the Lipin 1 Gene Is Associated with Birth Weight and Selected Carcass Traits in New Zealand Romney Sheep" Animals 10, no. 2: 237. https://doi.org/10.3390/ani10020237
APA StyleWang, G., Zhou, H., Gong, H., He, J., Luo, Y., Hickford, J. G. H., Hu, J., Wang, J., Liu, X., & Li, S. (2020). Variation in the Lipin 1 Gene Is Associated with Birth Weight and Selected Carcass Traits in New Zealand Romney Sheep. Animals, 10(2), 237. https://doi.org/10.3390/ani10020237