Minimal Effects of Rearing Enrichments on Pullet Behaviour and Welfare



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
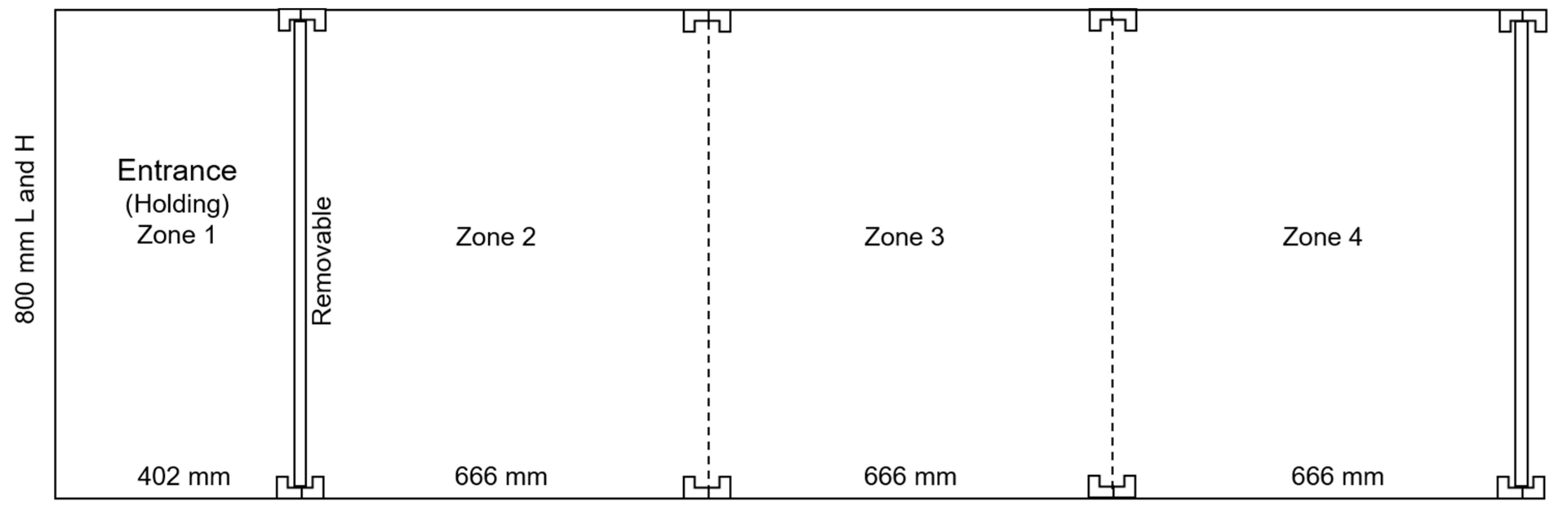
2.2. Animals and Pullet Housing
2.3. Data Collection Procedures
2.3.1. Body Weight
2.3.2. Novel Arena Test
2.3.3. Manual Restraint
2.3.4. Organ Weights
2.3.5. Blood Samples
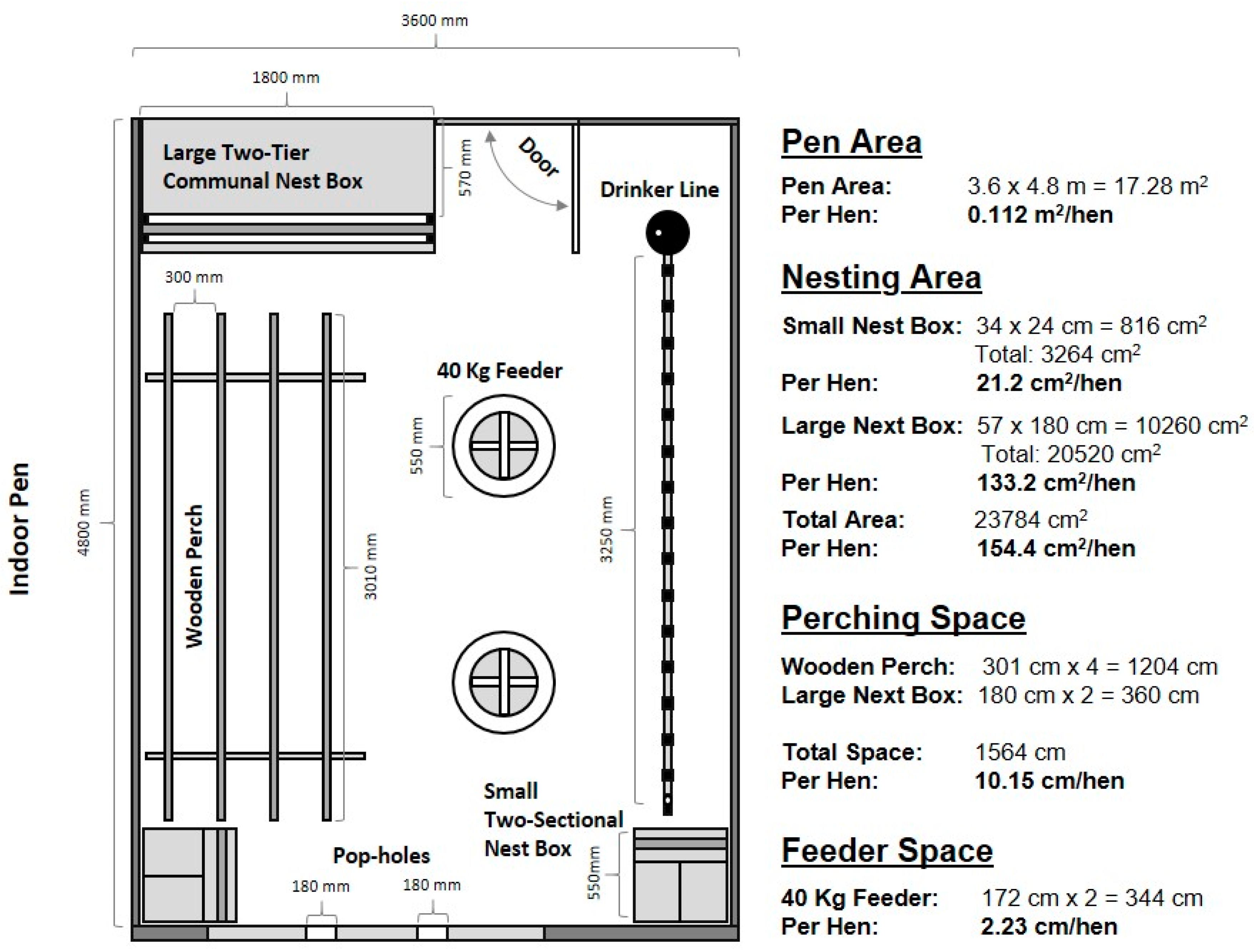
2.3.6. Layer Housing and Video Recording
2.4. Data and Statistical Analyses
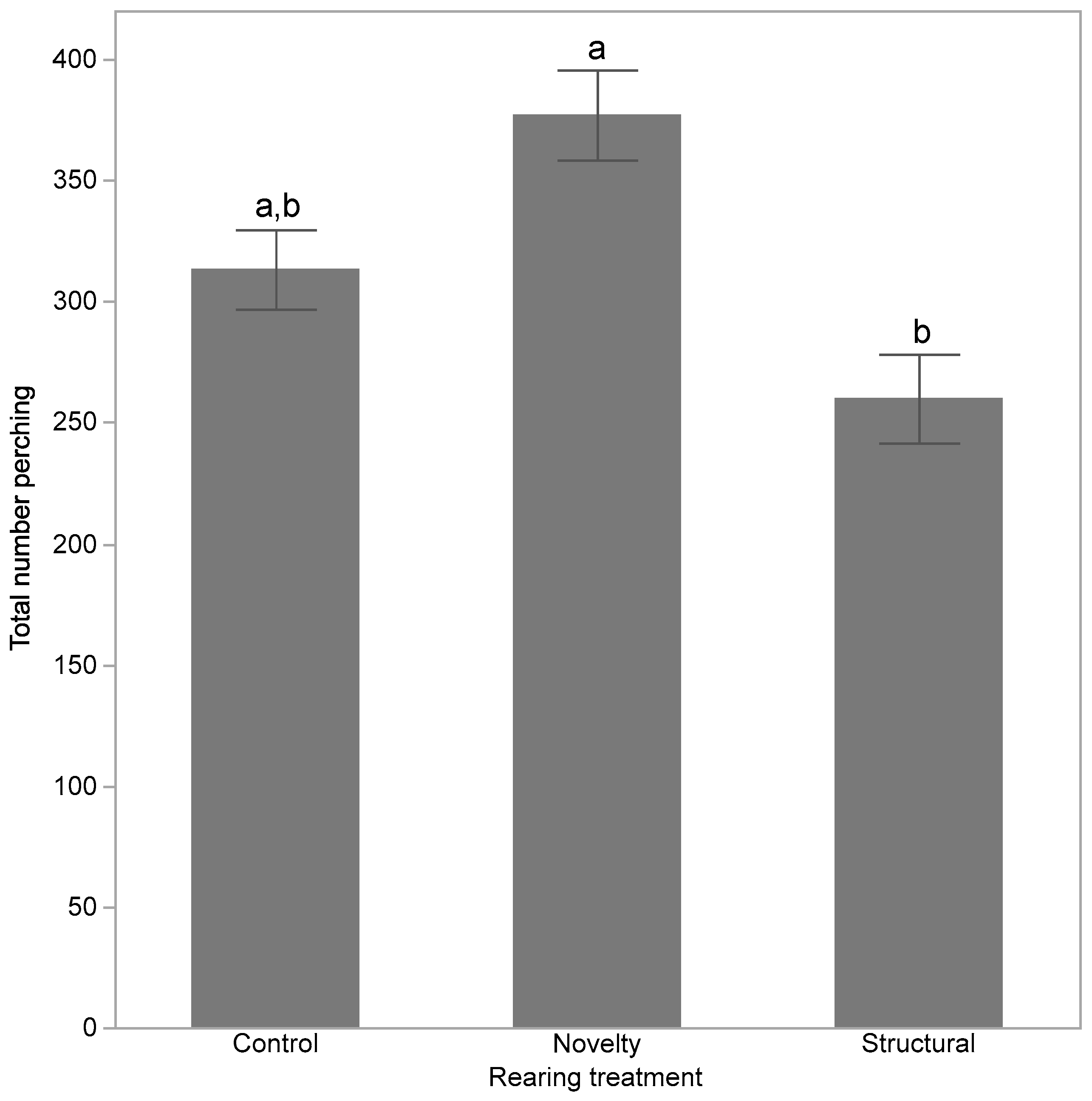
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campbell, D.L.M.; de Haas, E.N.; Lee, C. A review of environmental enrichment for laying hens during rearing in relation to their behavioral and physiological development. Poult. Sci. 2019, 98, 9–28. [Google Scholar] [CrossRef] [PubMed]
- de Haas, E.N.; Bolhuis, J.E.; Kemp, B.; Groothuis, T.G.G.; Rodenburg, T.B. Parents and early life environment affect behavioral development of laying hen chickens. PLoS ONE 2014, 9, e90577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regmi, P.; Deland, T.S.; Steibel, J.P.; Robison, C.I.; Haut, R.C.; Orth, M.W.; Karcher, D.M. Effect of rearing environment on bone growth of pullets. Poult. Sci. 2015, 94, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Matur, E.; Eraslan, E.; Akyazi, I.; Ergul Ekiz, E.; Eseceli, H.; Keten, H.; Metiner, K.; Aktaran Bala, D. The effect of furnished cages on the immune response of laying hens under social stress. Poult. Sci. 2015, 94, 2853–2862. [Google Scholar] [CrossRef]
- Matur, E.; Akyazi, I.; Eraslan, E.; Ergulekiz, E.; Eseceli, H.; Keten, M.; Metiner, K.; Aktaran Bala, D. The effects of environmental enrichment and transport stress on the weights of lymphoid organs, cell-mediated immune response, heterophil functions and antibody production in laying hens. Anim. Sci. J. 2016, 87, 284–292. [Google Scholar] [CrossRef]
- Janczak, A.M.; Riber, A.B. Review of rearing-related factors affecting the welfare of laying hens. Poult. Sci. 2015, 94, 1454–1469. [Google Scholar] [CrossRef]
- Colson, S.; Arnould, C.; Michel, V. Influence of rearing conditions of pullets on space use and performance of hens placed in aviaries at the beginning of the laying period. Appl. Anim. Behav. Sci. 2008, 111, 286–300. [Google Scholar] [CrossRef]
- Grigor, P.N.; Hughes, B.O.; Appleby, M.C. Effects of regular handling and exposure to an outside area on subsequent fearfulness and dispersal in domestic hens. Appl. Anim. Behav. Sci. 1995, 44, 47–55. [Google Scholar] [CrossRef]
- Newberry, R.C. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Dávila, S.G.; Campo, J.L.; Gil, M.G.; Prieto, M.T.; Torres, O. Effects of auditory and physical enrichment on 3 measurements of fear and stress (tonic immobility duration, heterophil to lymphocyte ratio, and fluctuating asymmetry) in several breeds of layer chicks. Poult. Sci. 2011, 90, 2459–2466. [Google Scholar] [CrossRef]
- Hartcher, K.M.; Tran, M.K.T.N.; Wilkinson, S.J.; Hemsworth, P.H.; Thomson, P.C.; Cronin, G.M. Plumage damage in free-range laying hens: Behavioural characteristics in the rearing period and the effects of environmental enrichment and break-trimming. Appl. Anim. Behav. Sci. 2015, 164, 64–72. [Google Scholar] [CrossRef]
- Brantsæter, M.; Tahamtani, F.M.; Moe, R.O.; Hansen, T.B.; Orritt, R.; Nicol, C.; Janczak, A.M. Rearing laying hens in aviaries reduces fearfulness following transfer to furnished cages. Front. Vet. Sci. 2016, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Brantsæter, M.; Nordgreen, J.; Rodenburg, T.B.; Tahamtani, F.M.; Popova, A.; Janczak, A.M. Exposure to increased environmental complexity during rearing reduces fearfulness and increases use of three-dimensional space in laying hens (Gallus gallus domesticus). Front. Vet. Sci. 2016, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Primary Industries Standing Committee. Model Code of Practice for the Welfare of Animals: Domestic Poultry; CSIRO PUBLISHING: Victoria, Australia, 2002. [Google Scholar]
- Hy-Line® 2016. Hy-Line® Brown Management Guide for Alternative Systems-Australia. Available online: https://www.hyline.com/userdocs/pages/BRN_ALT_COM_AUS.pdf (accessed on 1 November 2017).
- Downing, J.A.; Bryden, W.L. Determination of corticosterone concentrations in egg albumen: A non-invasive indicator of stress in laying hens. Physiol. Behav. 2008, 95, 381–387. [Google Scholar] [CrossRef]
- Gulland, F.M.D.; Hawkey, C.M. Avian hematology. Vet. Annu. 1990, 30, 126–136. [Google Scholar]
- Hocking, P.M.; Channing, C.E.; Waddington, D.; Jones, R.B. Age-related changes in fear, sociality and pecking behaviours in two strains of laying hens. Br. Poult. Sci. 2001, 42, 414–423. [Google Scholar] [CrossRef]
- Jones, R.B.; Waddington, D. Modification of fear in domestic chicks, Gallus gallus domesticus, via regular handling and early environmental enrichment. Anim. Behav. 1992, 43, 1021–1033. [Google Scholar] [CrossRef]
- McKeegan, D.E.F.; Savory, C.J. Behavioural and hormonal changes associated with sexual maturity in layer pullets. Br. Poult. Sci. 1998, 39 (Suppl. 1), 6–7. [Google Scholar] [CrossRef]
- Campbell, D.L.M.; Hinch, G.N.; Downing, J.A.; Lee, C. Fear and coping styles of outdoor-preferring, moderate-outdoor and indoor-preferring free-range laying hens. Appl. Anim. Behav. Sci. 2016, 185, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; de Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping styles in animals: Current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Van der Eijk, J.A.J.; Lammers, A.; Kjaer, J.B.; Rodenburg, T.B. Stress response, peripheral serotonin and natural antibodies in feather pecking genotypes and phenotypes and their relation with coping style. Physiol. Behav. 2019, 199, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korte, S.M.; Beuving, G.; Ruesink, W.; Blokhuis, H.J. Plasma catecholamine and corticosterone levels during manual restraint in chicks from a high and low feather pecking line of laying hens. Physiol. Behav. 1997, 62, 437–441. [Google Scholar] [CrossRef]
- Rodenburg, T.B.; Bolhuis, J.E.; Koopmanschap, R.E.; Ellen, E.D.; Decuypere, E. Maternal care and selection for low mortality affect post-stress corticosterone and peripheral serotonin in laying hens. Physiol. Behav. 2009, 98, 519–525. [Google Scholar] [CrossRef]
- Zidar, J.; Balogh, A.; Favati, A.; Jensen, P.; Leimear, O.; Løvlie, H. A comparison of animal personality and coping styles in the red junglefowl. Anim. Behav. 2017, 130, 209–220. [Google Scholar] [CrossRef]
- Gunnarsson, S.; Yngvesson, J.; Keeling, L.J.; Forkman, B. Rearing without early access to perches impairs the spatial skills of laying hens. Appl. Anim. Behav. Sci. 2000, 67, 217–218. [Google Scholar] [CrossRef]
- Heikkilä, M.; Wichman, A.; Gunnarsson, S.; Valros, A. Development of perching behaviour in chicks reared in enriched environment. Appl. Anim. Behav. Sci. 2006, 99, 145–156. [Google Scholar] [CrossRef]
- Bari, M.S.; Cohen-Barnhouse, A.M.; Campbell, D.L.M. Early enrichments influenced nest use and egg quality in free-range laying hens. Animal 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M.; Tobalske, B.; Martins, C.; Bowley, S.; Wuerbel, H.; Harlander-Matauschek, A. Use of space by domestic chicks housed in complex aviaries. Appl. Anim. Behav. Sci. 2016, 181, 115–121. [Google Scholar] [CrossRef]
- Müller, B.R.; Medeiros, H.A.S.; de Sousa, R.S.; Molento, C.F.M. Chronic welfare restrictions and adrenal gland morphology in broiler chickens. Poult. Sci. 2015, 94, 575–578. [Google Scholar] [CrossRef]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2006, 95, 2208–2215. [Google Scholar] [CrossRef]
- Shini, S.; Shini, A.; Huff, G.R. Effects of chronic and repeated corticosterone administration in rearing chickens on physiology, the onset of lay and egg production of hens. Physiol. Behav. 2009, 98, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Iyasere, O.S.; Beard, A.P.; Guy, J.H.; Bateson, M. Elevated levels of the stress hormone, corticosterone, cause ’pessimistic’ judgment bias in broiler chickens. Sci. Rep. 2017, 7, 6860. [Google Scholar] [CrossRef] [PubMed]
- Puvadolpirod, S.; Thaxton, J.P. Model of physiological stress in chickens 1. Response parameters. Poult. Sci. 2000, 79, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Bari, M.S.; Laurenson, Y.C.S.M.; Cohen-Barnhouse, A.M.; Walkden-Brown, S.W.; Campbell, D.L.M. Effects of outdoor ranging on external and internal health parameters for hens from different rearing enrichments. PeerJ 2020, accepted. [Google Scholar]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Lilliehöök, I.; Wall, H.; Tauson, R.; Tvedten, H. Differential leukocyte counts determined in chicken blood using the Cell-Dyn 3500. Vet. Clin. Pathol. 2004, 33, 133–3138. [Google Scholar] [CrossRef]
- Post, J.; Rebel, J.M.; ter Huurne, A.A. Automated blood cell count: A sensitive and reliable method to study corticosterone-related stress in broilers. Poult. Sci. 2003, 82, 591–595. [Google Scholar] [CrossRef]
- Ruhnke, I.; Andronicos, N.M.; Swick, R.A.; Hine, B.; Sharma, N.; Kheravii, S.K.; Hunt, P. Immune responses following experimental infection with Ascaridia galli and necrotic enteritis in broiler chickens. Avian Pathol. 2017, 46, 602–609. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Body Weight (kg) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | 5 Weeks | n | 8 Weeks | n | 12 Weeks | n | 16 Weeks | n |
| Control | 0.35 ± 0.004 | 15 | 0.70 ± 0.006 d | 567 | 1.12 ± 0.006 b,c | 548 | 1.37 ± 0.006 a | 532 |
| Novelty | 0.35 ± 0.004 | 15 | 0.69 ± 0.006 d | 551 | 1.09 ± 0.006 c | 547 | 1.36 ± 0.006 a | 519 |
| Structural | 0.35 ± 0.004 | 15 | 0.79 ± 0.006 d | 569 | 1.13 ± 0.006 b | 570 | 1.38 ± 0.006 a | 539 |
| Treatment | % Liver Wgt | % Spleen Wgt | % Bursa Wgt | % Thymus Wgt | % Adrenal Wgt |
|---|---|---|---|---|---|
| Control | 1.77 ± 0.03 | 0.16 ± 0.03 | 0.29 ± 0.01 | 0.21 ± 0.009 | 0.029 ± 0.0001 b |
| Novelty | 1.71 ± 0.03 | 0.16 ± 0.03 | 0.27 ± 0.01 | 0.21 ± 0.008 | 0.031 ± 0.001 b |
| Structural | 1.72 ± 0.03 | 0.16 ± 0.004 | 0.29 ± 0.02 | 0.21 ± 0.008 | 0.034 ± 0.001 a |
| Treatment | Latency Struggle (s) | Latency Vocalise (s) | Number of Struggles | Number of Vocals | Corticosterone ng/mL |
|---|---|---|---|---|---|
| Control | 163.47 ± 18.96 | 31.87 ± 12.66 | 3.9 ± 1.30 | 58.69 ± 6.84 | 3.76 ± 0.28 |
| Novelty | 156.33 ± 19.53 | 32.37 ± 10.78 | 3.47 ± 0.70 | 71.66 ± 8.08 | 3.96 ± 0.34 |
| Structural | 190.07 ± 18.53 | 54.17 ± 16.68 | 2.47 ± 0.58 | 63.67 ± 13.17 | 3.46 ± 0.16 |
| Variable | Transport | Control | Novelty | Structural |
|---|---|---|---|---|
| Heterophils 109/L | Before | 8.05 ± 0.43 | 8.58 ± 0.43 | 9.06 ± 0.43 |
| After | 8.57 ± 0.43 | 8.18 ± 0.43 | 8.50 ± 0.43 | |
| Lymphocytes 109/L | Before | 21.91 ± 2.03 | 19.62 ± 2.03 | 15.48 ± 2.03 |
| After * | 22.34 ± 2.03 | 23.40 ± 2.03 | 21.35 ± 2.03 | |
| H/L ratio | Before | 0.52 ± 0.21 a,b | 0.73 ± 0.21 a,b | 1.48 ± 0.21 a |
| After | 0.70 ± 0.21 a,b | 0.41 ± 0.21 b | 0.51 ± 0.21 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, D.L.M.; Gerber, P.F.; Downing, J.A.; Lee, C. Minimal Effects of Rearing Enrichments on Pullet Behaviour and Welfare. Animals 2020, 10, 314. https://doi.org/10.3390/ani10020314
Campbell DLM, Gerber PF, Downing JA, Lee C. Minimal Effects of Rearing Enrichments on Pullet Behaviour and Welfare. Animals. 2020; 10(2):314. https://doi.org/10.3390/ani10020314
Chicago/Turabian StyleCampbell, Dana L.M., Priscilla F. Gerber, Jeff A. Downing, and Caroline Lee. 2020. "Minimal Effects of Rearing Enrichments on Pullet Behaviour and Welfare" Animals 10, no. 2: 314. https://doi.org/10.3390/ani10020314
APA StyleCampbell, D. L. M., Gerber, P. F., Downing, J. A., & Lee, C. (2020). Minimal Effects of Rearing Enrichments on Pullet Behaviour and Welfare. Animals, 10(2), 314. https://doi.org/10.3390/ani10020314