2. Materials and Methods
The experiment was carried out at the experimental station of the Institute of Animal Nutrition, Friedrich-Loeffler-Institut (FLI), in Braunschweig, Germany, in accordance with the German Animal Welfare Act approved by the LAVES (Lower Saxony Office for Consumer Protection and Food Safety, Germany) (AZ33.19-42502-04-16/2378).
2.1. Experimental Design
The experiment started individually for each cow with the expected day 42 ante partum (ap) and ended at day 110 pp. A total of 59 pluriparous German Holstein dairy cows, including eight rumen- and duodenum-cannulated cows, were assigned to two groups, a control (CON, n = 30) and an L-carnitine group (CAR, n = 29), balanced for numbers of lactation (2–5 lactations), body weight (568–1008 kg), body condition score (2.5–4.75) and fat-corrected milk yield of previous lactation. To circumvent ruminal degradation, the cows in CAR received 125 g of a rumen-protected L-carnitine product (Carneon 20 Rumin-Pro, Kaesler Nutrition GmbH, Cuxhaven, Germany) per cow and day, which was included in the concentrate feed. This amount corresponded to a daily L-carnitine intake of 25 g per cow and day. To balance the fat content of the L-carnitine product, CON obtained an equivalent fat product (BergaFat F-100 HP, Berg+Schmidt GmbH & Co. KG, Hamburg, Germany) as used for the rumen protection of the L-carnitine. The cows were kept in a free-stall barn with slatted floors and cubicles with rubber pads and were rehoused for calving in the calving pen, where a maximum of two cows were kept in one straw bedding box.
Both groups were fed with a partial mixed ration (PMR). Whereas the composition of roughage remained unchanged during the whole trial (70% maize silage and 30% grass silage), the proportion of roughage to concentrate was variable in accordance with the recommendation of nutrient and energy supply of the Society of Nutrition Physiology (GfE). Initially, up to day 1
ap, diets of 80% roughage and 20% concentrate were fed. The amount of concentrate was increased from 30% to 50% up to 14 days
pp and from then on, 50% concentrate was constantly fed up to day 110
pp. The PMR was offered by feed-weigh troughs (Roughage Intake Control, System Insentec B.V., Marknesse, The Netherlands) and the supplementary, restricted, pelleted concentrate was provided via concentrate feeding stations (Insentec B.V., Marknesse, The Netherlands). Water was offered for ad libitum intake. The components and the chemical composition of roughages and concentrate feed are shown in
Table 1.
2.2. Measurements and Sample Collection
Samples of the PMR were taken daily, whereas the concentrate feed was sampled once a week. The feed samples were pooled over four-week periods. Cows were equipped with ear transponders to enable the individual recording of feed and water intake.
Before calving, the body weight (BW) was recorded once a week, and during the lactation period, twice a day with a scale situated between the milking parlour and barn. Once a week, the same person also evaluated the body condition score (BCS) according to Edmonson et al. [
14] (5-point scale, 1 = lean, 5 = obese). The cows were milked at 5:30 am and 3:30 pm. During milking, the milk yield was recorded by a milk counter (Lemmer Fullwood GmbH, Lohmar, Germany). Milk samples were taken twice a week during morning and evening milking to analyse milk ingredients.
Blood samples were collected from
Vena jugularis externa by needle puncture or for frequent sampling,
pp by indwelling catheters (from 0.5 h until 12 h after calving). Prior to blood collection, 20 mL of blood were aspirated and discarded from a catheter which was flushed with 20 mL of 0.9% saline solution after the collection. Blood samples were taken at the following time points: day 42, 14, 7, 3 and 1
ap, 0.5, 1, 2, 3, 4, 6, 9, 12, 24, 48 and 72 h
pp, day 7, 14, 21, 28, 42, 56, 100 and 110
pp. Furthermore, ultrasonic measurements according to Raschka et al. [
15] were performed on day 42
ap, 72 h
pp, day 42
pp and day 100
pp to estimate masses of adipose tissues (subcutaneous (SAT), retroperitoneal (RAT), omental (OAT), mesenteric adipose tissue (MAT)) by using a Mindray M5 Vet (Mindray, Shenzhen, China) diagnostic ultrasound system with a linear (6 MHz, Mindray 6LE5Vs) and a convex probe (3 MHz, Mindray 3C5s).
2.3. Analyses
In accordance with the methods of the Association of German Agricultural Analytic and Research Institutes (VDLUFA) [
16], the pooled feed samples were analysed for dry matter (method number 3.1), crude ash (method number 8.1), crude protein (method number 4.1.2), ether extract (method number 5.1.1), crude fibre (method number 6.1), neutral detergent fibre without ash (NDFom, method number 6.5.1) and acid detergent fibre without ash (ADFom, method number 6.5.2).
Milk ingredients (fat, protein, lactose and urea) were analysed by an infrared milk analyser (Milkoscan FT 6000®, Foss Electric, Hillerød, Denmark) and somatic cell count (SCC) was determined by a flow cytometric measurement (Fossomatic 500®, Foss Electric, Hillerød, Denmark).
For analyses of NEFA, BHB and TG, serum tubes were incubated for 30 min at 30 °C and the serum was separated via centrifugation of blood samples for 15 min at 15 °C and 3000 g (Varifuge 3.0, Heraeus, Hanau, Germany). The extracted serum was kept at a temperature of −80 °C until the day of measurement. Analyses were performed by a photometric method using the Eurolyser CCA 180 (Eurolyser Diagnostica GmbH, Salzburg, Austria). The insulin concentration in serum was determined by a sandwich-enzyme-linked immunosorbent assay (ELISA) (Bovine Insulin ELISA, 10-1201-01, Mecordia AB, Uppsala, Sweden) according to the manufacturer’s protocol.
Furthermore, an automated blood gas and electrolyte analyser (GEM Premier 4000, Werfen, Kirchheim, Germany) was used for determination of sodium, potassium, chloride, hydrogen carbonate and calcium ions, glucose and lactate concentration, the temperature-corrected partial pressure of oxygen and carbon dioxide (TpO
2, TpCO
2), oxygen saturation (sO
2), total carbon dioxide, temperature-corrected pH, base excess (BE) and base excess in extra cellular fluid (Beecf) in heparinized blood which was collected in sample syringes (Werfen, Kirchheim, Germany). Total calcium and total phosphorous were measured spectrometrically in blood serum (Calcium AS III Arsenazo III colour test/Phosphorus (mono) ultraviolet (UV)-method kit, Greiner Diagnostic GmbH, Bahlingen am Kaiserstuhl, Germany). The concentrations of N
Ƹ-trimethyllysine (TML), y-butyrobetaine (yBB), acetylcarnitine (ACA) and free carnitine (CA) were analysed in ethylenediamine tetraacetic acid (EDTA) plasma by a tandem mass spectrometry method according to Hirche et al. [
17].
2.4. Clinical Examinations and Assessment of Health Status
Each cow was frequently clinically examined from 0.5 up to 72 h after calving according to the methods of Dirksen et al. [
18]. To evaluate the collected clinical findings, a cumulative clinical score was created, in which the physiological level was defined as 0 and deviations with numbers increasing with severity (see
Table A1,
Appendix A). For example, a wet muzzle was regarded as physiological and received the score of 0, while a score of 1 was assigned to a dry muzzle. Clinical variables which might differ into both directions from the physiological range were treated differently. For example, a score of 0 was assigned to the physiological range of heart rate (65 to 85 beats per minute, bpm) while a deviation in direction of bradycardia or tachycardia was scored with 1, 2 or 3. Finally, the individual scores were cumulated and related to the maximum possible score of 29. Thus, a score of 29 represented a cow which deviated at maximum from the physiological value for each individual clinical parameter while a score of 0 indicated a clinically inconspicuous cow.
2.5. Calculations
The fat-corrected milk (FCM) was calculated according to Gaines et al. [
19]:
Energy-corrected milk (ECM), milk energy, net energy requirement for maintenance (NEM) and net energy requirement for lactation (NEL) were calculated according to the Society of Nutrition Physiology (GfE) [
11]:
Net energy balance (NEB) was calculated by using the following equation:
Gestational requirements were taken into account by addition of 13 MJ NEL/d for week six up to week 3 ap and of 18 MJ NEL/d for the last 3 weeks ap.
According to Hurley et al. [
20], the feed efficiency (FE) corresponds to the ratio of ECM and dry matter intake (DMI) (kg) and the residual energy intake (REI) was calculated by using the following formula:
To calculate the EEI, a non-linear regression model was used, whereby significant variables and related coefficients were determined:
The Revised Quantitative Insulin Sensitivity Check Index (RQUICKI) was calculated according to Caloni et al. [
21]:
Anion gap was calculated by using following equation according to Brouwer et al. [
22]:
The mass of SAT, RAT, OAT and MAT was calculated according to Raschka et al. [
15] in consideration of different anatomic locations for ultra-sonographic measurements: twelfth rib at the level of the greater
trochanter (R12), centre of
paralumbar fossa from skin to
peritoneum (AW3c), intertransverse space cranial to lumbar, intertransverse space where caudal pole of kidney is visible from skin to kidney margin away from transducer (KD3b), back fat thickness (BFT), centre of
paralumbar fossa from skin to muscle margin away from transducer (AW3b), point of interception of a vertical line through the last
lumbar vertebra and a horizontal line through the greater
trochanter from skin to muscle margin away from transducer (AW1b), intertransverse space cranial to lumbar intertransverse space where caudal pole of kidney is visible from skin to
peritoneum (KD2c).
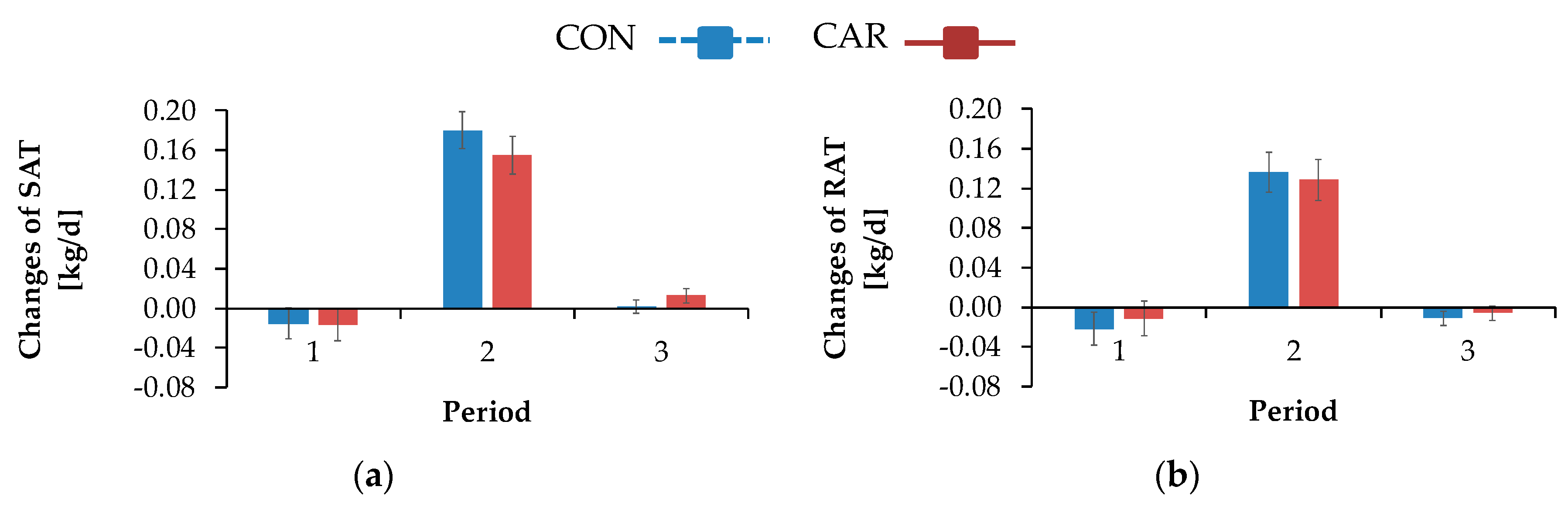
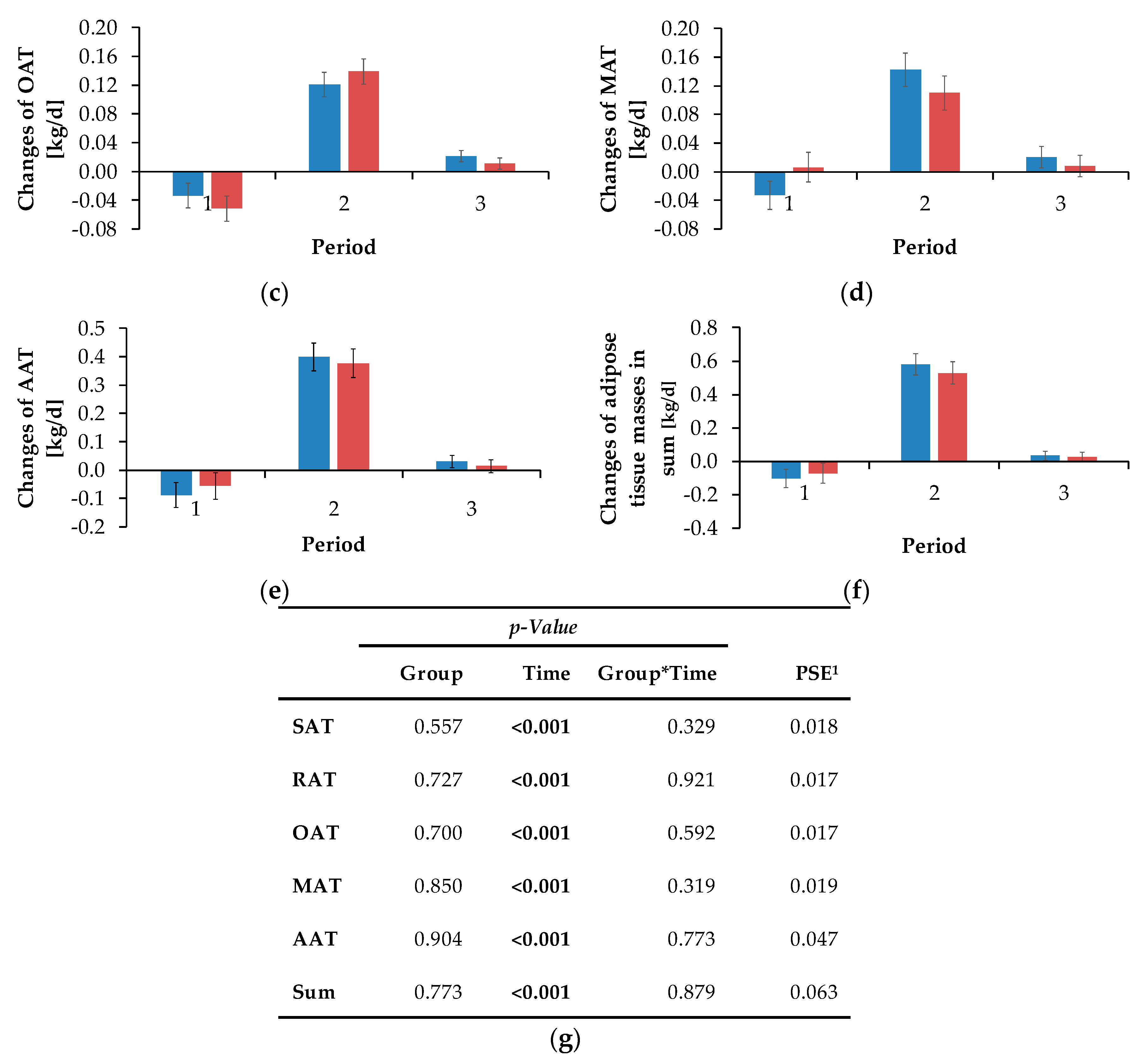
The sum of the masses of OAT, MAT and RAT results in the abdominal adipose tissue mass (AAT). Additionally, the changes in adipose tissue masses (FM (kg/d)) were calculated by the differences between the different time points, whereby negative values indicated a gain and positive values indicated a reduction of masses in the particular localisation. The corresponding release of energy (considering that 1 g fat results in 39.8 kJ gross energy [
23], whereby 16% of them is lost as heat [
24]) was calculated as follows:
2.6. Statistical Analyses
Before statistical analyses were carried out, weekly mean values were calculated for performance and feeding data. Analyses were performed using the MIXED procedure of SAS software (SAS Enterprise Guide 6.1, SAS Institute Inc., Cary, NC, USA) with the restricted maximum likelihood method (reml). The model included the group (G, CON or CAR), the time (T) and the interaction between G and T as fixed factors. For blood variables, the values of day 42 ap were considered as covariables. The choice of covariance structure (compound symmetry, autoregressive or unstructured) for the respective parameter were based on the smallest Akaike information criterion for a finite sample size (AICC). Effects were assumed as significant when p-values were equal or smaller than 0.05. Least Squares (LS) Mean comparisons were carried out with the Tukey–Kramer test.
The relationships between REI and NEB, NEB and energy release from adipose tissue were evaluated by linear regressions for each experimental group. Additionally, linear regressions for the relationship between REI and NEB for cows in negative NEB and positive NEB (independent of treatment groups) were carried out and slopes were compared. All results are given as LS Means and additionally, the standard deviation is listed in the text. In tables and figures, the pooled standard error (PSE) of LS Means over both groups and all time points is indicated.
4. Discussion
The aim of the present study was to investigate the hypothesis that L-carnitine supplementation improves the energy status of the cow around calving with consequences on the energy-consuming puerperium, including the recovery from calving and the magnitude of negative energy balance.
In doing so, clinical signs, milking performance, several indicators of energy metabolism including energy balance, adipose tissue dynamics, blood clinical-chemical traits and systemic carnitine levels, and electrolyte balance were evaluated with particular consideration of the first 72 h pp.
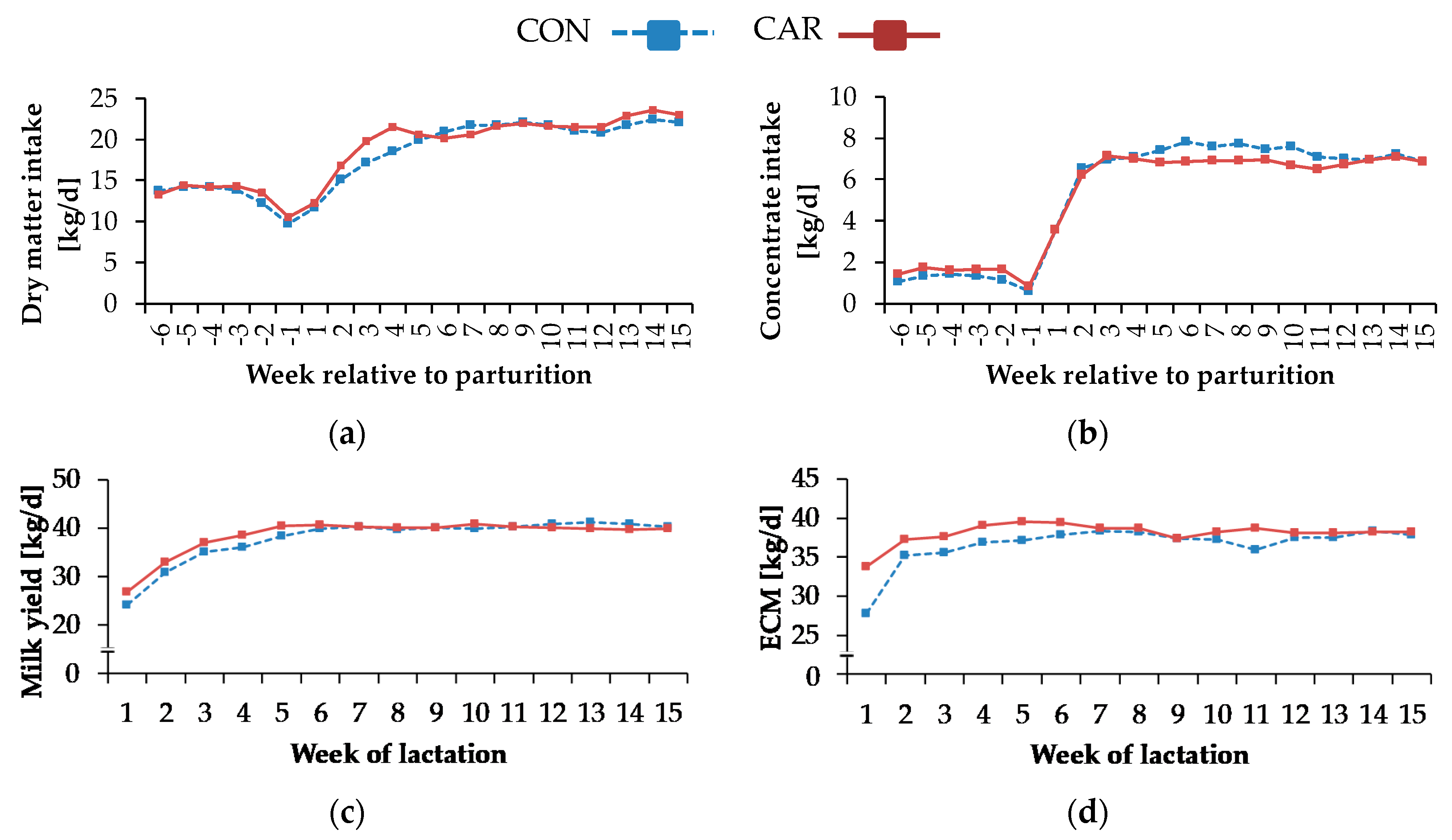
As rations were designed on an isoenergetic basis and DMI remained unaffected by the L-carnitine supplementation, the resulting NEL intake followed a similar pattern. Carlson et al. [
6] reported that DMI did not differ between non-supplemented cows and cows receiving 6 or 50 g of non-rumen-protected L-carnitine per day, whereas at a dosage of 100 g dietary L-carnitine per day, DMI was lower during the first two weeks of lactation. Based on these findings and on the present results, we conclude that a moderate L-carnitine supplementation neither stimulates nor depresses feed intake of transition cows.
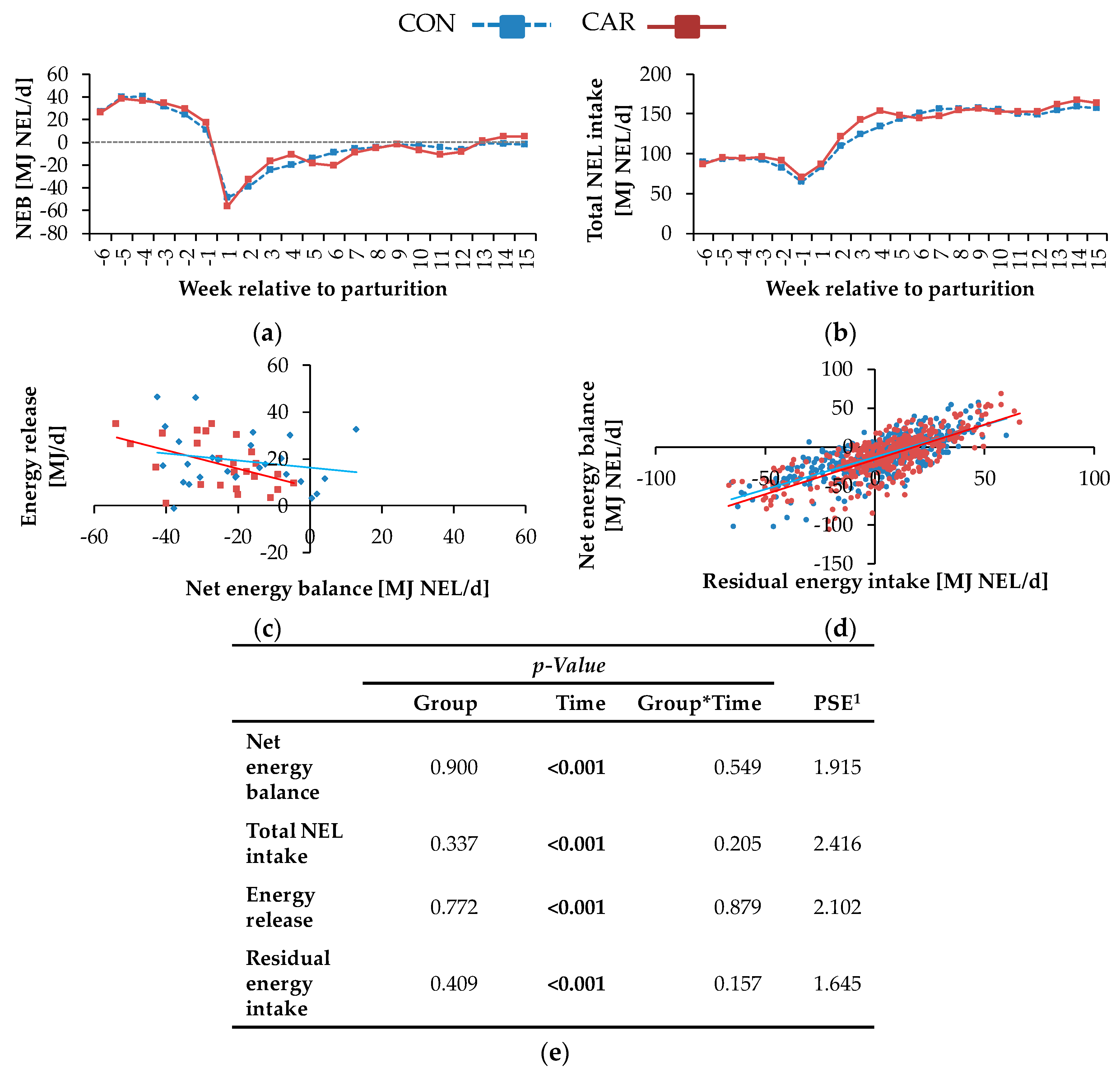
The REI represented the deviation of the EEI from total NEL intake, that could not be explained by the corresponding regression equation (
Figure 3d). An individual positive REI represents a cow consuming more energy than expected and which is consequently less efficient compared to a cow showing a negative REI. As L-carnitine supplementation did not influence the REI, it might be concluded that energetic efficiency was not improved. Independent of dietary treatment, the NEB increased linearly and simultaneously with REI, suggesting the energetic efficiency to decrease with an increase in NEB. Here, cows in a negative NEB were as efficient as cows in a positive NEB, suggesting that efficiency during mobilisation is not different compared to periods of positive NEB. While the REI represented an estimate for the overall energetic efficiency, the contribution to the NEB by the energy mobilised from adipose tissues gave an indication for the energetic efficiency of lipomobilisation.
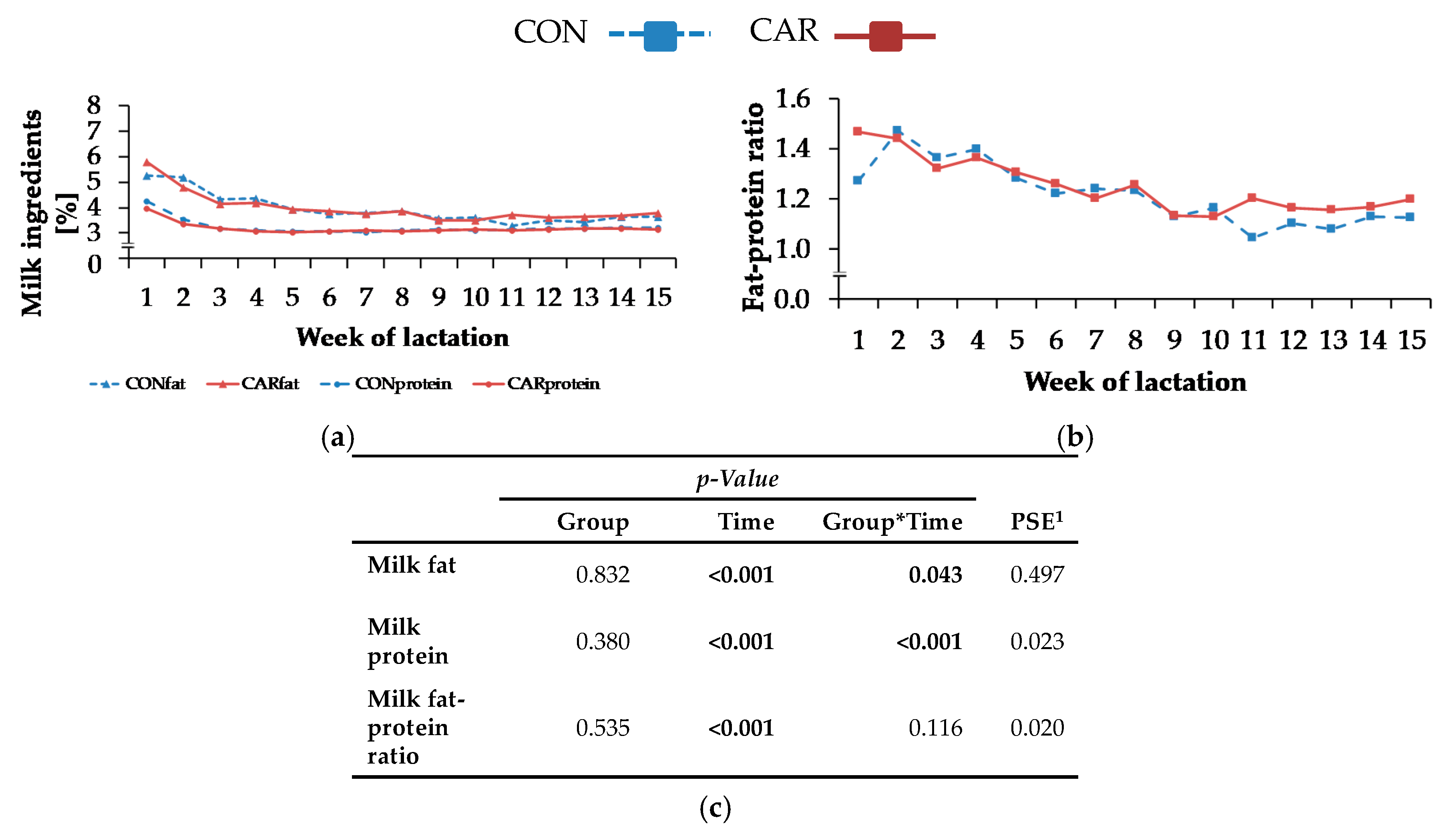
The reason for the initially higher milk energy of L-carnitine-supplemented cows might be the simultaneously increased milk fat at this time point, which in turn did not result from differences in various time intervals from first milk sampling to the colostrum phase. Furthermore, L-carnitine had no effect on NEB. The absence of an improved NEB in CAR during early lactation might have been caused by the increase of energy output in milk in this group and the simultaneously unchanged NEL intake. Carlson et al. [
6] found lower milk yields in the first 6 weeks of lactation for cows supplemented with 100 g L-carnitine/d, which might be due to the lower DMI as discussed above, whereas a supplementation with 6 or 50 g/d had no impact on milk yield. In the present experiment, milk fat and protein were also differently affected by L-carnitine supplementation over time. Carlson et al. [
6] detected higher milk fat concentrations in pluriparous Holstein cows, which received a top dress mixture with 50 or 100 g L-carnitine/d from day 14
ap until day 21
pp, and also, Pirestani et al. [
7] reported the same effect in L-carnitine-supplemented pluriparous Holstein cows (50 g/d/cow). Higher serum TG concentrations in CAR
ap, which were utilisable for milk fat synthesis, might be one possible explanation for higher milk fat percentages in the first week of lactation in CAR. According to Carlson et al. [
5], L-carnitine-supplemented cows might have a higher capacity to oxidise NEFA to BHB, which represent precursors for milk fat synthesis.
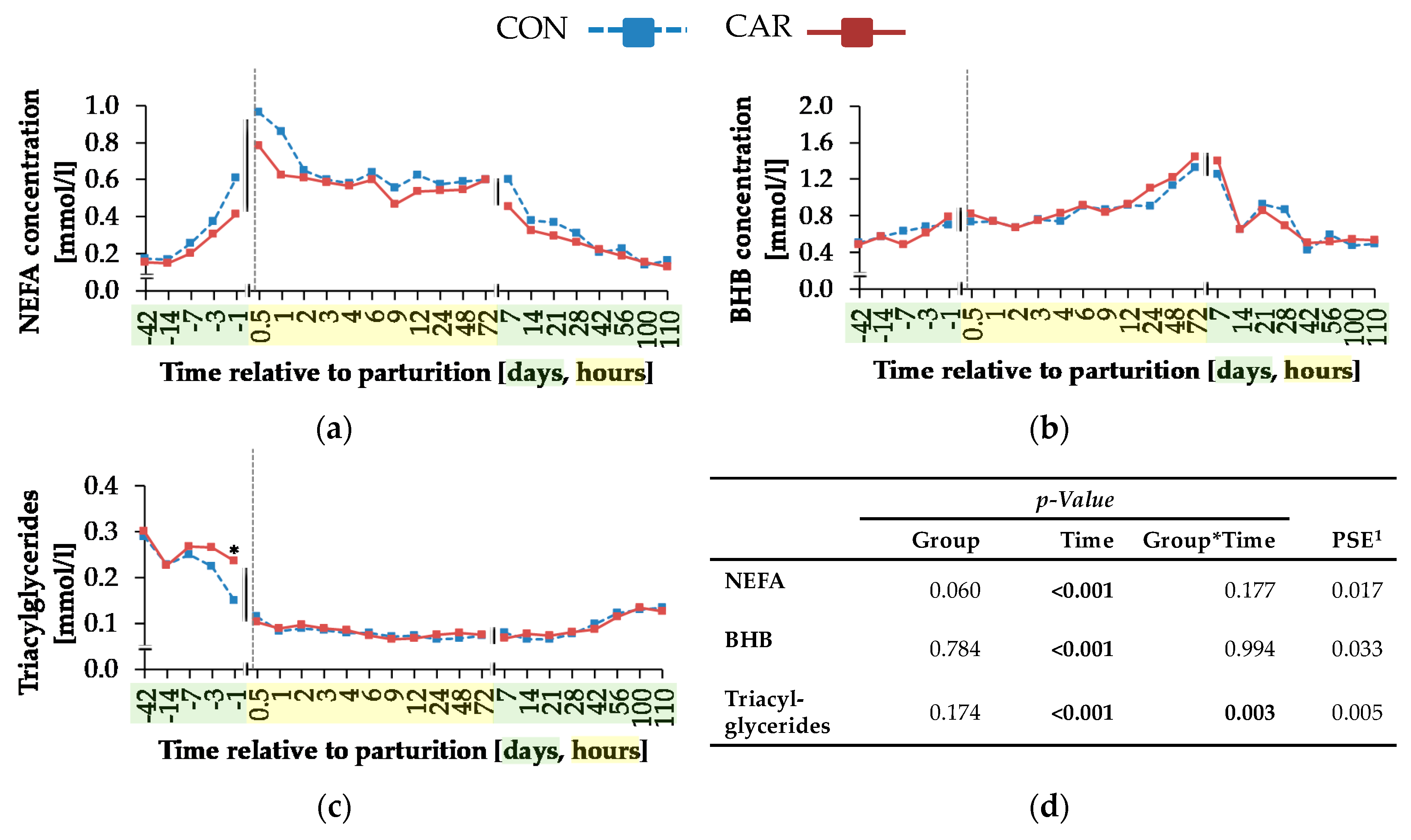
While NEFA and BHB concentrations were not influenced by L-carnitine, the concentrations of TG were significantly higher in L-carnitine-supplemented cows shortly before calving. The ultrasonographic evaluation of the dynamics of different adipose tissues confirmed the view that L-carnitine did not influence energy metabolism and thus lipomobilisation. Also, FPR was not affected by L-carnitine supplementation. According to Buttchereit et al. [
25], FPR is an adequate indicator for NEB during early lactation. Therefore, the unaltered FPR further supports the lack of effects by L-carnitine on lipomobilisation. Carlson et al. [
5] detected a decreased TG accumulation in liver and increased serum BHB concentrations in abomasal L-carnitine-infused (20 g L-carnitine/d) and feed-restricted cows by simultaneously unaltered NEFA concentrations in comparison to abomasal water-infused and feed-restricted cows. This L-carnitine supplementation was nearly equal to the dietary supplementation in the present experiment, in which 25 g L-carnitine per cow and day were available for intestinal absorption in consequence of using a rumen-protected L-carnitine product (80% of dietary carnitine would be degraded in the rumen [
6]). The higher levels of TG concentration in serum in CAR in the present study might be a hint at the mentioned decreased TG accumulation in liver. Also, serum NEFA concentrations were unaffected, but no increase of the BHB concentration in L-carnitine-supplemented cows was observed in the present study. This controversial result might be due to the difference between the physiological progression of a negative energy balance during the period of transition in the present experiment and the induction of this by an abrupt feed restriction outside this period, as in the trial of Carlson et al. [
5]. These results indicated that in dietary L-carnitine-supplemented periparturient cows, an increase of ketogenesis was not associated with putative simultaneously decreased TG accumulation in liver. Furthermore, Pirestani et al. [
7] detected a higher BHB concentration in cows receiving a combination of L-carnitine (50 g/d) and choline (60 g/d). Carlson et al. [
6] also found a decreased TG accumulation in liver of periparturient cows, which received 6, 50 or 100 g of a non-rumen-protected dietary L-carnitine product per day. The BHB concentration in blood was higher in the 50 and 100 g groups, while NEFA concentrations were unaffected by L-carnitine supplementation. Apart from the fact that the extent of ruminal degradation of L-carnitine is unclear, the cows in the experiment of Carlson et al. [
6] received anionic salts, minerals and vitamins from different sources, which might have had an effect, too. Furthermore, the L-carnitine supplementation in the experiment of Carlson et al. [
6] started at 14 days
ap, whereas the cows in the present study were already supplemented from day 42
ap. This longer adaptation period might have also been a reason for differences between the results of both studies.
In the present experiment, the maximum NEFA concentration was reached at 0.5 h
pp, when NEB was most negative and also fat mobilisation was at the maximum level. From day 100
pp, the initial NEFA concentration was reached again, which coincided with changing from a negative to a positive NEB. The increased BHB concentration went along with low glucose concentrations. The maximum BHB concentration occurred at 72 h
pp. This might have been the time point, when the demand for glucose was higher than the capacity for gluconeogenesis in liver, resulting in an incomplete oxidation of NEFA—named ketogenesis [
4]. Also, Carlson et al. [
6] described that glucose in blood was unaffected by 6, 50 or 100 g of a dietary L-carnitine product. The concentration of glucose increased rapidly in the first hour
pp, whereas insulin concentration reached the minimum level at this time point. Interestingly, the peak of glucose concentration in the present experiment occurred already at 2 h
pp and at 12 h
pp, the initial level was nearly reached again. According to Weber et al. [
26], the concentration of glucose decreased rapidly after calving and also, insulin concentration decreased around calving, which corresponds to the results in the present experiment. The time-dependent changes in glucose, insulin and NEFA concentrations suggested a lower
pp insulin sensitivity, as suggested by the lower RQUICKI that was, however, not influenced by L-carnitine supplementation. The significantly higher TG concentration in CAR on day 1
ap might be a hint at an increased hepatic export capacity for TG in the form of a very low density of lipoproteins in the blood.
Interestingly, in the first week of lactation, milk protein was higher in CON than in CAR. La Count et al. [
1] could not detect this effect in lactating Holstein cows receiving a ruminal or abomasal infusion of 6 g non-rumen-protected L-carnitine/d, whereas an abomasal infusion of a larger amount of L-carnitine (20 g/d) also resulted in lower milk protein concentrations in Holstein cows with provoked negative energy balance through feed restriction [
5]. Erfle et al. [
10] detected a negative correlation between milk carnitine and milk protein during the first weeks after calving in non-supplemented cows. A competition between milk protein synthesis and carnitine synthesis might exist, but is not very likely in the present case because precursors of carnitine synthesis derived from tissue protein catabolism [
10] and from milk protein were lower in CAR. Supplementation with L-carnitine should minimise the need for L-carnitine precursors from protein catabolism and endogenous L-carnitine synthesis should also be lower. In the study of Carlson et al. [
6], the percentage of milk protein tended to be higher in cows supplemented with 100 g L-carnitine/d than in cows supplemented with 50 g L-carnitine/d. In consideration of the present data, no conclusive explanation for this effect could be found.
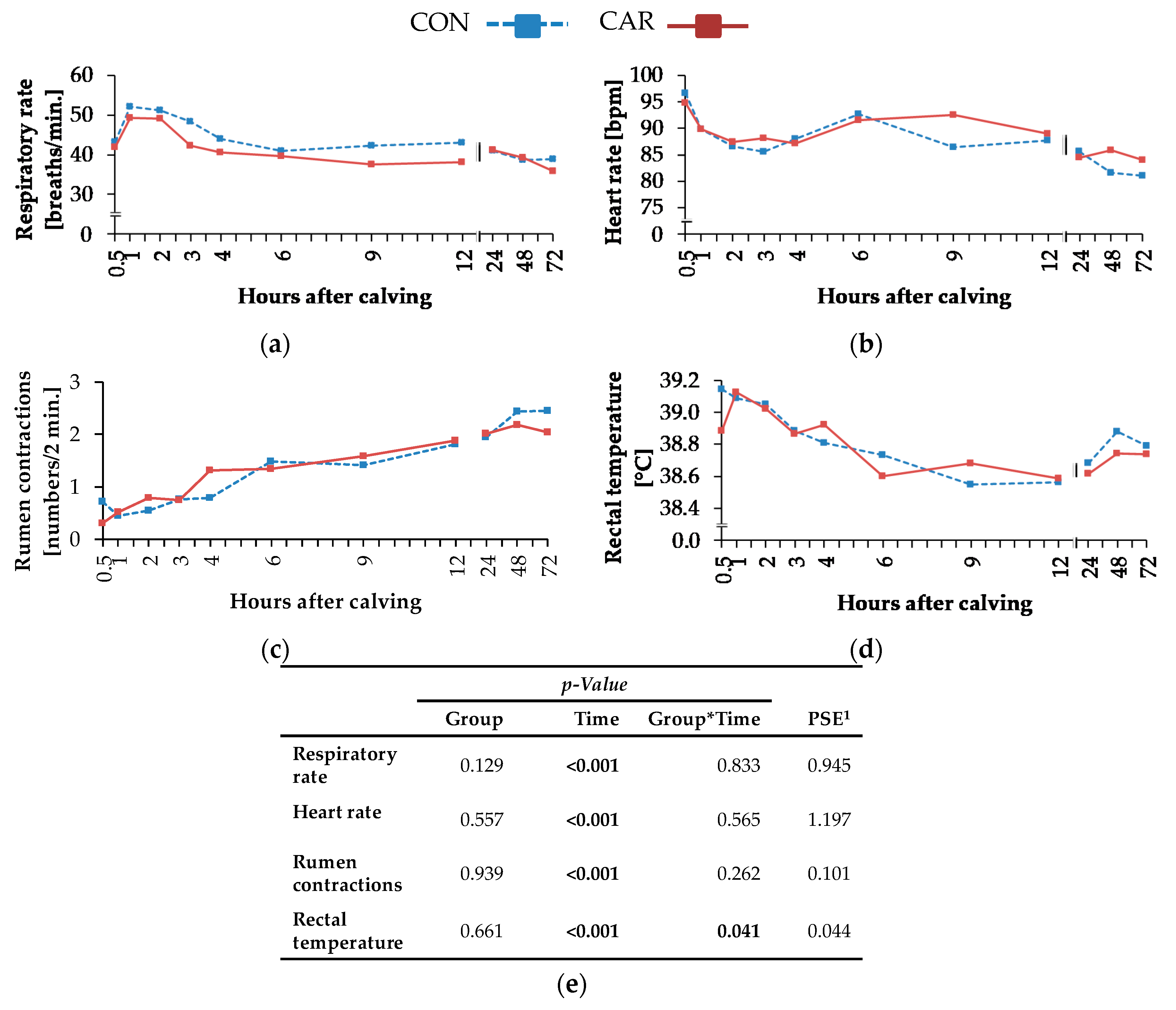
Most of the clinical signs and the cumulative clinical score were characterised by distinct dynamic profiles. Maximum heart rate of Holstein Friesian cows was detected directly during calving (Kovács et al. [
27,
28]). This effect resulted from exertions during parturition but also from loss of volume in the form of the calf and amniotic fluid. After this, peak heart rate remained on a high level from 0 h
pp until 0.5 h
pp and decreased until 1 h
pp, which corresponded to the results of the present experiment. Georg et al. [
29] found the heart frequency to increase 60–90 min
ap in Holstein cows. The process of calving is stressful and goes along with pain for the dam [
8]. Acute stress results in an activation of the sympathetic nervous system and concomitantly, a release of epinephrine from the adrenal glands causing an increase of heart rate [
30]. Furthermore, the mentioned development of heart rate might be due to the decreased parasympathetic activity, which resulted in an increase of sympathetic activity [
28]. Heart rate is closely related to metabolic oxygen consumption and consequently to energy expenditure [
31]. Thus, an increased heart rate goes along with increased oxygen consumption and indicates enhanced energy expenditure. Based on the fact that both treatment groups showed similar patterns of heart rate kinetics during the first 72 h
pp, the L-carnitine supplementation obviously failed to support energy metabolism in this particular period.
Respiratory rate is closely involved in oxygen supply of the organism and consequently to heart rate, but also in the regulation of acid-base homeostasis. In the present study, respiratory rate increased to maximum level 1 h
pp and was 26% higher at this time point than 72 h
pp, which was also caused by exertions due to parturition. Srikandakumar et al. [
32] detected an increase of respiratory rate and venous TpO
2 in heat-stressed Holstein cows, whereby TpCO
2 and oxygen saturation decreased. Taking into account that the calving process is a stress challenge, too, this corresponds to most of the results in the present experiment, where the maximum value of TpO
2 occurred simultaneously with highest respiratory rate (1 h
pp), whereby TpCO
2 was on a relatively low level at this time point in comparison to the following time points (maximum level of TpCO
2 in the first 72 h
pp occurred when respiratory rate reached the highest level). Maximum sO
2 also happened at 1 h
pp in this experiment, in which the cows calved outside the summer months and therefore, more oxygen was in the air. SO
2 also increased in Jerseys and Australian Milking Zebus, but not in Holstein cows under heat stress [
32]. The minimum concentration of hydrogen carbonate ions, total carbon dioxide, BE and Beecf occurred simultaneously to the peak of respiratory rate, whereby temperature-corrected pH reached the minimum level 1 h later. An increased respiratory rate results in an increase of carbon dioxide elimination [
32]. In the following, TpCO
2 and total carbon dioxide concentration decrease, whereby the limit of bicarbonate buffer system in blood is exceeded [
32]. Due to decreasing concentration of hydrogen carbonate ions, temperature-corrected pH in blood decreases, too [
32]. The discussed regulatory mechanisms between respiratory rate and acid-base homeostasis were not modulated by L-carnitine, further supporting the view that the energetic situation shortly after calving remained unaffected by L-carnitine.
Numbers of primary rumen contractions decreased due to a depressed DMI until week 1
ap. Also, Kandylis et al. [
33] ascribed decreased ruminal motility to a decreased feed intake in sulfur-supplemented ruminants. Jørgensen et al. [
34] detected decreased rumen motility in hypocalcaemic cows at a Ca
2+ concentration of ≤1.0 mmol/L in blood. In the present experiment, Ca
2+ concentration in blood was at 1.05 mmol/L when rumen contractions reached the minimal level (1 h
pp). According to Hove et al. [
35], decreased gastrointestinal motility went along with decreased calcium absorption because calcium absorption in the small intestine depends on a continuous flow of substrate.
Although the rectal temperature was differently affected by carnitine supplementation over time, the overall variation was minimum (38.58–39.11 °C) and inconsistently related to the dietary treatment. According to Suthar et al. [
36] and Burfeind et al. [
37], rectal temperature is influenced by parity, time of day and month, which might have contributed to the observed variation.
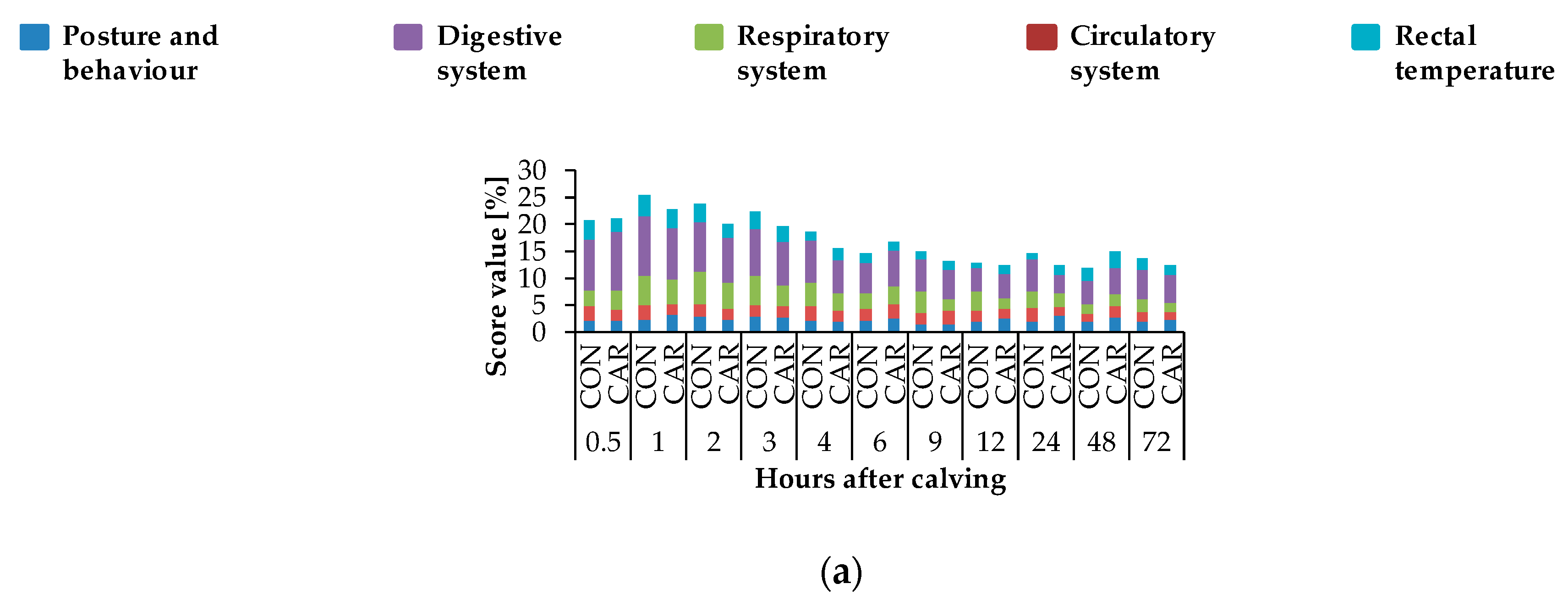
Just like most of the discussed clinical variables (heart rate, respiratory rate, primary rumen contractions and rectal temperature) showed maximum values shortly after calving, the resulting cumulative clinical score also peaked at 1 h pp, whereby deviations in the digestive system at each time point provided the largest part of the cumulative clinical score and reached the highest percentage during the first h pp.
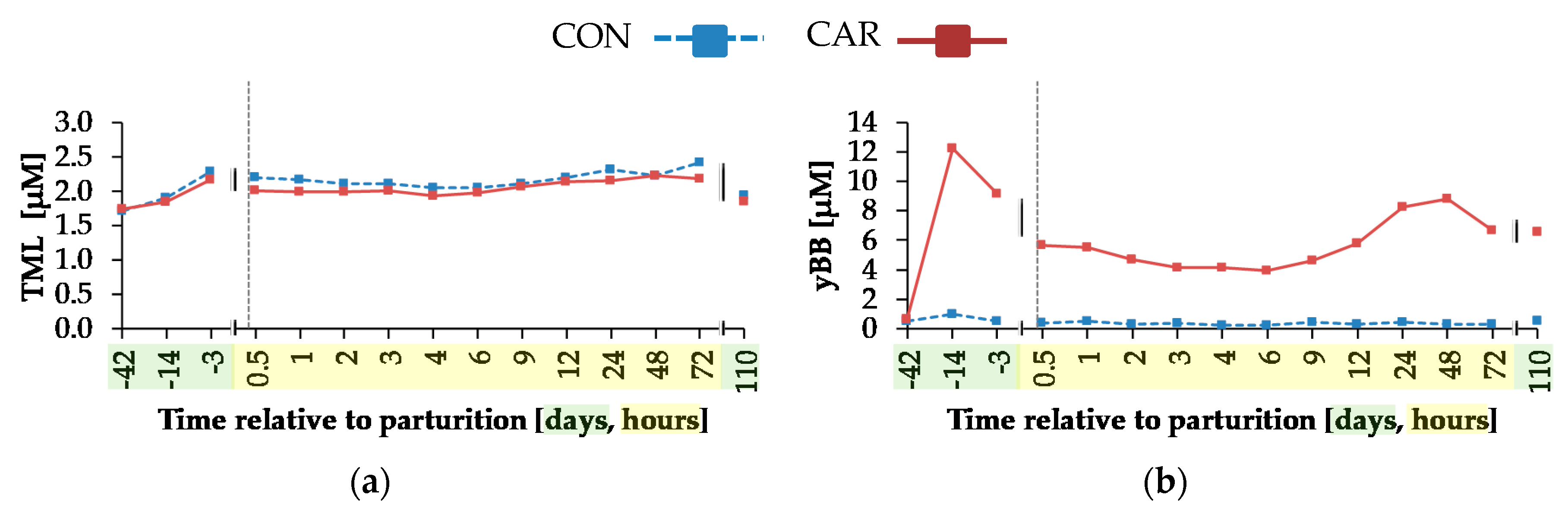
A time-dependent variation was detected for the concentration of TML as the precursor for endogenous carnitine synthesis. In mammals, TML originates primarily from protein degradation [
38] and was found in different proteins like cytochrome c, calmodulin, actin, myosin and histones [
39]. In the present experiment, increased TML concentrations were detected until day 3
ap independent of L-carnitine supplementation. The lack of treatment differences might be due to the equal total NEL intake, the NEB and therefore, an assumed equal protein catabolism between both groups. Davis et al. [
40] described a significant increase of TML concentration in plasma of starved rats as compared to fed rats, which possibly resulted from an induced proteolysis. Dairy cows in the transition period also have an insufficient energy supply, triggering not only fat mobilisation but also protein degradation as already indicated by the TML concentration in blood.
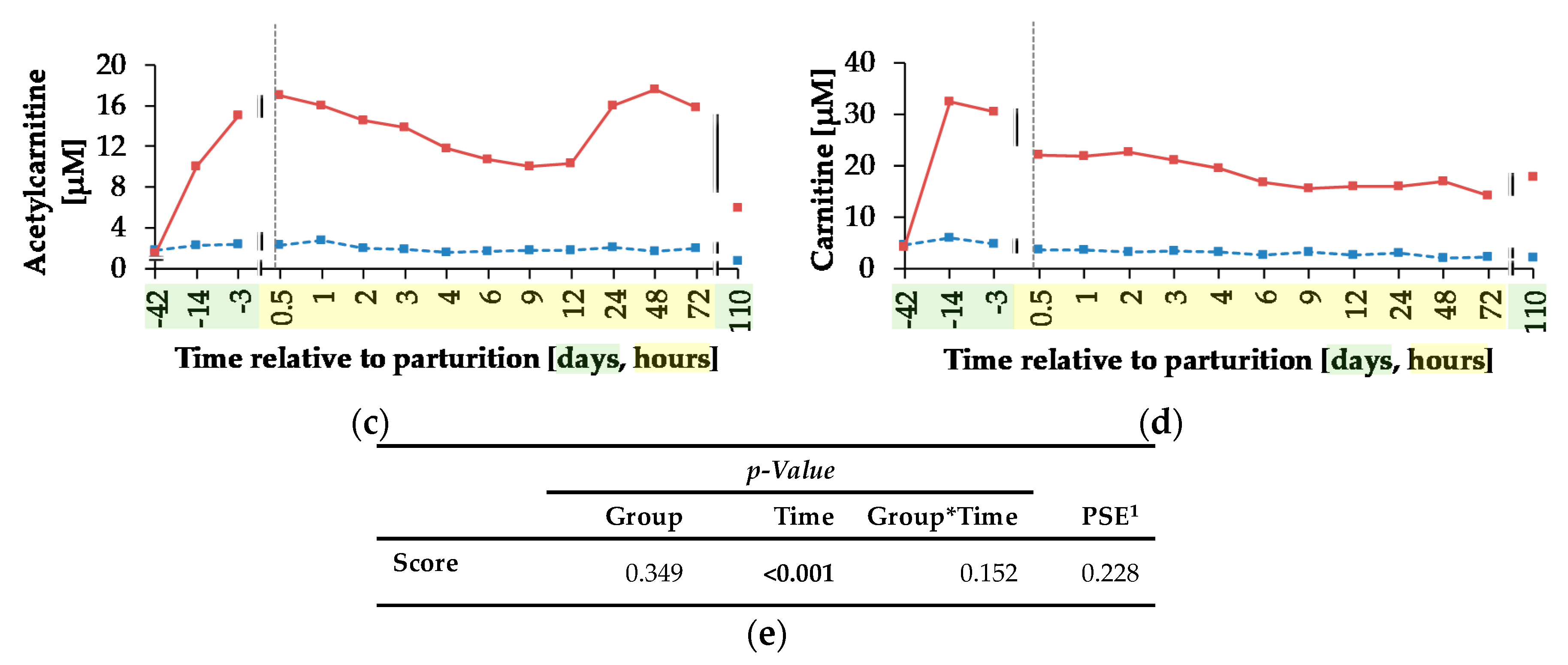
The concentrations of yBB, ACA and CA were affected by L-carnitine supplementation. All of them remained unchanged in CON over the whole trial, but in CAR, there was a sharp increase of yBB and CA concentration until day 14
ap, which represented the period of adaptation to the L-carnitine supplementation. yBB originated from the conversion of TML occurring primarily in kidney in rats [
41]. Rebouche et al. [
42] detected a decreased hepatic activity of yBB dioxygenase—converting yBB into CA—in carnitine-supplemented rats, which could be the reason for the higher yBB concentrations in CAR. The decreased yBB and CA concentrations from day 14
ap until 0.5 h
pp in both groups could result from variations in carnitine metabolism during this period. Also, according to Schlegel et al. [
43], mRNA abundances of genes involved in fatty acid uptake, fatty acid oxidation, ketogenesis and of enzymes of carnitine synthesis as well as carnitine uptake in liver increased from week 3
ap until week 1
pp in dairy Holstein cows and thus, liver cells received sufficient amounts of carnitine for the transport of excessive amounts of NEFA. CA is released primarily as ACA from liver cells into the blood stream [
44], which explains the persistent increase of ACA concentration at 0.5 h
pp, when concentrations of yBB and CA decreased in blood, but increased in liver cells [
43].

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











