Live Feeds Used in the Larval Culture of Red Cusk Eel, Genypterus chilensis, Carry High Levels of Antimicrobial-Resistant Bacteria and Antibiotic-Resistance Genes (ARGs)




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Live-Feed Culture Sampling
2.3. Sample Processing and Culturable Bacterial Count
2.4. Bacterial Isolates
2.5. Bacterial Identification
2.6. Minimum Inhibitory Concentrations (MICs) of Florfenicol
2.7. Antibacterial Resistance Patterns
2.8. Detection of Genes Encoding for Florfenicol Resistance
2.9. Isolation of Plasmid DNA
2.10. Statistical Analysis
3. Results
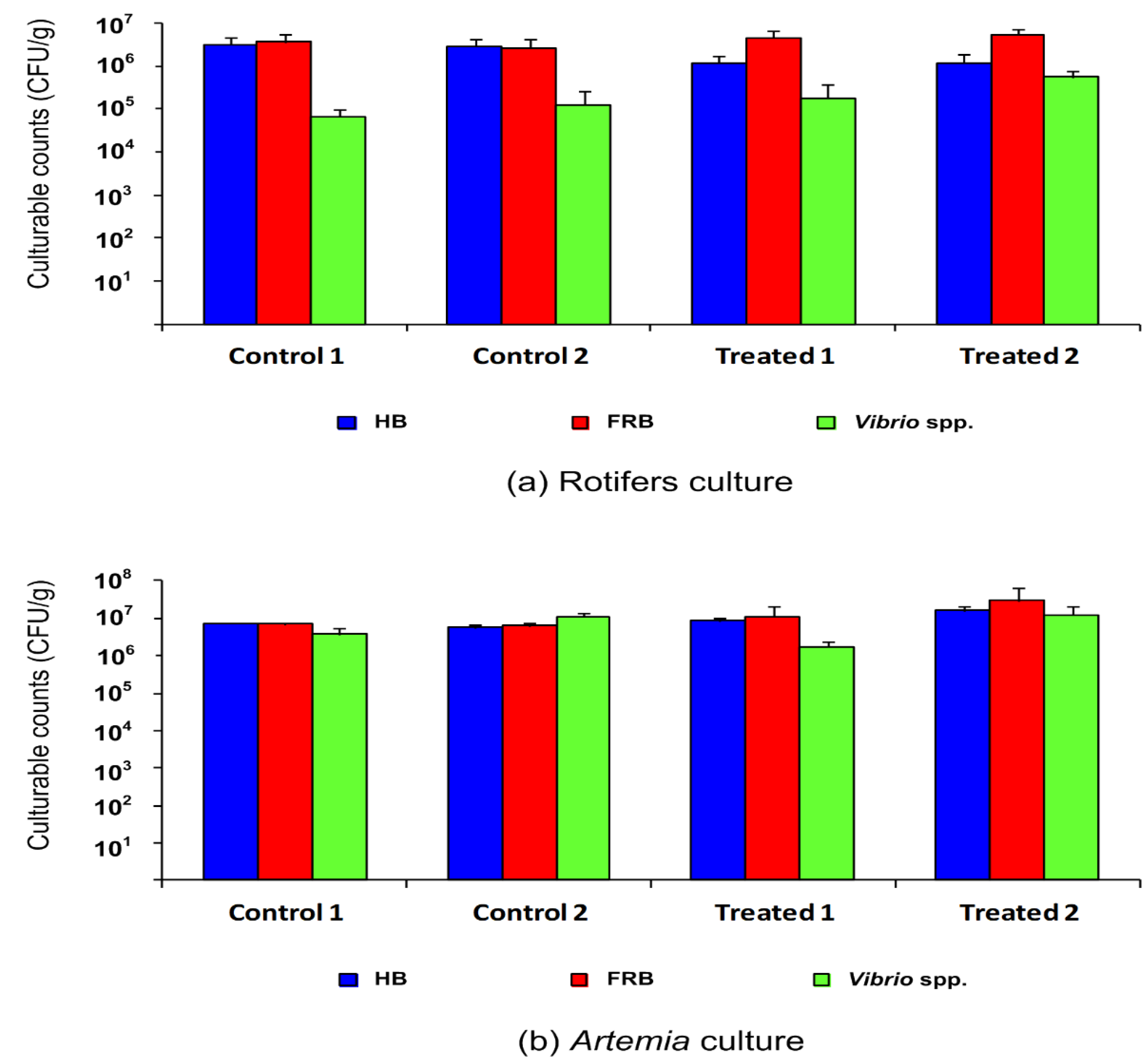
3.1. Culturable Bacterial Counts
3.2. Bacterial Isolates
3.3. Minimum Inhibitory Concentrations (MICs)
3.4. Antimicrobial Resistance Patterns
3.5. Phenicol Resistance Genes
3.6. Plasmid Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jara-Seguel, P.; Ubilla, A.; Estrada, J.M.; Ramírez, D.; Valdebenito, I. Nuclear DNA content in the red conger eel Genypterus chilensis (Guichenot, 1881) (Actinopterygii: Ophidiidae). Gayana 2011, 75, 198–200. [Google Scholar] [CrossRef] [Green Version]
- Córdova-Alarcón, V.R.; Araneda, C.; Jilberto, F.; Magnolfi, P.; Toledo, M.I.; Lam, N. Genetic Diversity and Population Structure of Genypterus chilensis, a Commercial Benthic Marine Species of the South Pacific. Front. Mar. Sci. 2019, 6, 748. [Google Scholar] [CrossRef] [Green Version]
- Levican, A.; Avendaño-Herrera, R. Bacteria associated with mass mortality of post-larvae of red conger eel (Genypterus chilensis) cultured in a Chilean farm. Bull. Eur. Assoc. Fish Pathol. 2015, 35, 162–169. [Google Scholar]
- Lasa, A.; Avendaño-Herrera, R.; Estrada, J.M.; Romalde, J.L. Isolation and identification of Vibrio toranzoniae associated with diseased red conger eel (Genypterus chilensis) farmed in Chile. Vet. Microbiol. 2015, 179, 327–331. [Google Scholar] [CrossRef]
- Levican, A.; Lasa, A.; Irgang, R.; Romalde, J.L.; Poblete-Morales, M.; Avendaño-Herrera, R. Isolation of Vibrio tapetis from two native fish species (Genypterus chilensis and Paralichthys adspersus) reared in Chile and description of Vibrio tapetis subsp. quintayensis subsp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 716–723. [Google Scholar] [CrossRef]
- Irgang, R.; González-Luna, R.; Gutíerrez, J.; Poblete-Morales, M.; Rojas, V.; Tapia-Cammas, D.; Avendaño-Herrera, R. First identification and characterization of Tenacibaculum dicentrarchi isolated from Chilean red conger eel (Genypterus chilensis, Guichenot 1881). J. Fish Dis. 2017, 40, 1915–1920. [Google Scholar] [CrossRef]
- Verdonck, L.; Swings, J.; Kersters, K.; Dehasque, M.; Sorgeloos, P.; Leger, P. Variability of the microbial environment of rotifer Brachionus plicatilis and Artemia production systems. J. World Aquac. Soc. 1994, 25, 55–59. [Google Scholar] [CrossRef]
- Verdonck, L.; Grisez, L.; Sweetman, E.; Minkoff, I.G.; Sorgeloos, P.; Ollevier, F.; Swings, J. Vibrios associated with routine productions of Brachionus plicatilis. Aquaculture 1997, 149, 203–214. [Google Scholar] [CrossRef]
- Rombaut, G.; Suantika, G.; Boon, N.; Maertens, S.; Dhert, P.; Top, E.; Sorgeloos, P.; Verstraete, W. Monitoring of the evolving diversity of the microbial community present in rotifer cultures. Aquaculture 2001, 198, 237–252. [Google Scholar] [CrossRef]
- Pérez-Benavente, G.P.; Gatesoupe, F.J. Bacteria associated with cultured rotifers and Artemia are detrimental to larval turbot, Scophthalmus maximus L. Aquac. Eng. 1988, 7, 289–293. [Google Scholar] [CrossRef]
- Nicolas, J.L.; Robic, E.; Ansquer, D. Bacterial flora associated with a trophic chain consisting of microalgae, rotifers, and turbot larvae: Influence of bacteria on larval survival. Aquaculture 1989, 83, 237–248. [Google Scholar] [CrossRef]
- Orozco-Medina, C.; Maeda-Martínez, A.; López-Cortés, A. Effect of aerobic Gram-positive heterotrophic bacteria associated with Artemia franciscana cysts on the survival and development of its larvae. Aquaculture 2002, 213, 15–29. [Google Scholar] [CrossRef]
- Kumaran, T.; Citarasu, T. Isolation and Characterization of Vibrio Species from Shrimp and Artemia Culture and Evaluation of the Potential Virulence Factor. Intel. Prop. Rights 2016, 4, 153. [Google Scholar] [CrossRef] [Green Version]
- Vallés, R.; Roque, A.; Caballero, A.; Estévez, A. Use of Ox-Aquaculture© for disinfection of live prey and meagre larvae, Argyrosomus regius (Asso, 1801). Aquac. Res. 2015, 46, 413–419. [Google Scholar] [CrossRef]
- Skjermo, J.; Vadstein, O. Characterization of the bacterial flora of mass cultivated Brachionus plicatilis. Hydrobiologia 1993, 255, 185–191. [Google Scholar] [CrossRef]
- Verschuere, L.; Dhont, J.; Sorgeloos, P.; Verstraete, W. Monitoring Biolog patterns and r/K-strategists in the intensive culture of Artemia juveniles. J. Appl. Microbiol. 1997, 83, 603–612. [Google Scholar] [CrossRef]
- Grisez, L.; Chair, M.; Sorgeloos, P.; Ollevier, F. Mode of infection and spread of Vibrio anguillarum in turbot (Scophthalmus maximus) larvae after oral challenge through live feed. Dis. Aquat. Org. 1996, 26, 181–187. [Google Scholar] [CrossRef]
- Dhert, P.; Rombaut, G.; Suantika, G.; Sorgeloos, P. Advancement of rotifer culture and manipulation techniques in Europe. Aquaculture 2001, 200, 129–146. [Google Scholar] [CrossRef] [Green Version]
- Giménez, G.; Padrós, F.; Roque, A.; Estévez, A.; Furones, D. Bacterial load reduction of live prey for fish larval feeding using Ox-Aquaculture©. Aquac. Res. 2006, 37, 1130–1139. [Google Scholar] [CrossRef]
- Jayasree, L.; Sonali, S.M.; Purbali, S.; Philipose, K.K. Bacterial Flora of Water and Rotifers in Outdoor Mass Culture Tanks Fed with Different Microalagal Diets. J. Life Sci. 2016, 10, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Prol-Garcia, M.J.; Planas, M.; Pintado, J. Different Colonization and Residence Time of Listonella anguillarum and Vibrio splendidus in the Rotifer Brachionus plicatilis Determined by Real-time PCR and DGGE. Aquaculture 2010, 302, 26–35. [Google Scholar] [CrossRef]
- Austin, B.; Allen, B.W. Microbiology of laboratory-hatched brine shrimp (Artemia). Aquaculture 1981, 26, 369–383. [Google Scholar] [CrossRef]
- Reid, H.I.; Treasurer, J.W.; Adam, B.; Birkbeck, T.H. Analysis of bacterial populations in the gut of developing cod larvae and identification of Vibrio logei, Vibrio anguillarum and Vibrio splendidus as pathogens of cod larvae. Aquaculture 2009, 288, 36–43. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The Gut Microbiota of Marine Fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef] [PubMed]
- Vadstein, O.; Attramadal, K.J.K.; Bakke, I.; Forberg, T.; Olsen, Y.; Verdegem, M.; Giatsis, C.; Skjermo, J.; Aasen, I.M.; Gatesoupe, F.-J.; et al. Managing the Microbial Community of Marine Fish Larvae: A Holistic Perspective for Larviculture. Front. Microbiol. 2018, 9, 1820. [Google Scholar] [CrossRef] [PubMed]
- Grisez, L.; Reyniers, J.; Verdonck, L.; Swings, J.; Ollevier, F. Dominant intestinal microflora of sea bream and sea bass larvae, from two hatcheries, during larval development. Aquaculture 1997, 155, 387–399. [Google Scholar] [CrossRef]
- Gómez-Gil, B.; Abreu-Grobois, F.A.; Romero-Jarero, J.; Herrera-Vega, M. Chemical disinfection of Artemia nauplii. J. World Aquac. Soc. 1994, 25, 579–583. [Google Scholar] [CrossRef]
- Martínez-Díaz, S.F.; Álvarez-González, C.A.; Moreno Legorreta, M.; Vázquez-Juárez, R.; Barrios-González, J. Elimination of the associated microbial community and bioencapsulation of bacteria in the rotifer Brachionus plicatilis. Aquac. Int. 2003, 11, 95–108. [Google Scholar] [CrossRef]
- Munro, P.D.; Henderson, R.J.; Barbour, A.; Birkbeck, T.H. Partial decontamination of rotifers with ultraviolet radiation: The effect of changes in the bacterial load and flora of rotifers on mortalities in start-feeding larval turbot. Aquaculture 1999, 170, 229–244. [Google Scholar] [CrossRef]
- Douillet, P.A. Bacterial additives that consistently enhance rotifer growth under synxenic culture conditions. 2. Use of single and multiple bacterial probiotics. Aquaculture 2000, 182, 241–248. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatesoupe, F.-J. Probiotic and formaldehyde treatments of Artemia nauplii as food for larval pollack, Pollachius pollachius. Aquaculture 2002, 212, 347–360. [Google Scholar] [CrossRef] [Green Version]
- Villamil, L.; Figueras, A.; Planas, M.; Novoa, B. Control of Vibrio alginolyticus in Artemia culture by treatment with bacterial probiotics. Aquaculture 2003, 219, 43–56. [Google Scholar] [CrossRef]
- Tolomei, A.; Burke, C.; Crear, B.; Carson, J. Bacterial decontamination of on-grown Artemia. Aquaculture 2004, 232, 357–371. [Google Scholar] [CrossRef]
- Marques, A.; Thanh, T.H.; Sorgeloos, P.; Bossier, P. Use of microalgae and bacteria to enhance protection of gnotobiotic Artemia against different pathogens. Aquaculture 2006, 258, 116–126. [Google Scholar] [CrossRef]
- Haché, R.; Lanteigne, C.; Hébert, Y. Salt as a decontamination agent to control bacterial load in Artemia salina cultures. Aquaculture 2006, 452, 24–27. [Google Scholar] [CrossRef]
- Gatesoupe, F.J.; Lambert, C.; Nicolas, J.L. Pathogenicity of Vibrio splendidus strains associated with turbot larvae, Scophthalmus maximus. J. Appl. Microbiol. 1999, 87, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Planas, M.; Pérez-Lorenzo, M.; Hjelm, M.; Gram, L.; Uglenes Fiksdal, I.; Bergh, Ø.; Pintado, J. Probiotic effect in vivo of Roseobacter strain 27-4 against Vibrio (Listonella) anguillarum infections in turbot (Scophthalmus maximus L.) larvae. Aquaculture 2006, 255, 323–333. [Google Scholar] [CrossRef] [Green Version]
- D’Alvise, P.W.; Lillebø, S.; Prol-Garcia, M.J.; Wergeland, H.; Nielsen, K.F.; Bergh, Ø.; Gram, L. Phaeobacter gallaeciensis reduces Vibrio anguillarum in cultures of microalgae and rotifers, and prevents vibriosis in cod larvae. PLoS ONE 2012, 7, e43996. [Google Scholar] [CrossRef] [Green Version]
- Skjermo, J.; Bakke, I.; Dahle, S.W.; Vadstein, O. Probiotic strains introduced through live feed and rearing water have low colonizing success in developing Atlantic cod larvae. Aquaculture 2015, 438, 17–23. [Google Scholar] [CrossRef]
- Quiroz-Guzmán, E.; Vázquez-Juárez, R.; Luna-González, A.; Balcázar, J.L.; Barajas-Sandoval, D.R.; Martínez-Díaz, S.F. Administration of Probiotics Improves the Brine Shrimp Production and Prevents Detrimental Effects of Pathogenic Vibrio Species. Mar. Biotechnol. 2018, 20, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Sahandi, J.; Sorgeloos, P.; Xiao, H.; Wang, X.; Qi, Z.; Zheng, Y.; Tang, X. The Use of Selected Bacteria and Yeasts to Control Vibrio spp. in Live Food. Antibiotics 2019, 8, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haamed, A.S.; Balasubramanian, G. Antibiotic resistance in bacteria isolated from Artemia nauplii and efficacy of formaldehyde to control bacterial load. Aquaculture 2000, 183, 195–205. [Google Scholar] [CrossRef]
- Miranda, C.D.; Godoy, F.A.; Lee, M.R. Current Status of the Use of Antibiotics and the Antimicrobial Resistance in the Chilean Salmon Farms. Front. Microbiol. 2018, 9, 1284. [Google Scholar] [CrossRef]
- Fernández-Alarcón, C.; Miranda, C.D.; Singer, R.S.; López, Y.; Rojas, R.; Bello, H.; Domínguez, M.; González-Rocha, G. Detection of the floR gene in a diversity of florfenicol resistant Gram-negative bacilli from freshwater salmon farms in Chile. Zoon. Public Health 2010, 57, 181–188. [Google Scholar] [CrossRef]
- Roberts, M.C.; Schwarz, S. Tetracycline and Phenicol Resistance Genes and Mechanisms: Importance for Agriculture, the Environment, and Humans. J. Environ. Qual. 2016, 45, 576–592. [Google Scholar] [CrossRef]
- Hong, B.; Ba, Y.; Niu, L.; Lou, F.; Zhang, Z.; Liu, H.; Pan, Y.; Zhao, Y. A Comprehensive Research on Antibiotic Resistance Genes in Microbiota of Aquatic Animals. Front. Microbiol. 2018, 9, 1617. [Google Scholar] [CrossRef]
- Schwarz, S.; White, D.G. Phenicol Resistance. In Frontiers in Antimicrobial Resistance; White, D.G., Alekshun, M.N., McDermott, P.F., Eds.; American Society for Microbiology, ASM Press: Washington, DC, USA, 2005; pp. 124–147. [Google Scholar]
- Schmidt, A.S.; Bruun, M.S.; Dalsgaard, I.; Larsen, J.L. Incidence, distribution, and spread of tetracycline resistance determinants and integron-associated antibiotic resistance genes among motile aeromonads from a fish farming environment. Appl. Environ. Microbiol. 2001, 67, 5675–5682. [Google Scholar] [CrossRef] [Green Version]
- Miranda, C.D.; Rojas, R.; Abarca, A.; Hurtado, L. Effect of florfenicol and oxytetracycline treatments on the intensive larval culture of the Chilean scallop Argopecten purpuratus (Lamarck, 1819). Aquac. Res. 2014, 45, 16–30. [Google Scholar] [CrossRef]
- Miranda, C.D.; Rojas, R. Occurrence of florfenicol resistance in bacteria associated with two Chilean salmon farms with different history of antibacterial usage. Aquaculture 2007, 266, 39–46. [Google Scholar] [CrossRef]
- Buller, N.B. Bacteriological Culture Techniques: Microscopy, Culture and Identification. In Bacteria from Fish and Other Aquatic Animals: A Practical Identification Manual, 1st ed.; Buller, N.B., Ed.; CABI Publishing: Cambridge, MA, USA, 2004; pp. 83–116. [Google Scholar]
- Opazo, R.; Ortúzar, F.; Navarrete, P.; Espejo, R.; Romero, J. Reduction of soybean meal non-starch polysaccharides and α-Galactosides by solid-state fermentation using cellulolytic bacteria obtained from different environments. PLoS ONE 2012, 7, e44783. [Google Scholar] [CrossRef] [PubMed]
- Ribosomal Database Project. Available online: http://rdp.cme.msu.edu/ (accessed on 5 January 2020).
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Approved Standard, 10th ed.; M07-A10; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2015. [Google Scholar]
- Steers, E.; Foltz, E.L.; Graves, B.S.; Riden, J. An inocula replicating apparatus for routine testing of bacterial susceptibility to antibiotics. Antibiot. Chemother. 1959, 9, 307–311. [Google Scholar]
- CLSI. Methods for Antimicrobial Disk Susceptibility Testing of Bacteria Isolated from Aquatic Animals; Approved Guideline VET03-A; Number 23; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2006; Volume 26. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI Supplement VET08; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2018. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Test; Approved Standard, 12th ed.; M02-A12; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2015. [Google Scholar]
- Hinton, M.; Hedges, A.J.; Linton, A.H. The ecology of Escherichia coli in market claves fed a milk-substitute diet. J. Appl. Bacteriol. 1985, 58, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Higuera-Llantén, S.; Vásquez-Ponce, F.; Barrientos-Espinoza, B.; Mardones, F.O.; Marshall, S.H.; Olivares-Pacheco, J. Extended antibiotic treatment in salmon farms select multiresistant gut bacteria with a high prevalence of antibiotic resistance genes. PLoS ONE 2018, 13, e0203641. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, M.; Miranda, C.D.; Fuentes, O.; de la Fuente, M.; Godoy, F.A.; Bello-Toledo, H.; González-Rocha, G. Occurrence of transferable integrons and sul and dfr genes among sulfonamide-and/or trimethoprim-resistant bacteria isolated from Chilean salmonid farms. Front. Microbiol. 2019, 10, 748. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 1999; p. 931. [Google Scholar]
- Norusis, M.J. Statistical Package for the Social Sciences (SPSS) 12.0 Statistical Procedures Companion; Prentice Hall: Englewood Cliffs, NJ, USA, 2004; p. 601. [Google Scholar]
- NCCLS. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Approved Standard—2nd ed. ; NCCLS document M31-A2; NCCLS: Wayne, PA, USA, 2002; ISBN 1-56238-461-9. [Google Scholar]
- Miranda, C.D.; Rojas, R.; Garrido, M.; Geisse, J.; González, G. Role of shellfish hatchery as a reservoir of antimicrobial resistant bacteria. Mar. Pollut. Bull. 2013, 74, 334–343. [Google Scholar] [CrossRef]
- Abu-Rezq, T.; Al-Abdul-Elah, K.; Duremdez, R.; Al-Marzouk, A.; James, C.M.; Al-Gharabally, H.; Al-Shimmari, J. Studies on the effect of using the rotifer, Brachionus plicatilis, treated with different nutritional enrichment media and antibiotics on the growth and survival of blue-fin sea bream, Sparidentex hasta (Valenciennes), larvae. Aquac. Res. 2002, 33, 117–128. [Google Scholar] [CrossRef]
- Interaminense, J.A.; Calazans, N.F.; Cáritas do Valle, B.; Vogeley, J.L.; Peixoto, S.; Soares, R.; Lima Filho, J.V. Vibrio spp. Control at Brine Shrimp, Artemia, Hatching and Enrichment. J. World Aquac. 2014, 45, 65–74. [Google Scholar] [CrossRef]
- Williams, R.R.; Bell, T.A.; Lightner, D.V. Shrimp Antimicrobial Testing. II. Toxicity Testing and Safety Determination for Twelve Antimicrobials with Penaeid Shrimp Larvae. J. Aquat. Anim. Health 1992, 4, 262–270. [Google Scholar] [CrossRef]
- Soto-Rodríguez, S.; Armenta, M.; Gómez-Gil, B. Effects of enrofloxacin and florfenicol on survival and bacterial population in an experimental infection with luminescent Vibrio campbellii in shrimp larvae of Litopenaeus vannamei. Aquaculture 2006, 255, 48–54. [Google Scholar] [CrossRef]
- Botelho, R.G.; Christofoletti, C.A.; Correia, J.E.; Ansoar, Y.; Olinda, R.A.; Tornisielo, V.L. Genotoxic responses of juvenile tilapia (Oreochromis niloticus) exposed to florfenicol and oxytetracycline. Chemosphere 2015, 132, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Zhang, Y.; Zhang, X.; Zhang, O.; Cheng, R.; Mostafizur, R.M.; Liu, Y. Effects of florfenicol exposure on growth, development and antioxidant capacity of flounder Paralichthys olivaceus larvae at different developmental stages. J. Ocean. Limnol. 2019. [Google Scholar] [CrossRef]
- Patra, S.K.; Mohamed, K.S. Enrichment of Artemia nauplii with the probiotic yeast Sacharomyces boulardii and its resistance against a pathogenic Vibrio. Aquac. Int. 2003, 11, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Kalatzis, P.G.; Bastías, R.; Kokkari, C.; Katharios, P. Isolation and Characterization of Two Lytic Bacteriophages, φSt2 and φGrn1; Phage Therapy Application for Biological Control of Vibrio alginolyticus in Aquaculture Live Feeds. PLoS ONE. 2016, 11, e0151101. [Google Scholar] [CrossRef] [Green Version]
- Sotomayor, M.A.; Reyes, J.K.; Restrepo, L.; Domínguez-Borbor, C.; Maldonado, M.; Bayot, B. Efficacy assessment of commercially available natural products and antibiotics, commonly used for mitigation of pathogenic Vibrio outbreaks in Ecuadorian Penaeus (Litopenaeus) vannamei hatcheries. PLoS ONE 2019, 14, e0210478. [Google Scholar] [CrossRef]
- Gómez-Gil, B.; Thompson, F.; Thompson, C.C.; Swings, J. Vibrio rotiferianus sp nov., isolated from cultures of the rotifer Brachionus plicatilis. Int. J. Syst. Evol. Microbiol. 2003, 53, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Cataluña, A.; Lucena, T.; Tarazona, E.; Arahal, D.R.; Macián, M.C.; Pujalte, M.J. An MLSA approach for the taxonomic update of the Splendidus clade, a lineage containing several fish and shellfish pathogenic Vibrio spp. Syst. Appl. Microbiol. 2016, 39, 361–369. [Google Scholar] [CrossRef]
- Sawabe, T.; Ogura, Y.; Matsumura, Y.; Feng, G.; Gao, F.; Amin, A.K.M.; Mino, S.; Nakagawa, S.; Sawabe, T.; Kumar, R.; et al. Updating the Vibrio clades defined by multilocus sequence phylogeny: Proposal of eight new clades, and the description of Vibrio tritonius sp. nov. Front. Microbiol. 2013, 4, 414. [Google Scholar] [CrossRef] [Green Version]
- Al-Saari, N.; Gao, F.; Rohul, A.A.; Sato, K.; Sato, K.; Mino, S.; Suda, W.; Oshima, K.; Hattori, M.; Ohkuma, M.; et al. Advanced Microbial Taxonomy Combined with Genome-Based-Approaches Reveals that Vibrio astriarenae sp. nov., an Agarolytic Marine Bacterium, Forms a New Clade in Vibrionaceae. PLoS ONE 2015, 27, 10. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef] [Green Version]
- White, D.G.; Hudson, C.; Maurer, J.J.; Ayers, S.; Zhao, S.; Lee, M.D.; Bolton, L.; Foley, T.; Sherwood, J. Characterization of chloramphenicol and florfenicol resistance in Escherichia coli associated with bovine diarrhea. J. Clin. Microbiol. 2000, 38, 4593–4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, R.S.; Patterson, S.K.; Meier, A.E.; Gibson, J.K.; Lee, H.L.; Maddox, C.W. Relationship between phenotypic and genotypic florfenicol resistance in Escherichia coli. Antimicrob. Agents Chemother. 2004, 48, 4047–4049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, L.; Cloeckaert, A.; Doublet, B.; Schwarz, S.; Bouju-Albert, A.; Ganiere, J.P.; Le, B.H.; Le Fleche-Mateos, A.; Giraud, E. Complete sequence of the floR-carrying multiresistance plasmid pAB5S9 from freshwater Aeromonas bestiarum. J. Antimicrob. Chemother. 2008, 62, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, H.; Zhang, X.; Song, L.; Chang, Y.; Yang, G. Molecular determination of oxytetracycline-resistant bacteria and their resistance genes from mariculture environments of China. J. Appl. Microbiol. 2007, 103, 2580–2592. [Google Scholar] [CrossRef]
- Kim, E.; Aoki, T. Sequence analysis of the florfenicol resistance gene encoded in the transferable R-plasmid from a fish pathogen, Pasteurella piscicida. Microbiol. Immunol. 1996, 40, 665–669. [Google Scholar] [CrossRef]
- Cloeckaert, A.; Baucheron, S.; Flaujac, G.; Schwarz, S.; Kehrenberg, C.; Martel, J.L.; Chaslus-Dancla, E. Plasmid-mediated florfenicol resistance encoded by the floR gene in Escherichia coli isolated from cattle. Antimicrob. Agents Chemother. 2000, 44, 2858–2860. [Google Scholar] [CrossRef] [Green Version]
- Cloeckaert, A.; Baucheron, S.; Chaslus-Dancla, E. Nonenzymatic chloramphenicol resistance mediated by IncC plasmid R55 is encoded by a floR gene variant. Antimicrob. Agents Chemother. 2001, 45, 2381–2382. [Google Scholar] [CrossRef] [Green Version]
- da Silva, G.C.; Rossi, C.C.; Santana, M.F.; Langford, P.R.; Bosse, J.T.; Bazzolli, D.M.S. p518, a small floR plasmid from a south American isolate of Actinobacillus pleuropneumoniae. Vet. Microbiol. 2017, 204, 129–132. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, J.; Xu, L.; Liu, Y.; Li, P.; Zhu, T.; Cheng, C.; Lu, S.; Xu, T.; Yi, H.; et al. Spread of the florfenicol resistance floR gene among clinical Klebsiella pneumoniae isolates in China. Antimicrob. Resist. Infect. Cont. 2018, 7, 127. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Day | Culture | Group | Culturable Counts ± SD (CFU/g) | |||
|---|---|---|---|---|---|---|
| Control Cultures | Treated Cultures | |||||
| 1 | 2 | 1 | 2 | |||
| 6 | Rotifer | HB | 7.90 × 108 ± 9.16 × 107 | 5.80 × 108 ± 2.33 × 108 | N/A | N/A |
| FRB | 1.03 × 107 ± 4.94 × 106 | 8.29 × 106 ± 2.39 × 106 | N/A | N/A | ||
| Vibrio | 3.04 × 108 ± 4.48 × 107 | 1.48 × 108 ± 4.95 × 107 | N/A | N/A | ||
| 18 | Rotifer | HB | 2.21 × 108 ± 2.11 × 108 | 2.57 × 108 ± 1.37 × 108 | N/A | N/A |
| FRB | 8.03 × 104 ± 1.04 × 104 | 5.20 × 105 ± 3.74 × 105 | N/A | N/A | ||
| Vibrio | 3.29 × 107 ± 9.05 × 106 | 3.08 × 107 ± 2.08 × 107 | N/A | N/A | ||
| 18 | Artemia | HB | 4.21 × 108 ± 1.68 × 108 | 3.09 × 108 ± 2.37 × 108 | 3.81 × 108 ± 2.45 × 108 | 1.58 × 108 ± 5.21 × 107 |
| FRB | 1.57 × 107 ± 7.88 × 106 | 1.47 × 106 ± 4.88 × 104 | 3.82 × 107 ± 1.58 × 107 | 7.84 × 107 ± 1.47 × 107 | ||
| Vibrio | 1.62 × 107 ± 1.46 × 107 | 3.60 × 104 ± 2.70 × 104 | 1.82 × 107 ± 8.44 × 106 | 4.42 × 106 ± 3.08 × 106 | ||
| 32 | Rotifer | HB | 6.79 × 108 ± 9.57 × 107 | 1.71 × 109 ± 2.63 × 108 | N/A | N/A |
| FRB | 1.12 × 106 ± 9.57 × 105 | 1.79 × 107 ± 7.27 × 106 | N/A | N/A | ||
| Vibrio | 8.33 × 107 ± 3.56 × 107 | 2.63 × 108 ± 8.25 × 107 | N/A | N/A | ||
| 32 | Artemia | HB | 4.55 × 108 ± 2.98 × 108 | 1.74 × 109 ± 4.99 × 108 | 1.27 × 109 ± 3.66 × 108 | 1.21 × 109 ± 7.55 × 108 |
| FRB | 3.33 × 107 ± 2.01 × 107 | 1.00 × 108 ± 5.92 × 107 | 1.22 × 108 ± 7.18 × 107 | 3.50 × 108 ± 6.41 × 107 | ||
| Vibrio | 1.20 × 107 ± 1.38 × 107 | 3.54 × 108 ± 1.41 × 108 | 2.14 × 106 ± 1.05 × 107 | 4.02 × 107 ± 2.44 × 106 | ||
| Isolate | Source | Phenotypic Properties | Accession No. | Closest Species (% Identity) | ||||
|---|---|---|---|---|---|---|---|---|
| Gram | OF Glucose | Oxidase | O-129 | Growth on TCBS | ||||
| RVF24 | Treated culture | − | F | + | S | + | MN920721 | Vibrio azureus (100.00) |
| RVF27 | Untreated culture | − | F | + | S | + | MN920722 | Vibrio xuii (99.65) |
| RVF33 | Treated culture | − | F | + | S | + | MN920723 | Vibrio xuii (99.49) |
| RGF51 | Untreated culture | − | I | + | R | − | MN920724 | Pseudoalteromonas carrageenovora (99.49) |
| RGF65 | Treated culture | − | I | + | R | − | MN920725 | Pseudoalteromonas tetraodonis (90.81) |
| RGF67 | Treated culture | − | I | + | R | − | MN920726 | Pseudoalteromonas paragorgicola (99.04) |
| RGF70 | Treated culture | − | I | + | R | − | MN920727 | Pseudoalteromonas carrageenovora (99.15) |
| RGT71 | Untreated culture | − | F | + | S | + | MN920728 | Vibrio alginolyticus (98.97) |
| RGFR106 | Untreated culture | − | F | + | S | + | MN920729 | Vibrio xuii (98.56) |
| RGFR107 | Untreated culture | − | F | + | S | + | MN920730 | Vibrio xuii (98.25) |
| RGFR108 | Untreated culture | − | F | + | S | + | MN955424 | Vibrio xuii (98.01) |
| RGFR109 | Untreated culture | − | F | + | S | + | MN920731 | Vibrio xuii (98.60) |
| RGFR112 | Treated culture | − | F | + | S | + | MN920732 | Vibrio xuii (97.31) |
| RGFR113 | Treated culture | − | F | + | S | + | MN920733 | Vibrio xuii (97.91) |
| RGFR114 | Treated culture | − | F | + | S | + | MN920734 | Vibrio algoinfesta (97.08) |
| RGFR115 | Treated culture | − | F | + | S | + | MN920735 | Vibrio xuii (97.21) |
| RGFR116 | Treated culture | − | F | + | S | + | MN920736 | Vibrio xuii (97.95) |
| RGFR118 | Treated culture | − | F | + | S | + | MN920737 | Vibrio xuii (97.80) |
| RGFR119 | Treated culture | − | F | + | S | + | MN920738 | Vibrio xuii (97.23) |
| RGFR120 | Treated culture | − | F | + | S | + | MN920739 | Vibrio xuii (98.08) |
| RGFR121 | Treated culture | − | F | + | S | + | MN920740 | Vibrio xuii (97.65) |
| RGTR139 | Untreated culture | − | F | + | S | + | MN920741 | Vibrio xuii (99.36) |
| RGTR146 | Untreated culture | − | F | + | S | + | MN920742 | Vibrio xuii (99.48) |
| RGTR150 | Treated culture | − | F | + | S | + | MN920743 | Vibrio algoinfesta (99.78) |
| RGTR154 | Treated culture | − | F | + | S | + | MN920744 | Vibrio xuii (98.45) |
| RGTR157 | Treated culture | − | F | + | S | + | MN920745 | Vibrio xuii (98.55) |
| RGTR160 | Treated culture | − | F | + | S | + | MN920746 | Vibrio xuii (99.54) |
| Isolate | Source | Phenotypic Properties | Accession No. | Closest Species (% Identity) | ||||
|---|---|---|---|---|---|---|---|---|
| Gram | OF Glucose | Oxidase | O-129 | Growth on TCBS | ||||
| AVF09 | Untreated culture | − | F | + | S | + | MN920747 | Vibrio tasmaniensis (99.57) |
| AVF32 | Untreated culture | − | F | + | S | + | MN920748 | Vibrio alginolyticus (99.53) |
| AVF45 | Untreated culture | − | F | + | S | + | MN920749 | Vibrio toranzoniae (99.72) |
| AVF53 | Untreated culture | − | F | + | S | + | MN920750 | Vibrio rotiferianus (95.90) |
| AVF58 | Untreated culture | − | F | + | S | + | MN920751 | Vibrio rotiferianus (95.90) |
| AVF60 | Untreated culture | − | F | + | S | + | MN920752 | Vibrio neocaledonicus (100.00) |
| AVF61 | Untreated culture | − | F | + | S | + | MN920753 | Vibrio harveyi (100.00) |
| AVF68 | Untreated culture | − | F | + | S | + | MN920754 | Vibrio toranzoniae (99.14) |
| AGF75 | Untreated culture | − | F | + | S | + | MN920755 | Vibrio natriegens (98.35) |
| AGF78 | Untreated culture | − | O | + | R | − | MN920756 | Pseudomonas pachastrellae (98.14) |
| AGF82 | Untreated culture | − | F | + | S | + | MN920758 | Vibrio alginolyticus (100.00) |
| AGF83 | Untreated culture | − | F | + | S | + | MN920760 | Vibrio alginolyticus (99.90) |
| AGF85 | Untreated culture | − | F | + | S | + | MN920773 | Vibrio alginolyticus (99.89) |
| AGF86 | Untreated culture | − | F | + | S | + | MN920759 | Vibrio neocaledonicus (97,62) |
| AGF100 | Treated culture | − | F | + | S | + | MN920757 | Vibrio parahaemolyticus (100.00) |
| AGF101 | Treated culture | − | F | + | S | + | MN920774 | Vibrio natriegens (100.00) |
| AGF104 | Untreated culture | − | F | + | S | + | MN920775 | Vibrio alginolyticus (100.00) |
| AGF105 | Untreated culture | − | F | + | S | + | MN920776 | Vibrio natriegens (99.69) |
| AGT90 | Untreated culture | − | O | + | R | − | MN920761 | Pseudoalteromonas ganghwensis (99.69) |
| AGT92 | Untreated culture | − | O | + | R | − | MN920777 | Pseudoalteromonas tetraodonis (94.08) |
| AGT93 | Untreated culture | − | O | + | R | − | MN920766 | Psychrobacter pacificensis (83.80) |
| AGT94 | Untreated culture | − | O | + | R | − | MN920767 | Pseudoalteromonas atlantica (98.05) |
| AGT103 | Untreated culture | − | F | + | S | + | MN920778 | Vibrio neocaledonicus (99.90) |
| AGT104 | Untreated culture | − | O | + | R | − | MN920779 | Pseudoaleteromonas carrageenovora (85.03) |
| AGT106 | Untreated culture | − | I | + | R | − | MN955425 | Pseudoalteromonas shioyasakiensis (97.20) |
| AGT109 | Treated culture | − | F | + | S | + | MN920780 | Vibrio hyugaensis (98.30) |
| AGT110 | Treated culture | − | F | + | S | + | MN920768 | Vibrio alginolyticus (97.24) |
| AGT111 | Treated culture | − | F | + | S | + | MN920783 | Vibrio xuii (97.64) |
| AGT112 | Treated culture | − | F | + | S | + | MN920765 | Vibrio alginolyticus (99.80) |
| AGT114 | Treated culture | − | F | + | S | + | MN920764 | Vibrio neocaledonicus (99.60) |
| AGT119 | Untreated culture | − | F | + | S | + | MN920763 | Vibrio parahaemolyticus (99.30) |
| AGT122 | Treated culture | − | F | + | S | + | MN920769 | Vibrio azureus (98.29) |
| AGT123 | Treated culture | − | F | + | S | + | MN920770 | Vibrio diabolicus (99.90) |
| AGT124 | Treated culture | − | F | + | S | + | MN920771 | Vibrio alginolyticus (99.79) |
| AGT125 | Treated culture | − | F | + | S | + | MN920772 | Vibrio alginolyticus (98.65) |
| AGT127 | Treated culture | − | O | + | R | − | MN920781 | Pseudoalteromonas tetraodonis (86.82) |
| AGT128 | Treated culture | − | F | + | S | + | MN920782 | Vibrio alginolyticus (98.23) |
| AGT129 | Treated culture | − | F | + | S | + | MN920762 | Vibrio parahaemolyticus (97.71) |
| Isolate | MIC FLO (µg/mL) | Resistance Pattern | FLO Resistance Genes | No. of Plasmids | Approximate Size (kb) | |
|---|---|---|---|---|---|---|
| floR | fexA | |||||
| RVF24 | 64 | AMO-STR-KAN-FLO-OXY-OXA-FLQ | − | − | 0 | − |
| RVF27 | 64 | AMO-STR-KAN-FLO-OXY-OXA-FLQ-SXT | − | + | 0 | − |
| RVF33 | 32 | AMO-STR-KAN-FLO-OXY-OXA-FLQ-SXT | − | − | 0 | − |
| RGF51 | 256 | FLO-OXY | + | + | 0 | − |
| RGF65 | 256 | FLO-OXY-OXA-FLQ-SXT | + | + | 1 | 1.4 |
| RGF67 | 256 | FLO-OXY | − | − | 0 | − |
| RGF70 | 256 | FLO-OXA | − | − | 0 | − |
| RGT71 | 128 | AMO-STR-KAN-FLO-OXY | + | − | 2 | 40/60 |
| RGFR106 | 256 | STR-KAN-FLO-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGFR107 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | + | 0 | − |
| RGFR108 | 512 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | − | − | 0 | − |
| RGFR109 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 0 | − |
| RGFR112 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGFR113 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 0 | − |
| RGFR114 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGFR115 | 256 | AMO-STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 80 |
| RGFR116 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGFR118 | 128 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGFR119 | 256 | AMO-STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 0 | − |
| RGFR120 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 0 | − |
| RGFR121 | 512 | STR-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 35 |
| RGTR139 | 128 | STR-FLO-OXY-OXA-FLQ-SXT | + | − | 0 | − |
| RGTR146 | 512 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGTR150 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGTR154 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGTR157 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| RGTR160 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 35 |
| Isolate | MIC FLO (µg/mL) | Resistance Pattern | FLO Resistance Genes | No. of Plasmids | Approximate Size (kb) | |
|---|---|---|---|---|---|---|
| floR | fexA | |||||
| AVF09 | 32 | AMO-STR-FLO-OXY-OXA-FLQ | − | + | 1 | 40 |
| AVF32 | 32 | STR-FLO | − | − | 0 | − |
| AVF45 | 32 | STR-KAN-FLO-OXY | − | − | 3 | 5/6/15 |
| AVF53 | 32 | AMO-STR-KAN-FLO-OXY-SXT | − | − | 1 | 40 |
| AVF58 | 64 | AMO-STR-KAN-FLO-OXY-OXA-FLQ-SXT | + | − | 1 | 40 |
| AVF60 | 32 | AMO-STR-KAN-FLO-OXY-OXA-FLQ-SXT | − | − | 1 | 40 |
| AVF61 | 64 | AMO-STR-KAN-FLO-OXY-OXA-SXT | + | − | 1 | 40 |
| AVF68 | 512 | STR-KAN-FLO-OXY-OXA | − | − | 1 | 40 |
| AGF75 | 512 | AMO-STR-FLO-OXY-SXT | + | − | 2 | 35/100 |
| AGF78 | 256 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | − | + | 0 | − |
| AGF82 | 512 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 1 | 35 |
| AGF83 | 512 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 1 | 40 |
| AGF85 | 512 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 1 | 35 |
| AGF86 | 512 | AMO-STR-KAN-FLO-SXT | + | − | 1 | 50 |
| AGF100 | 256 | AMO-STR-KAN-FLO-SXT | + | − | 0 | − |
| AGF101 | 256 | AMO-STR-KAN-FLO-SXT | + | − | 1 | 35 |
| AGF104 | 256 | AMO-STR-KAN-FLO-OXA-FLQ-SXT | + | − | 1 | 35 |
| AGF105 | 1024 | AMO-STR-FLO-OXY-SXT | + | − | 2 | 35/100 |
| AGT90 | 256 | STR-KAN-FLO-OXY-SXT | − | − | 1 | 40 |
| AGT92 | 1024 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | − | − | 0 | − |
| AGT93 | 128 | AMO-STR-FLO-OXY-SXT | − | + | 1 | 40 |
| AGT94 | 1024 | AMO-STR-KAN-FLO-OXY-SXT | − | − | 1 | 50 |
| AGT103 | 512 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 1 | 35 |
| AGT104 | 128 | STR-FLO-OXY-OXA-FLQ | − | + | 1 | 50 |
| AGT106 | 256 | STR-KAN-FLO-OXY-SXT | − | − | 1 | 50 |
| AGT109 | 512 | AMO-STR-KAN-FLO-OXY-SXT | + | + | 2 | 35/100 |
| AGT110 | 128 | AMO-STR-FLO-OXY-SXT | + | − | 2 | 15/45 |
| AGT111 | 1024 | STR-KAN-FLO-OXY-OXA-FLQ-SXT | − | + | 2 | 1.3/2 |
| AGT112 | 256 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 1 | 55 |
| AGT114 | 128 | AMO-STR-FLO-OXY-SXT | + | − | 2 | 15/45 |
| AGT119 | 256 | AMO-STR-FLO-OXY-SXT | + | − | 2 | 15/45 |
| AGT122 | 512 | AMO-STR-FLO-OXY-SXT | + | − | 2 | 15/45 |
| AGT123 | 256 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 2 | 40/60 |
| AGT124 | 128 | AMO-STR-FLO-OXY-SXT | + | − | 2 | 40/60 |
| AGT125 | 256 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 1 | 50 |
| AGT127 | 512 | AMO-STR-KAN-FLO-OXY-SXT | − | − | 2 | 1/40 |
| AGT128 | 256 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 1 | 35 |
| AGT129 | 512 | AMO-STR-KAN-FLO-OXY-SXT | + | − | 1 | 35 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hurtado, L.; Miranda, C.D.; Rojas, R.; Godoy, F.A.; Añazco, M.A.; Romero, J. Live Feeds Used in the Larval Culture of Red Cusk Eel, Genypterus chilensis, Carry High Levels of Antimicrobial-Resistant Bacteria and Antibiotic-Resistance Genes (ARGs). Animals 2020, 10, 505. https://doi.org/10.3390/ani10030505
Hurtado L, Miranda CD, Rojas R, Godoy FA, Añazco MA, Romero J. Live Feeds Used in the Larval Culture of Red Cusk Eel, Genypterus chilensis, Carry High Levels of Antimicrobial-Resistant Bacteria and Antibiotic-Resistance Genes (ARGs). Animals. 2020; 10(3):505. https://doi.org/10.3390/ani10030505
Chicago/Turabian StyleHurtado, Luz, Claudio D. Miranda, Rodrigo Rojas, Félix A. Godoy, Mark A. Añazco, and Jaime Romero. 2020. "Live Feeds Used in the Larval Culture of Red Cusk Eel, Genypterus chilensis, Carry High Levels of Antimicrobial-Resistant Bacteria and Antibiotic-Resistance Genes (ARGs)" Animals 10, no. 3: 505. https://doi.org/10.3390/ani10030505
APA StyleHurtado, L., Miranda, C. D., Rojas, R., Godoy, F. A., Añazco, M. A., & Romero, J. (2020). Live Feeds Used in the Larval Culture of Red Cusk Eel, Genypterus chilensis, Carry High Levels of Antimicrobial-Resistant Bacteria and Antibiotic-Resistance Genes (ARGs). Animals, 10(3), 505. https://doi.org/10.3390/ani10030505