Endometritis Changes the Neurochemical Characteristics of the Caudal Mesenteric Ganglion Neurons Supplying the Gilt Uterus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Procedures
2.3. Immunohistochemical Analysis
2.4. Statistical Analysis
3. Results
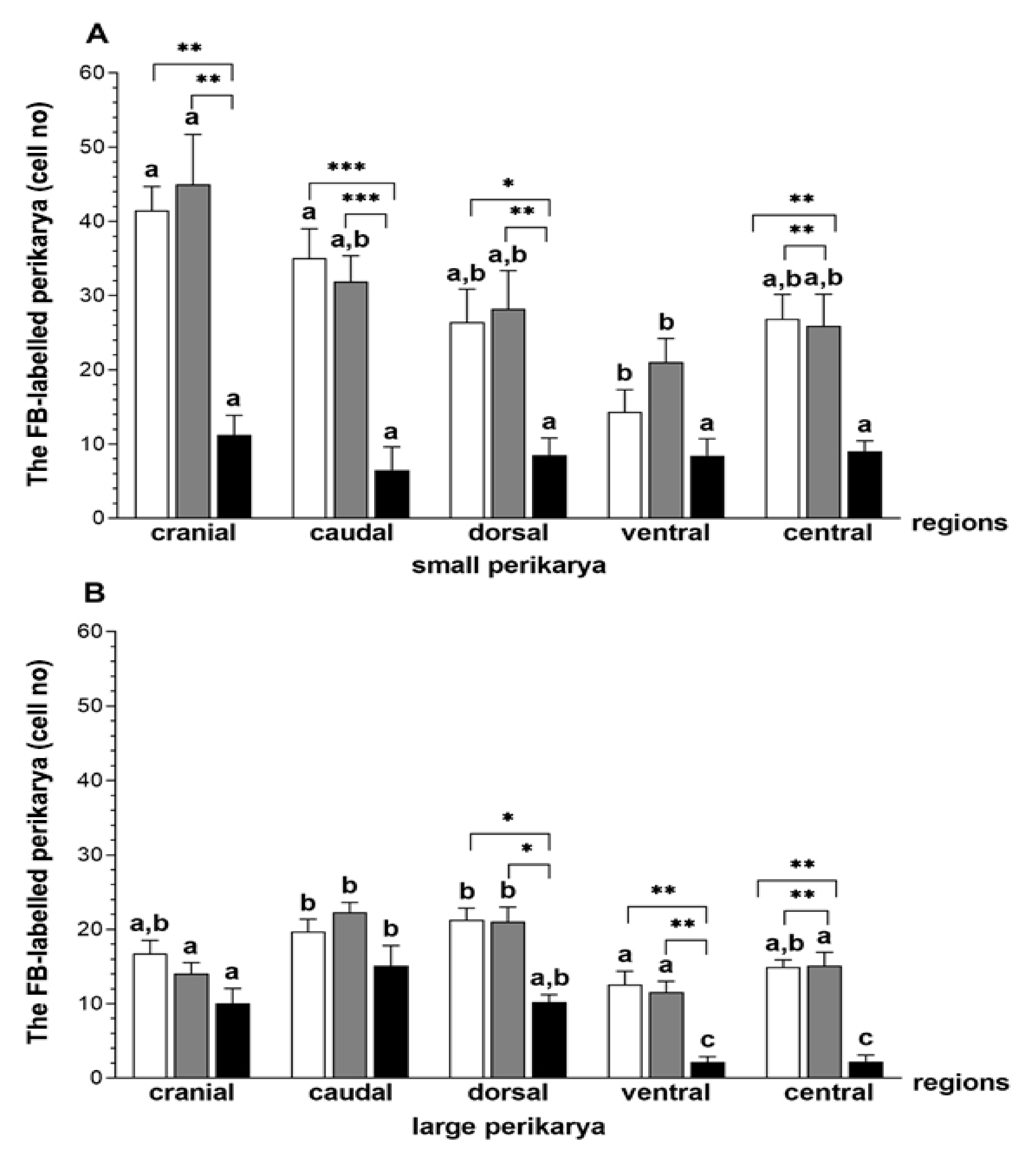
3.1. The Population and Localization of the CaMG Uterine Perikarya
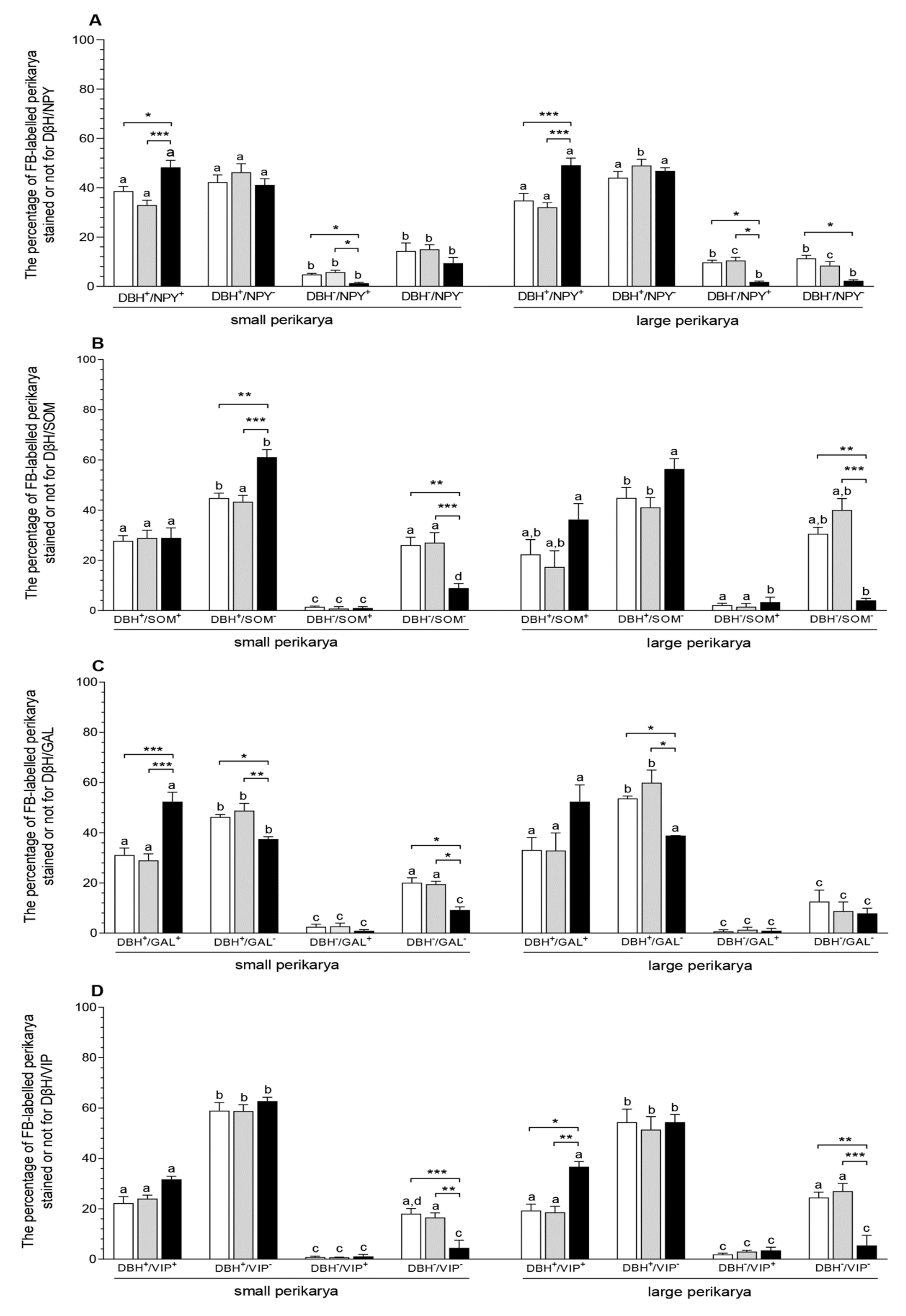
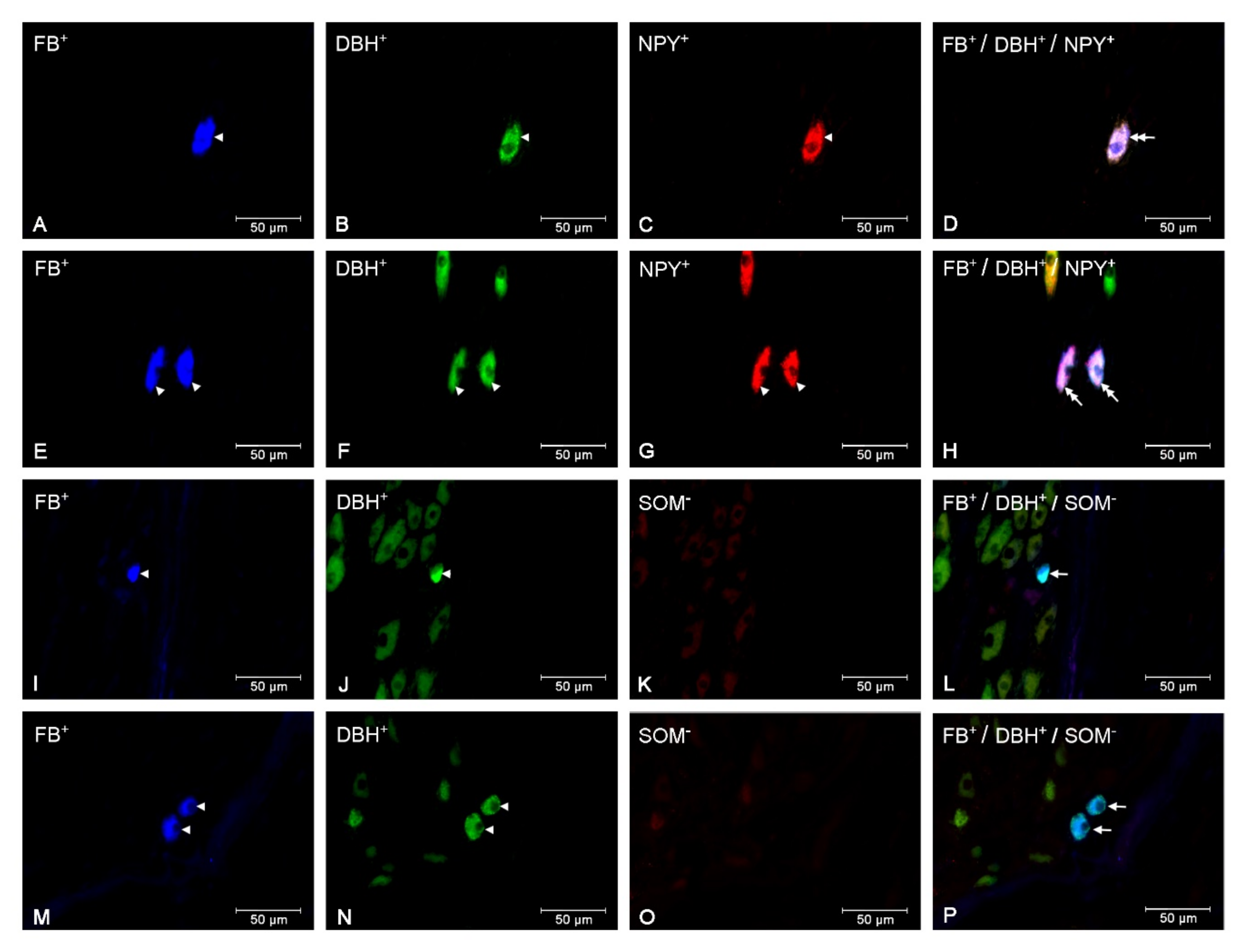
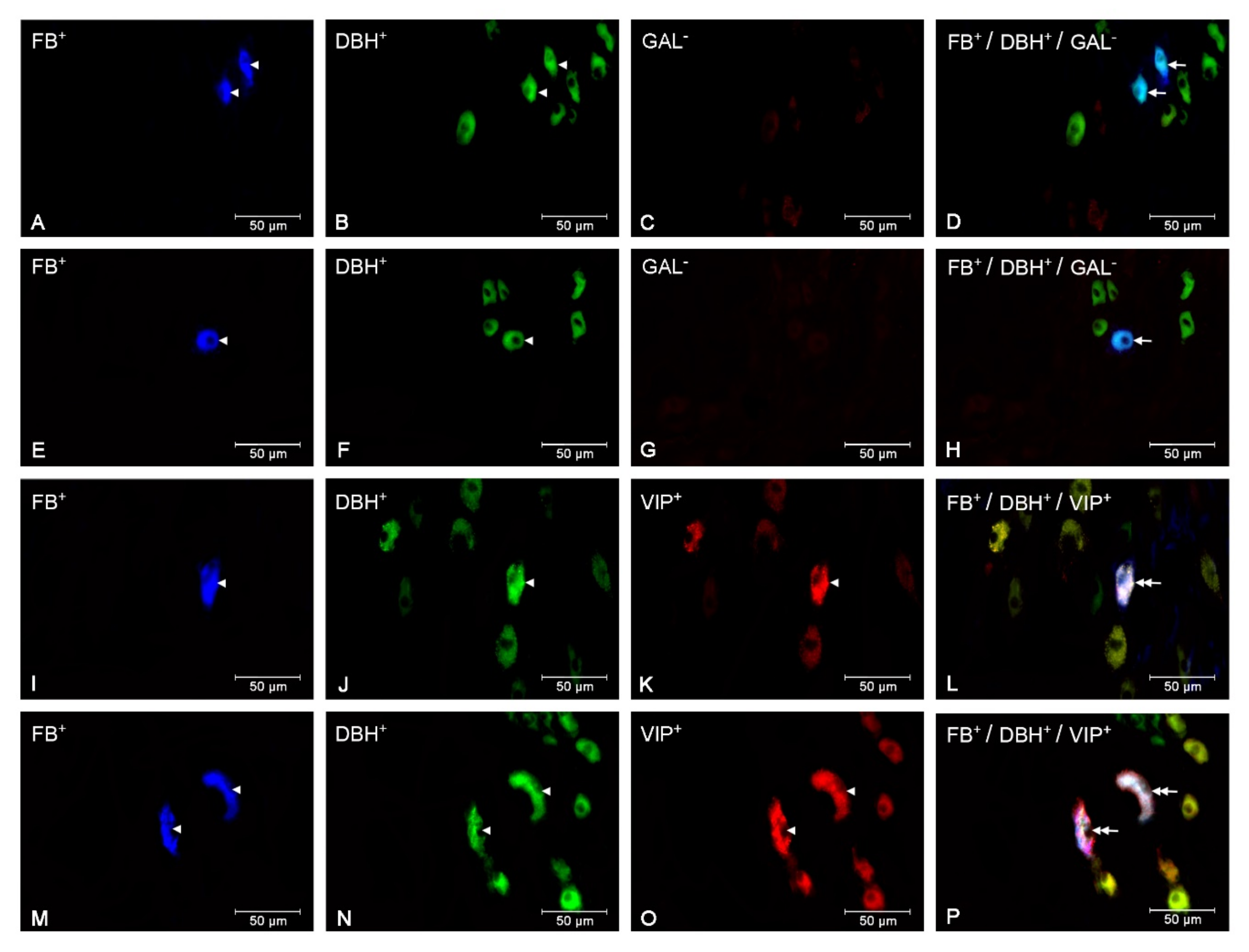
3.2. The Populations of the CaMG Uterine Perikarya Expressing DβH, NPY, SOM, GAL and VIP
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Winter, P.J.J.; Verdonck, M.; De Kruif, A.; Devriese, L.A.; Haesebrouck, R. Bacterial endometritis and vaginal discharge in the sow: Prevalence of different bacterial species and experimental reproduction of the syndrome. Anim. Reprod. Sci. 1995, 37, 325–335. [Google Scholar] [CrossRef]
- Mateus, L.; Lopes da Costa, L.; Diniz, P.; Ziecik, A.J. Relationship between endotoxin and prostaglandin (PGE2 and PGFM) concentrations and ovarian function in dairy cows with puerperal endometritis. Anim. Reprod. Sci. 2003, 76, 143–154. [Google Scholar] [CrossRef]
- Roberson, J.; Moll, D.; Saunders, G. Chronic Staphylococcus aureus endometritis in a virgin gilt. Vet. Rec. 2007, 161, 821–822. [Google Scholar] [PubMed]
- Tummaruk, P.; Kesdangsakonwut, S.; Prapasarakul, N.; Kaeoket, K. Endometritis in gilts: Reproductive data, bacterial culture, histopathology, and infiltration of immune cells in the endometrium. Comp. Clin. Pathol. 2010, 19, 575–584. [Google Scholar] [CrossRef]
- Shynlova, O.; Lee, Y.H.; Srikhajon, K.; Lye, S.J. Physiologic uterine inflammation and labor onset: Integration of endocrine and mechanical signals. Reprod. Sci. 2013, 20, 154–167. [Google Scholar] [CrossRef]
- Herath, S.; Fischer, D.P.; Werling, D.; Williams, E.J.; Lilly, S.T.; Dobson, H.; Bryant, C.E.; Sheldon, I.M. Expression and function of Toll-like receptor 4 in the endometrial cells of the uterus. Endocrinology 2006, 147, 562–570. [Google Scholar] [CrossRef]
- Gabler, C.; Drillich, M.; Fischer, C.; Holder, C.; Heuwieser, W.; Einspanier, R. Endometrial expression of selected transcripts involved in prostaglandin synthesis in cows with endometritis. Theriogenology 2009, 71, 993–1004. [Google Scholar] [CrossRef]
- Christoffersen, M.; Woodward, E.M.; Bojesen, A.M.; Petersen, M.R.; Squires, E.L.; Lehn-Jensen, H.; Troedsson, M.H. Effect of immunomodulatory therapy on the endometrial inflammatory response to induced infectious endometritis in susceptible mares. Theriogenology 2012, 78, 991–1004. [Google Scholar] [CrossRef]
- Shao, C.Y.; Wang, H.; Meng, X.; Zhu, J.Q.; Wu, Y.Q.; Li, J.J. Characterization of the innate immune response in goats after intrauterine infusion of E. coli using histopathological, cytologic and molecular analyses. Theriogenology 2012, 78, 593–604. [Google Scholar] [CrossRef]
- Jana, B.; Kucharski, J.; Dzienis, A.; Deptuła, K. Changes in prostaglandin production and ovarian function in gilts during endometritis induced by Escherichia coli infection. Anim. Reprod. Sci. 2007, 97, 137–150. [Google Scholar] [CrossRef]
- Jana, B.; Jaroszewski, J.J.; Czarzasta, J.; Włodarczyk, M.; Markiewicz, W. Synthesis of prostacyclin and its effect on the contractile activity of the inflamed porcine uterus. Theriogenology 2013, 79, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Jana, B.; Czarzasta, J.; Jaroszewski, J. Synthesis of leukotrienes in porcine uteri with endometritis induced by infection with Escherichia coli. Reprod. Fertil. Dev. 2014, 26, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Jana, B.; Andronowska, A.; Kucharski, J. Nitric oxide mediates an inflammatory effect of Escherichia coli in the porcine uterus. Polish J. Vet. Sci. 2000, 4, 207–212. [Google Scholar]
- Wasowicz, K.; Majewski, M.; Lakomy, M. Distribution of neurons innervating the uterus of the pig. J. Auton. Nerv. Syst. 1998, 74, 13–22. [Google Scholar] [CrossRef]
- Wasowicz, K. Uterus-innervating neurons in porcine inferior mesenteric ganglion: An immunohistochemical characteristic. Anat. Histol. Embryol. 2003, 32, 154–160. [Google Scholar] [CrossRef]
- Majewski, M.; Sienkiewicz, W.; Kaleczyc, J.; Mayer, B.; Czaja, K.; Lakomy, M. The distribution and co-localization of immunoreactivity to nitric oxide synthase, vasoactive intestinal polypeptide and substance P within nerve fibres supplying bovine and porcine female genital organs. Cell Tissue Res. 1995, 281, 445–464. [Google Scholar] [CrossRef]
- Rytel, L.; Gonkowski, S.; Janowski, T.; Wojtkiewicz, J.; Pomianowski, A. The neurochemical characterization of parasympathetic nerve fibres in the porcine wall under physiological conditons and after exposure to bisphenol A (BPA). Neurotox. Res. 2019, 35, 867–882. [Google Scholar]
- Kitazawa, T.; Nakagoshi, K.; Teraoka, H.; Taneike, T. 5-HT(7) receptor and beta(2)-adrenoceptor share in the inhibition of porcine uterine contractility in a muscle layer-dependent manner. Eur. J. Pharmacol. 2001, 433, 187–197. [Google Scholar] [CrossRef]
- Markiewicz, W.; Jaroszewski, J.J. β2- and β3-adrenergic receptors stimulation relaxes porcine; myometrium in the peri-implantation period. J. Anim. Sci. 2016, 94, 4611–4618. [Google Scholar] [CrossRef]
- Quaas, L.; Zahradnik, P. The effects of alpha- and beta-adrenergic stimulation on contractility and prostaglandin (prostaglandins E2 and F2a and 6-keto-prostaglandin F1 alpha) production of pregnant human myometrial strips. Am. J. Obstet. Gynecol. 1985, 152, 852–856. [Google Scholar] [CrossRef]
- Skarzynski, D.J.; Uenoyama, Y.; Kotwica, J.; Okuda, K. Noradrenaline stimulates the production of prostaglandin f2alpha in cultured bovine endometrial cells. Biol. Reprod. 1999, 60, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Atke, A.; Henriksen, J.S.; Jacobsen, H.S.; Vilhardt, H. Characterization of the rat myometrial contractile response to neuropeptide Y. Recept. Signal Transduct. Res. 1996, 16, 25–38. [Google Scholar] [CrossRef]
- Morris, J.L. Neuropeptide Y inhibits relaxations of the guinea pig uterine artery produced by VIP. Peptides 1990, 11, 381–386. [Google Scholar] [CrossRef]
- Jovanovic, S.; Grbovic, L.; Jovanovic, A. Pregnancy is associated with altered response to neuropeptide Y in uterine artery. Mol. Hum. Reprod. 2000, 6, 352–360. [Google Scholar] [CrossRef] [Green Version]
- Niiro, N.; Nishimura, J.; Hirano, K.; Nakano, H.; Kanaide, H. Mechanisms of galanin-induced contraction in the rat myometrium. Br. J. Pharmacol. 1998, 124, 1623–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryman, I.; Norström, A.; Lindblom, B.; Dahlström, A. Histochemical localization of vasoactive intestinal polypeptide and its influence on contractile activity in the non-pregnant and pregnant human cervix. Gynecol. Obstet. Investig. 1989, 28, 57–61. [Google Scholar] [CrossRef]
- Annunziata, M.; Luque, R.M.; Durán-Prado, M.; Baragli, A.; Grande, C.; Volante, M.; Gahete, M.D.; Deltetto, F.; Camanni, M.; Ghigo, E.; et al. Somatostatin and somatostatin analogues reduce PDGF-induced endometrial cell proliferation and motility. Hum. Reprod. 2012, 27, 2117–2129. [Google Scholar] [CrossRef] [Green Version]
- Wesselmann, U.; Czakanski, P.P.; Affaitati, G.; Giamberardino, M.A. Uterine inflammation as a noxious visceral stimulus: Behavioral characterization in the rat. Neurosci. Lett. 1998, 246, 73–76. [Google Scholar] [CrossRef]
- Li, J.; Micevych, P.; McDonald, J.; Rapkin, A.; Chaban, V. Inflammation in the uterus induces phosphorylated extracellular signal-regulated kinase and substance P immunoreactivity in dorsal root ganglia neurons innervating both uterus and colon in rats. J. Neurosci. Res. 2008, 86, 2746–2752. [Google Scholar] [CrossRef] [Green Version]
- Verma, N.; Rettenmeier, A.W.; Schmitz-Spanke, S. Recent advances in the use of Sus scrofa (pig) as a model system for proteomic studies. Proteomics 2011, 11, 776–793. [Google Scholar] [CrossRef]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Proc. Natl. Acad. Sci. USA 2012, 109, 16612–16617. [Google Scholar] [CrossRef] [PubMed]
- Bulc, M.; Całka, J.; Meller, K.; Jana, B. Endometritis affects chemical coding of the dorsal root ganglia neurons supplying uterus in the sexually mature gilts. Res. Vet. Sci. 2019, 124, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Meller, K.; Całka, J.; Palus, K.; Jana, B. Inflammation-induced changes in expression of DβH, SP and GAL in the porcine uterine nerve fibres. In Proceedings of the 4th Winter Workshop of the Society for Biology of Reproduction “Central and local Regulations of Reproductive Processes”, Zakopane, Poland, 3–4 February 2016. [Google Scholar]
- Majewski, M.; Heym, C. The origin of ovarian neuropeptide Y (NPY)-immunoreactive nerve fibres from the inferior mesenteric ganglion in the pig. Cell Tissue Res. 1991, 266, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Neuhuber, W.L.; Sandoz, P.A.; Fryscak, T. The central projections of primary afferent neurons of greater splanchnic and intercostal nerves in the rat. A horseradish peroxidase study. Anat. Embryol. 1986, 174, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Peter, A.T.; Bosu, W.T.K. Effects of intrauterine infection on the function of the corpora lutea formed after first postpartum ovulations in dairy cows. Theriogenology 1987, 127, 593–609. [Google Scholar] [CrossRef]
- Opsomer, G.; Gröhn, Y.T.; Hertl, J.; Coryn, M.; Deluyker, H.; de Kruif, A. Risk factors for post-partum ovarian dysfunction in high producing dairy cows in Belgium: A field study. Theriogenology 2000, 53, 841–857. [Google Scholar] [CrossRef]
- Jana, B.; Kucharski, J.; Ziecik, A.J. Effect of intrauterine infusion of Escherichia coli on hormonal patterns in gilts during the oestrous cycle. Reprod. Nutr. Dev. 2004, 44, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Green, P.S.; Bishop, J.; Simpkins, J.W. 17 alpha-estradiol exerts neuroprotective effects on SK-N-SH cells. J. Neurosci. 1997, 17, 511–515. [Google Scholar]
- Cardona-Gómez, G.P.; Mendez, P.; DonCarlos, L.L.; Azcoitia, I.; Garcia-Segura, L.M. Interactions of estrogens and insulin-like growth factor-I in the brain: Implications for neuroprotection. Brain Res. Brain Res. Rev. 2001, 37, 320–334. [Google Scholar] [CrossRef]
- Papka, R.E.; Workley, M.; Usip, S.; Mowa, C.N.; Fahrenkrug, J. Expression of pituitary adenylate cyclase activating peptide in the uterine cervix, lumbosacral dorsal root ganglia and spinal cord of rats during pregnancy. Peptides 2006, 27, 743–752. [Google Scholar] [CrossRef]
- Koszykowska, M.; Całka, J.; Szwajca, P.; Jana, B. Long-term estradiol-17β administration decreases the number of neurons in the caudal mesenteric ganglion innervating the ovary in sexually mature gilts. J. Reprod. Dev. 2011, 57, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jana, B.; Rytel, L.; Czarzasta, J.; Całka, J. Reduction of the number of neurones in the caudal mesenteric ganglion innervating the ovary in sexually mature gilts following testosterone administration. J. Neuroendocrinol. 2013, 25, 826–838. [Google Scholar] [CrossRef] [PubMed]
- Pieretti, S.; Mastriota, M.; Tucci, P.; Battaglia, G.; Trabace, L.; Nicoletti, F.; Scaccianoce, S. Brain nerve growth factor unbalance induced by anabolic androgenic steroids in rats. Med. Sci. Sports Exerc. 2013, 45, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Harada, N.; Okajima, K. Dihydrotestosterone inhibits hair growth in mice by inhibiting insulin-like growth factor-I production in dermal papillae. Growth Horm. IGF Res. 2011, 21, 260–267. [Google Scholar] [CrossRef]
- Andoh, T.; Kuraishi, Y. Expression of BLT1 leukotriene B4 receptor on the dorsal root ganglion neurons in mice. Brain Res. Mol. Brain Res. 2005, 137, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Okubo, M.; Yamanaka, H.; Kobayashi, K.; Fukuoka, T.; Dai, Y.; Noguchi, K. Expression of leukotriene receptors in the rat dorsal root ganglion and the effects on pain behaviors. Mol. Pain 2010, 17, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, B.D.; Ho, C.; Kuzin, I.; Bottaro, A.; O’Leary, M.E. Chronic exposure to tumor necrosis factor in vivo induces hyperalgesia, upregulates sodium channel gene expression and alters the cellular electrophysiology of dorsal root ganglion neurons. Neurosci. Lett. 2017, 13, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Peterson, P.K.; Chao, C.C. Cytokine-mediated neuronal apoptosis. Neurochem. Int. 1997, 30, 427–431. [Google Scholar] [CrossRef]
- Wojtkiewicz, J.; Równiak, M.; Crayton, R.; Barczewska, M.; Bladowski, M.; Robak, A.; Pidsudko, Z.; Majewski, M. Inflammation-induced changes in the chemical coding pattern of colon-projecting neurons in the inferior mesenteric ganglia of the pig. J. Mol. Neurosci. 2012, 46, 450–458. [Google Scholar] [CrossRef]
- Zoubina, E.V.; Smith, P.G. Distributions of estrogen receptors alpha and beta in sympathetic neurons of female rats: Enriched expression by uterine innervation. J. Neurobiol. 2002, 52, 14–23. [Google Scholar] [CrossRef]
- Wise, H. Lack of interaction between prostaglandin E2 receptor subtypes in regulating adenylyl cyclase activity in cultured rat dorsal root ganglion cells. Eur. J. Pharmacol. 2006, 27, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Sabban, E.L.; Maharjan, S.; Nostramo, R.; Serova, L.I. Divergent effects of estradiol on gene expression of catecholamine biosynthetic enzymes. Physiol. Behav. 2010, 9, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Mowa, C.N.; Papka, R.E. The role of sensory neurons in cervical ripening: Effects of estrogen and neuropeptides. J. Histochem. Cytochem. 2004, 52, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Theodorsson, A.; Holm, L.; Theodorsson, E. Hypothermia-induced increase in galanin concentrations and ischemic neuroprotection in the rat brain. Neuropeptides 2008, 1, 79–87. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jana, B.; Całka, J. Endometritis Changes the Neurochemical Characteristics of the Caudal Mesenteric Ganglion Neurons Supplying the Gilt Uterus. Animals 2020, 10, 891. https://doi.org/10.3390/ani10050891
Jana B, Całka J. Endometritis Changes the Neurochemical Characteristics of the Caudal Mesenteric Ganglion Neurons Supplying the Gilt Uterus. Animals. 2020; 10(5):891. https://doi.org/10.3390/ani10050891
Chicago/Turabian StyleJana, Barbara, and Jarosław Całka. 2020. "Endometritis Changes the Neurochemical Characteristics of the Caudal Mesenteric Ganglion Neurons Supplying the Gilt Uterus" Animals 10, no. 5: 891. https://doi.org/10.3390/ani10050891
APA StyleJana, B., & Całka, J. (2020). Endometritis Changes the Neurochemical Characteristics of the Caudal Mesenteric Ganglion Neurons Supplying the Gilt Uterus. Animals, 10(5), 891. https://doi.org/10.3390/ani10050891