Influence of Different Feed Physical Forms on Mandibular Gland in Growing Pigs

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dietary Treatments and Sampling Collection
2.2. Immuno- and Glycohistochemical Treatments
2.3. Statistical Analyses
3. Results
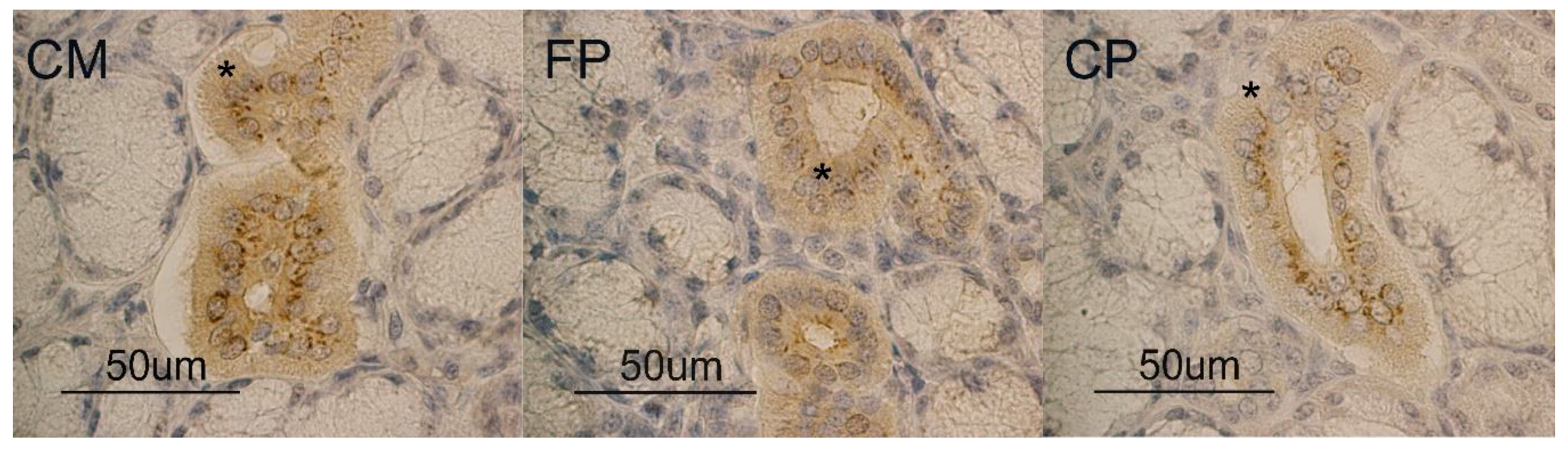
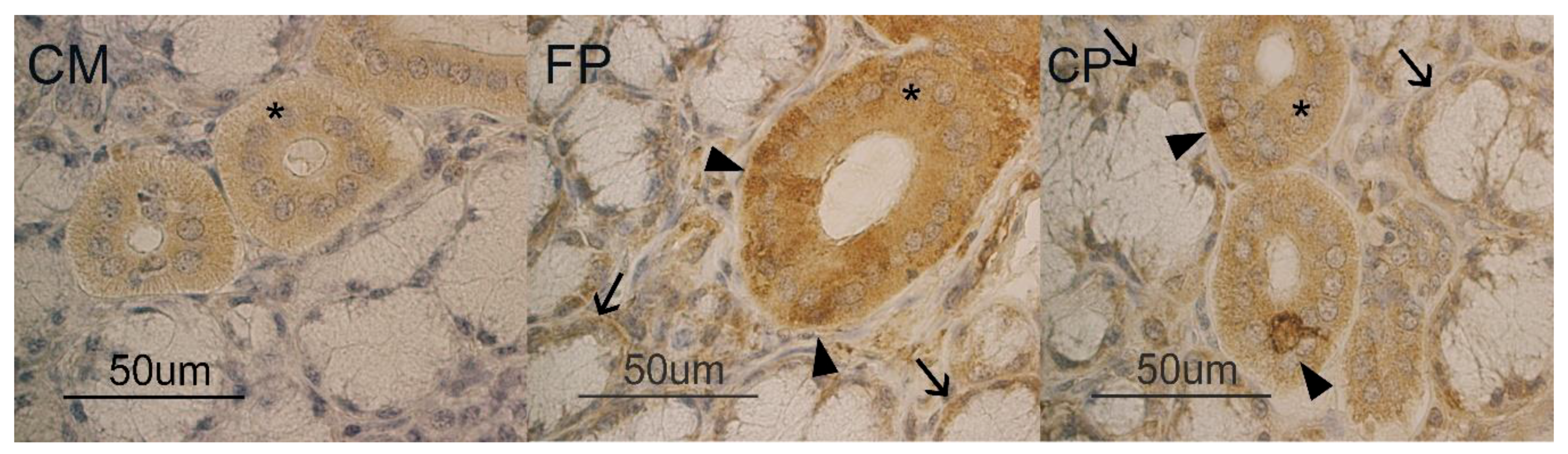
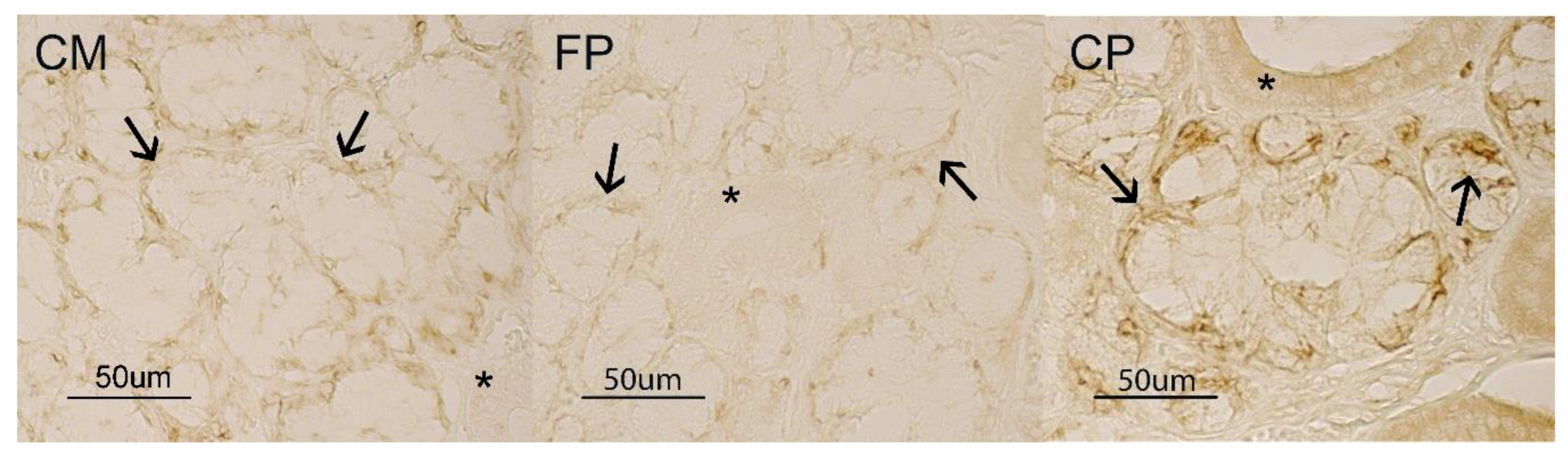
3.1. Immunohistochemistry
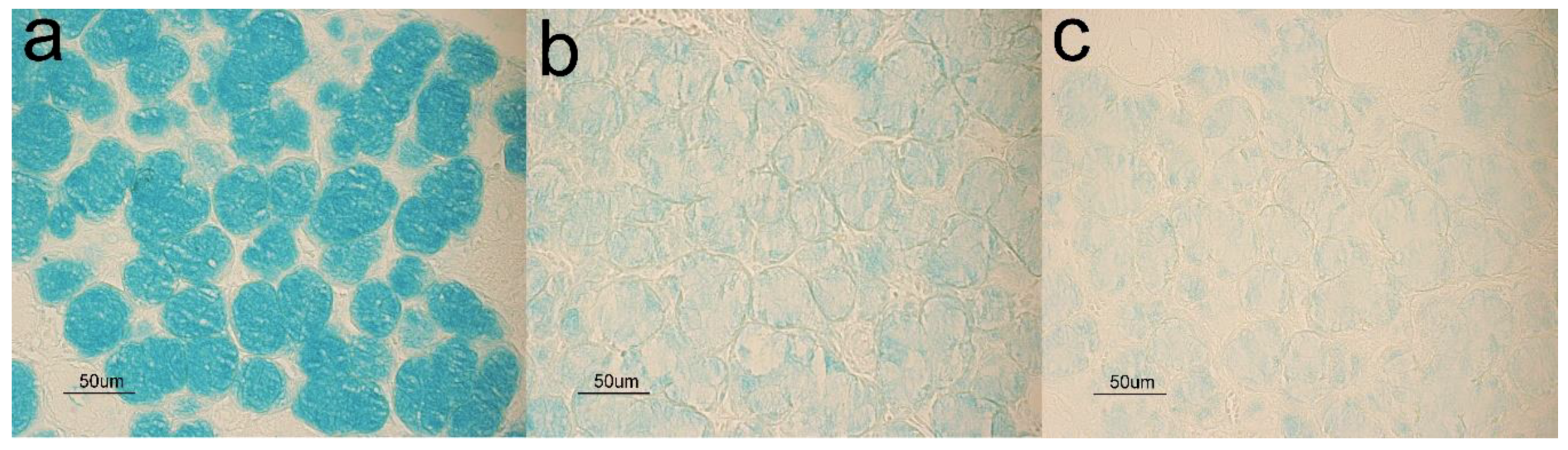
3.2. Glycohistochemistry
3.3. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cappai, M.G.; Dall’Aglio, C.; Sander, S.J.; Ratert, C.; Dimauro, C.; Pinna, W.; Kamphues, J. Different physical forms of one diet fed to growing pigs induce morphological changes in mandibular glands and local leptin (Ob) production and receptor (ObR) expression. J. Anim. Physiol. Anim. Nutr. 2016, 100, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Pirino, C.; Cappai, M.G.; Maranesi, M.; Tomassoni, D.; Giontella, A.; Pinna, W.; Boiti, C.; Kamphues, J.; Dall’Aglio, C. The presence and distribution of cannabinoid type 1 and 2 receptors in the mandibular gland: The influence of different physical forms of diets on their expression in piglets. J. Anim. Physiol. Anim. Nutr. 2018, 102, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Ito, k. The effect of food consistency and dehydration on reflex parotid and submandibular salivary secretion in conscious rats. Arch. Oral. Biol. 2001, 46, 353–363. [Google Scholar] [CrossRef]
- Johnson, D.A. Effect of a ground versus a pelleted bulk diet on the rat parotid gland. Arch. Oral Biol. 1981, 26, 1091–1093. [Google Scholar] [CrossRef]
- Johnson, D.A.; Cardenas, H. Effects of food mastication on rat parotid gland adrenergic and cholinergic cell surface receptors. Crit. Rev. Oral Biol. Med. 1993, 4, 591–597. [Google Scholar] [CrossRef]
- Kurahashi, M. The effect of dietary consistency and water content on the parotid glands of submandibular and sublingual duct-ligated rats. Arch. Oral Biol. 2002, 47, 369–374. [Google Scholar] [CrossRef]
- Johnson, D.A. Effect of a liquid diet on the protein composition of rat parotid saliva. J. Nutr. 1982, 112, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.A.; Lopez, H.; Navia, J.M. Effects of protein deficiency and diet consistency on the parotid gland and parotid saliva of rats. J. Dent. Res. 1995, 74, 1444–1452. [Google Scholar] [CrossRef]
- Tatemoto, K.; Hosoya, M.; Habata, Y.; Fujii, R.; Kakegawa, T.; Zou, M.X.; Kawamata, Y.; Fukusumi, S.; Hinuma, S.; Kitada, C.; et al. Isolation and characterization of a novel endogenous peptide ligand for the human APJ receptor. Biochem. Biophys. Res. Commun. 1998, 251, 471–476. [Google Scholar] [CrossRef]
- Kawamata, Y.; Habata, Y.; Fukusumi, S.; Hosoya, M.; Fujii, R.; Hinuma, S.; Nishizawa, N.; Kitada, C.; Onda, H.; Nishimura, O.; et al. Molecular properties of apelin: Tissue distribution and receptor binding. Biochim. Biophys. Acta 2001, 1538, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Kleinz, M.J.; Skepper, J.N.; Davenport, A.P. Immunocytochemical localisation of the apelin receptor, APJ, to human cardiomyocytes, vascular smooth muscle and endothelial cells. Regul. Pept. 2005, 126, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Medhurst, A.D.; Jennings, C.A.; Robbins, M.J.; Davis, R.P.; Ellis, C.; Winborn, K.Y.; Hervieu, G.; Riley, G.; Bolaky, J.E.; Herrity, N.C.; et al. Pharmacological and immunohistochemical characterization of the APJ receptor and its endogenous ligand apelin. J. Neurochem. 2003, 84, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- O’ Carroll, A.M.; Selby, T.L.; Palkovits, M.; Lolait, S.J. Distribution of mRNA encoding B78/apj, the rat homologue of the human APJ receptor, and its endogenous ligand apelin in brain and peripheral tissues. Biochim. Biophys. Acta 2000, 1492, 72–80. [Google Scholar] [CrossRef]
- Mercati, F.; Maranesi, M.; Dall’Aglio, C.; Petrucci, L.; Pasquariello, R.; Tardella, F.M.; De Felice, E.; Scocco, P. Apelin system in mammary gland of sheep reared in semi-natural pastures of the central apennines. Animals 2018, 8, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercati, F.; Scocco, P.; Maranesi, M.; Acuti, G.; Petrucci, L.; Cocci, P.; Renzi, A.; De Felice, E.; Dall’Aglio, C. Apelin system detection in the reproductive apparatus of ewes grazing on semi-natural pasture. Theriogenology 2019, 139, 156–166. [Google Scholar] [CrossRef]
- Nizam, M.; Basoglu, O.K.; Tasbakan, M.S.; Lappin, D.F.; Buduneli, N. Is there an association between obstructive sleep apnea syndrome and periodontal inflammation? Clin. Oral Investig. 2015, 20, 659–668. [Google Scholar] [CrossRef]
- Scocco, P.; Menghi, G.; Ceccarelli, P.; Pedini, V. Lectin histochemistry and identification of O-acetilated sialoderivatives in the horse sublingual gland. Eur. J. Histochem. 1999, 43, 47–54. [Google Scholar]
- Lynge Pedersen, A.M.; Belstrøm, D. The role of natural salivary defences in maintaining a healthy oral microbiota. J. Dent. 2019, 80, S3–S12. [Google Scholar] [CrossRef]
- Nagahama, M.; Ma, N.; Semba, R.; Naruse, S. Aquaporin 1 immunoreactive enteric neurons in the rat ileum. Neurosci. Lett. 2006, 395, 206–210. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Cho, G.; Yuan, Z.; Skowronski, M.T.; Pan, Y.; Ishida, H. Water channels and zymogen granules in salivary glands. J. Pharmacol. Sci. 2006, 100, 495–512. [Google Scholar] [CrossRef] [Green Version]
- Krane, C.M.; Melvin, J.E.; Nguyen, H.V.; Richardson, L.; Towne, J.E.; Doetschman, T.; Menon, A.G. Salivary acinar cells from aquaporin 5-deficient mice have decreased membrane water permeability and altered cell volume regulation. J. Biol. Chem. 2001, 276, 23413–23420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawedia, J.D.; Nieman, M.L.; Boivin, G.P.; Melvin, J.E.; Kikuchi, K.I.; Hand, A.R.; Lorenz, J.N.; Menon, A.G. Interaction between transcellular and paracellular water transport pathways through Aquaporin 5 and the tight junction complex. Proc. Natl. Acad. Sci. USA 2007, 104, 3621–3626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.S.; Preston, G.M.; Smith, B.L.; Guggino, W.B.; Agre, P. Molecular structure of the water channel through aquaporin CHIP. The hourglass model. J. Biol. Chem. 1994, 269, 14648–14654. [Google Scholar] [PubMed]
- Murata, K.; Mitsuoka, K.; Hirai, T.; Walz, T.; Agre, P.; Heymann, J.B.; Engel, A.; Fujiyoshi, Y. Structural determinants of permeation through aquaporin-1. Nature 2000, 407, 599–605. [Google Scholar] [CrossRef]
- Sui, H.; Han, B.G.; Lee, K.J.; Walian, P.; Jap, B.K. Structural basis of water-specific transport through the AQP1 water channel. Nature 2001, 414, 872–878. [Google Scholar] [CrossRef] [Green Version]
- Agre, P.; King, L.S.; Yasui, M.; Guggino, W.B.; Ottersen, O.P.; Fujiyoshi, Y.; Engel, A.; Nielsen, S. Aquaporin water channels— from atomic structure to clinical medicine. J. Physiol. 2002, 542, 3–16. [Google Scholar] [CrossRef]
- Yasui, M. Molecular mechanisms and drug development in aquaporin water channel diseases: Structure and function of aquaporins. J.Pharmacol. Sci. 2004, 96, 260–263. [Google Scholar] [CrossRef] [Green Version]
- Dall’Aglio, C.; Scocco, P.; Maranesi, M.; Petrucci, L.; Acuti, G.; De Felice, E.; Mercati, F. Immunohistocehmical identification of resistin in the uterus of ewes subjected to different diets: Preliminary results. Europ. J. Histochem. 2019, 63, 3020. [Google Scholar]
- Pappalardo, A.; Porreca, I.; Caputi, L.; De Felice, E.; Schulte-Merker, S.; Zannini, M.; Sordino, P. Thyroid development in zebrafish lacking Taz. Mech. Dev. 2015, 138, 268–278. [Google Scholar] [CrossRef]
- Scocco, P.; Aralla, M.; Catorci, A.; Belardinelli, C.; Arrighi, S. Immunodetection of aquaporin 5 in sheep salivary glands related to pasture vegetative cycle. Folia Histochem. Cytobiol. 2011, 49, 458–464. [Google Scholar] [CrossRef] [Green Version]
- Dall’Aglio, C.; Mercati, F.; Faeti, V.; Acuti, G.; Trabalza-Marinucci, M.; De Felice, E.; Tardella, F.M.; Franciosini, M.P.; Casagrande Proietti, P.; Catorci, D.; et al. Immuno- and glyco-histochemistry as a tool to evaluate the oregano supplemented feed effects in pig gut. Eur. J. Histochem. 2020, 64, 3110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall’Aglio, C.; Pascucci, L.; Mercati, F.; Boiti, C.; Ceccarelli, P. Localization of the orexin system in the gastrointestinal tract of fallow deer. Acta Histochem. 2012, 114, 74–78. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.R-project.org/ (accessed on 29 February 2020).
- Mercati, F.; Dall’Aglio, C.; Acuti, G.; Faeti, V.; Tardella, F.M.; Pirino, C.; De Felice, E.; Scocco, P. Oregano feed supplementation affects glycoconjugates production in swine gut. Animals 2020, 10, 149. [Google Scholar] [CrossRef] [Green Version]
- Scocco, P.; Rivaroli, S.; Mercati, F.; Tardella, F.M.; Malfatti, A.; De Felice, E.; Catorci, A. Anatomy for economy: Starting from the rumen keratinization degree to enhance the farm income. Econ. Agro-Aliment. 2018, 20, 261–272. [Google Scholar] [CrossRef]
- Domeneghini, C.; Arrighi, S.; Radaelli, G.; Bosi, G.; Veggetti, A. Histochemical analysis of glycoconjugate secretion in the alimentary canal of Anguilla anguilla L. Acta Histochem. 2005, 106, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Scocco, P.; Pedini, V. Histochemical characterisation of complex carbohydrates expressed in the alimentary tract of chickens. Vet. J. 2010, 185, 228–230. [Google Scholar] [CrossRef]
- Schulte, B.A.; Spicer, S.S. Light microscopic detection of sugar residues in glycoconjugates of salivary glands and the pancreas with lectin-horseradish peroxidase conjugates. I. Mouse. Histochem. J. 1983, 15, 1217–1238. [Google Scholar] [CrossRef]
- Menghi, G.; Bondi, A.M.; Accili, D.; Materazzi, G. Visualization of carbohydrate chains in rabbit salivary glands by means of enzymatic degradation and plant lectins. Acta Histochem. 1988, 84, 163–177. [Google Scholar] [CrossRef]
- Scocco, P.; Pedini, V. Localization of influenza virus sialoreceptors in equine respiratory tract. Histol. Histopathol. 2008, 23, 973–978. [Google Scholar]
- Pedini, V.; Dall’Aglio, C.; Parillo, F.; Scocco, P. Glycoconjugate distribution in gastric fundic mucosa of Umbrina cirrosa L. revealed by lectin histochemistry. J. Fish. Biol. 2005, 66, 222–229. [Google Scholar] [CrossRef]
- Nieuw-Amerongen, A.V.; Bolscher, J.G.M.; Veerman, E.C.I. Salivary mucins: Protective functions in relation to their diversity. Glycobiology 1995, 5, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Scocco, P.; Pedini, V. Equine mandibular gland: In situ characterization of sialoderivatives. Equine Vet. J. 2006, 38, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Sugiya, H.; Matsuki-Fukushima, M.; Hashimoto, S. Role of aquaporins and regulation of secretory vesicle volume in cell secretion. J. Cell. Mol. Med. 2008, 12, 1486–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lago, F.; Dieguez, C.; Gómez-Reino, J.; Gualillo, O. The emerging role of adipokines as mediators of inflammation and immune responses. Cytokine Growth Factor Rev. 2007, 8, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Kälin, R.E.; Kretz, M.P.; Meyer, A.M.; Kispert, A.; Heppner, F.L.; Brändli, A.W. Paracrine and autocrine mechanisms of apelin signalling govern embryonic and tumor angiogenesis. Dev. Biol. 2007, 305, 599–614. [Google Scholar] [CrossRef] [Green Version]
- Masri, B.; Knibiehler, B.; Audigier, Y. Apelin signalling: A promising pathway from cloning to pharmacology. Cell Signal. 2005, 17, 415–426. [Google Scholar] [CrossRef]
- Reaux-Le Goazigo, A.; Iturrioz, X.; Llorens-Cortens, C. Encyclopedia of Neuroscience; Elsevier: Amsterdam, NL, USA, 2009; pp. 509–516. [Google Scholar]
- Wang, G.; Anini, Y.; Wei, W.; Qi, X.; O’Carroll, A.M.; Mochizuki, T.; Wang, H.Q.; Hellmich, M.R.; Englander, E.W.; Greeley, G.H., Jr. Apelin, a new enteric peptide: Localization in the gastrointestinal tract, ontogeny, and stimulation of gastric cell proliferation and of cholecystokinin secretion. Endocrinol. 2004, 145, 1342–1348. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Wang, G.; Qi, X.; Lee, H.M.; Englander, E.W.; Greeley, G.H., Jr. A possible role for hypoxia-induced apelin expression in enteric cell proliferation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1832–R1839. [Google Scholar] [CrossRef] [Green Version]
- Rakonczay, Z., Jr.; Vág, J.; Földes, A.; Nagy, K.; Nagy, Á.; Hegyi, P.; Varga, G. Chronic inflammation in the pancreas and salivary glands-lessons from similarities and differences in pathophysiology and treatment modalities. Curr. Pharm. Des. 2014, 20, 1104–1120. [Google Scholar]
- Kapica, M.; Puzio, I.; Kato, I.; Kuwahara, A.; Zabielski, R. Role of feed-regulating peptides on pancreatic exocrine secretion. J. Physiol. Pharmacol. 2008, 59, 145–159. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Secretory Structures | Diet Groups | ||
|---|---|---|---|---|
| CM | FP | CP | ||
| APLN | Acini | − | − | − |
| Demilunes | − | − | − | |
| Ducts | + | ±/+ | ±/+ | |
| APLNR | Acini | − | − | − |
| Demilunes | − | + | + | |
| Ducts a | ± | + | + | |
| Ducts b | − | ++ | ++ | |
| APQ5 | Acini | − | − | − |
| Demilunes | ± | ± | + | |
| Ducts | − | − | ± | |
| Glycohistochemical Treatments | Secretory Structures | Diet Groups | ||
|---|---|---|---|---|
| CM | FP | CP | ||
| AB pH2.5 | Acini | +/++ | ++ | ++ |
| Demilunes | −/± | ±/+ | ± | |
| Ducts | − | − | − | |
| AB pH1 | Acini | ±/+ | ±/+ | ±/+ |
| Demilunes | − | − | − | |
| Ducts | − | − | − | |
| AB pH0.5 | Acini | − | − | −/± |
| Demilunes | − | − | − | |
| Ducts | − | − | − | |
| PAS | Acini | ++ | ++ | +++ |
| Demilunes | ± | ± | ± | |
| Ducts | − | − | − | |
| AB/PAS | Acini | B+/R++ | B++/R++ | B+++/R++ |
| Demilunes | B+/R± | B±/R± | B+/R± | |
| Ducts | − | − | − | |
| Histochemical Treatments | ANOVA/Kruskal–Wallis Test | t-test/Wilcoxon–Mann–Whitney Test | |||
|---|---|---|---|---|---|
| P CM vs. FP | P FP vs. CP | P CM vs. CP | |||
| APLN | Acini | − | − | − | − |
| Demilunes | − | − | − | − | |
| Ducts | 4.26 × 10−11 | 1.76 × 10−7 | 1.00 | 1.76 × 10−7 | |
| APLNR | Acini | − | − | − | − |
| Demilunes | 1.92 × 10−6 | 5.50 × 10−6 | 1.00 | 5.50 × 10−6 | |
| Ducts a | <10−16 | 8.36 × 10−13 | 1.00 | 1.61 × 10−12 | |
| Ducts b | 1.60 × 10−6 | 5.50 × 10−6 | 1.00 | 5.50 × 10−6 | |
| APQ5 | Acini | − | − | − | − |
| Demilunes | 3.16 × 10−6 | 1.00 | 1.92 × 10−5 | 1.92 × 10−5 | |
| Ducts | 6.57 × 10−9 | − | 5.50 × 10−6 | 5.50 × 10−6 | |
| AB pH2.5 | Acini | 3.16 × 10−6 | 1.92 × 10−5 | 1.00 | 1.92 × 10−5 |
| Demilunes | 2.33 × 10−8 | 1.80 × 10−5 | 1.90 × 10−5 | 1.92 × 10−5 | |
| Ducts | − | − | − | − | |
| AB pH1 | Acini | 1.00 | − | − | − |
| Demilunes | − | − | − | − | |
| Ducts | − | − | − | − | |
| AB pH0.5 | Acini | 6.12 × 10−9 | − | 5.28 × 10−6 | 5.28 × 10−6 |
| Demilunes | − | − | − | − | |
| Ducts | − | − | − | − | |
| PAS | Acini | <10−16 | 1.00 | <10−16 | <10−16 |
| Demilunes | 1.00 | − | − | − | |
| Ducts | − | − | − | − | |
| AB/AS | Acini | <10−16 | 1.08 × 10−13 | 4.08 × 10−14 | <10−16 |
| Demilunes | 1.03 × 10−12 | 1.24 × 10−9 | 1.05 × 10−10 | 1.00 | |
| Ducts | − | − | − | − | |
| Diet Groups | Secretory Structures | Wilcoxon Signed-Rank Test | |
|---|---|---|---|
| AB pH 2.5 vs. AB pH 1 | AB pH 1 vs. AB pH 0.5 | ||
| CM | Acini | 0.0035 | 0.0035 |
| Demilunes | 0.0035 | − | |
| Ducts | − | − | |
| FP | Acini | 0.0035 | 0.0035 |
| Demilunes | 0.0035 | − | |
| Ducts | − | − | |
| CP | Acini | 0.0035 | 0.0035 |
| Demilunes | 0.0035 | − | |
| Ducts | − | − | |
| Secretory Structures | CM | FP | CP |
|---|---|---|---|
| Acini | Hyaluronic acid and/or Chondroitin-like GAGs Chondroitin sulfate A/B/C-like GAGs | Hyaluronic acid and/or Chondroitin-like GAGs Chondroitin sulfate A/B/C-like GAGs | Hyaluronic acid and/or Chondroitin-like GAGs Chondroitin sulfate A/B/C-like GAGs Heparin and/or heparan-sulfate-like GAGs |
| Demilunes | Neutral and acid glycoproteins | Neutral and acid glycoproteins | Acid glycoproteins |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dall’Aglio, C.; Mercati, F.; De Felice, E.; Tardella, F.M.; Kamphues, J.; Cappai, M.G.; Scocco, P. Influence of Different Feed Physical Forms on Mandibular Gland in Growing Pigs. Animals 2020, 10, 910. https://doi.org/10.3390/ani10050910
Dall’Aglio C, Mercati F, De Felice E, Tardella FM, Kamphues J, Cappai MG, Scocco P. Influence of Different Feed Physical Forms on Mandibular Gland in Growing Pigs. Animals. 2020; 10(5):910. https://doi.org/10.3390/ani10050910
Chicago/Turabian StyleDall’Aglio, Cecilia, Francesca Mercati, Elena De Felice, Federico Maria Tardella, Josef Kamphues, Maria Grazia Cappai, and Paola Scocco. 2020. "Influence of Different Feed Physical Forms on Mandibular Gland in Growing Pigs" Animals 10, no. 5: 910. https://doi.org/10.3390/ani10050910
APA StyleDall’Aglio, C., Mercati, F., De Felice, E., Tardella, F. M., Kamphues, J., Cappai, M. G., & Scocco, P. (2020). Influence of Different Feed Physical Forms on Mandibular Gland in Growing Pigs. Animals, 10(5), 910. https://doi.org/10.3390/ani10050910