Immunohistochemical Analysis of Intestinal and Central Nervous System Morphology in an Obese Animal Model (Danio rerio) Treated with 3,5-T2: A Possible Farm Management Practice?


 ,
,  , ,
, , 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Zebrafish Feeding and Treatment
2.3. Body Weight Measurement
2.4. Serum Lipid Parameters
2.5. Morphological Analysis
2.6. Single Immunohistochemistry
2.7. Immunofluorescence
2.8. Controls
2.9. Statistical Analysis
3. Results
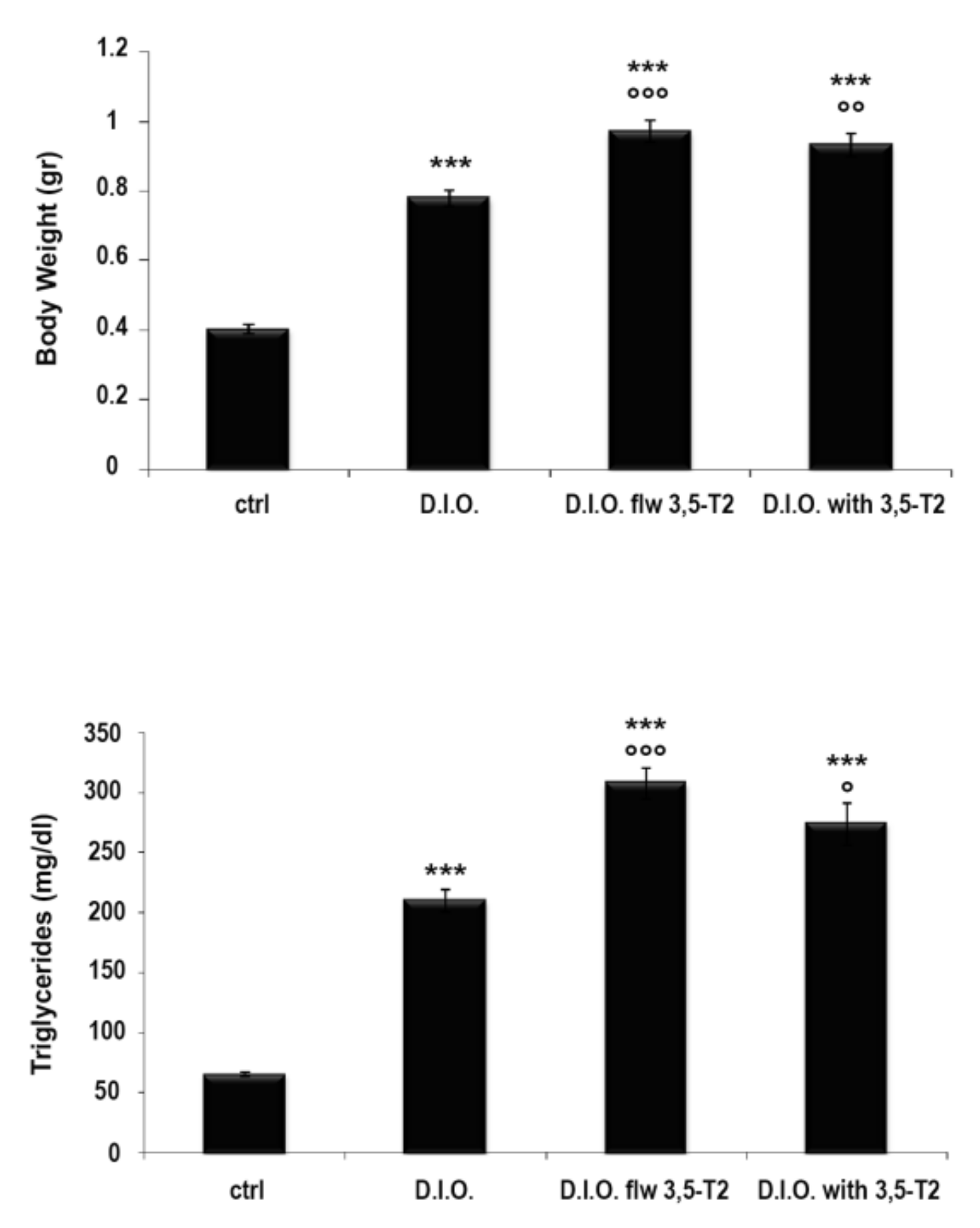
3.1. 3,5-T2 Increased Body Weight and Serum Triglyceride Levels in D.I.O. Zebrafish
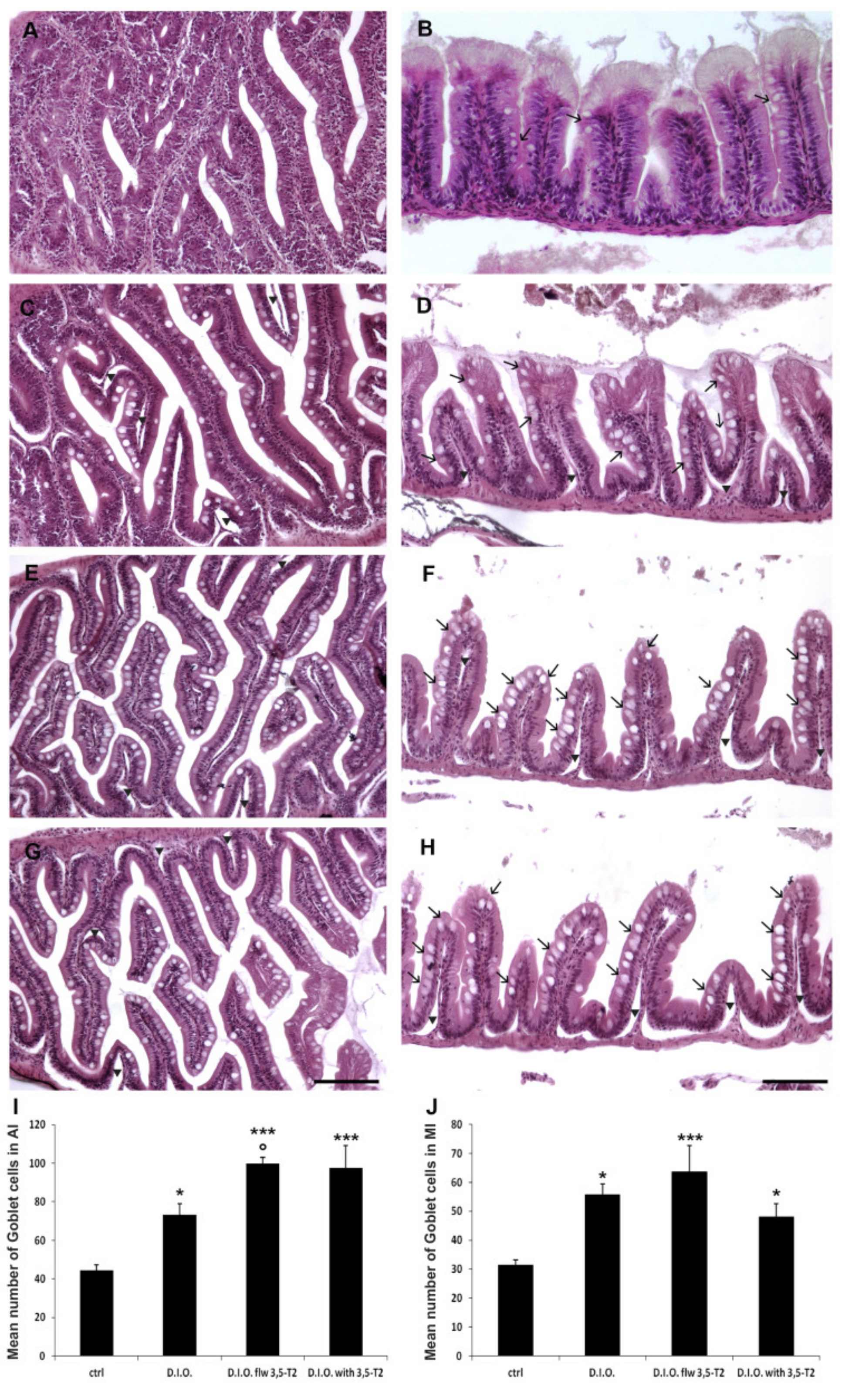
3.2. 3,5-T2 Sustained or Increased Intestinal Morphological Alterations Induced by D.I.O.
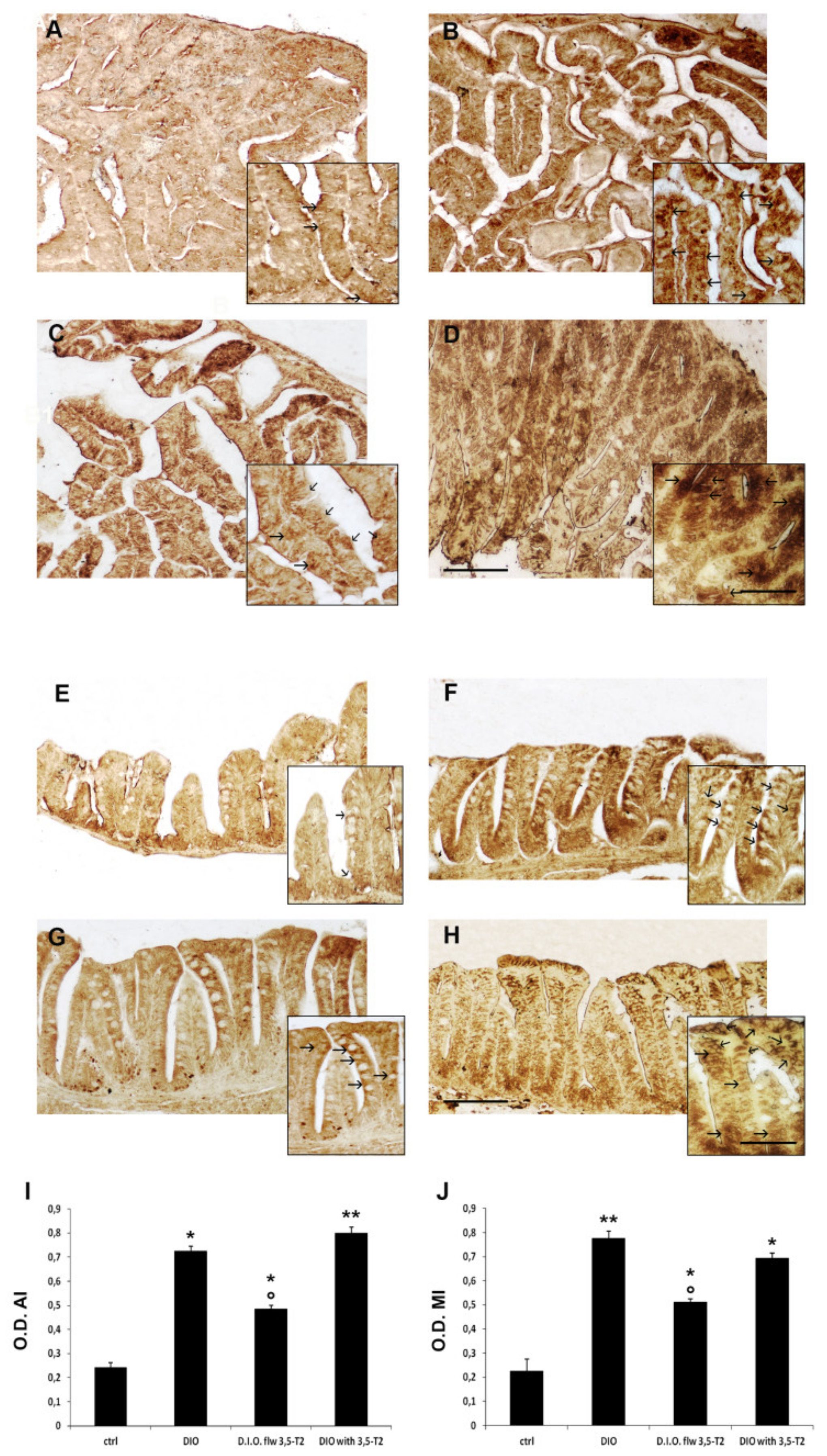
3.3. The Cotreatement with 3,5-T2 Sustained Intestinal Inflammation Induced by D.I.O.
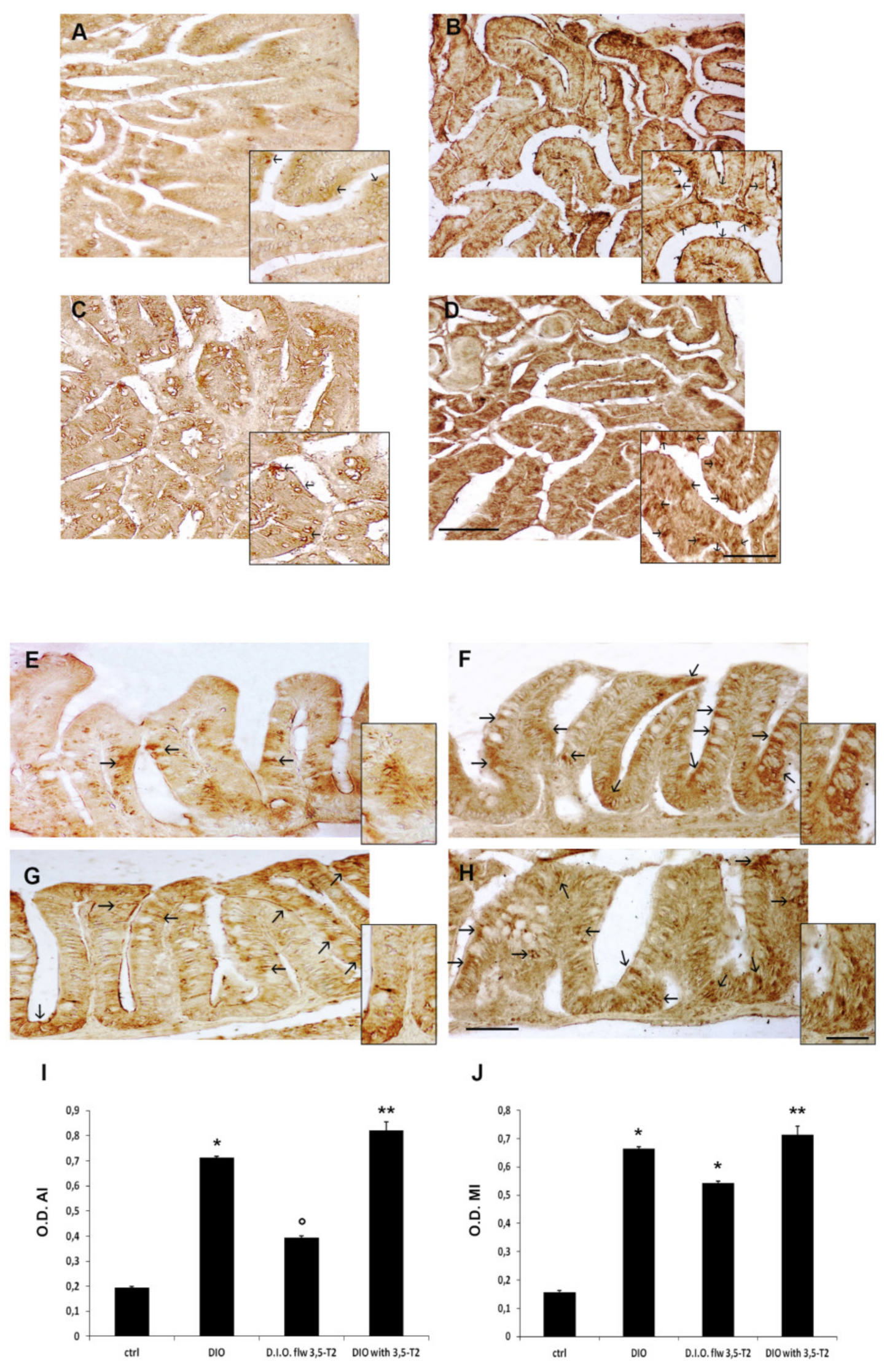
3.4. The 3,5-T2 Sustained Inflammation Induced by D.I.O. Was Accompanied by ER-Stress in the Anterior and Mid Intestine
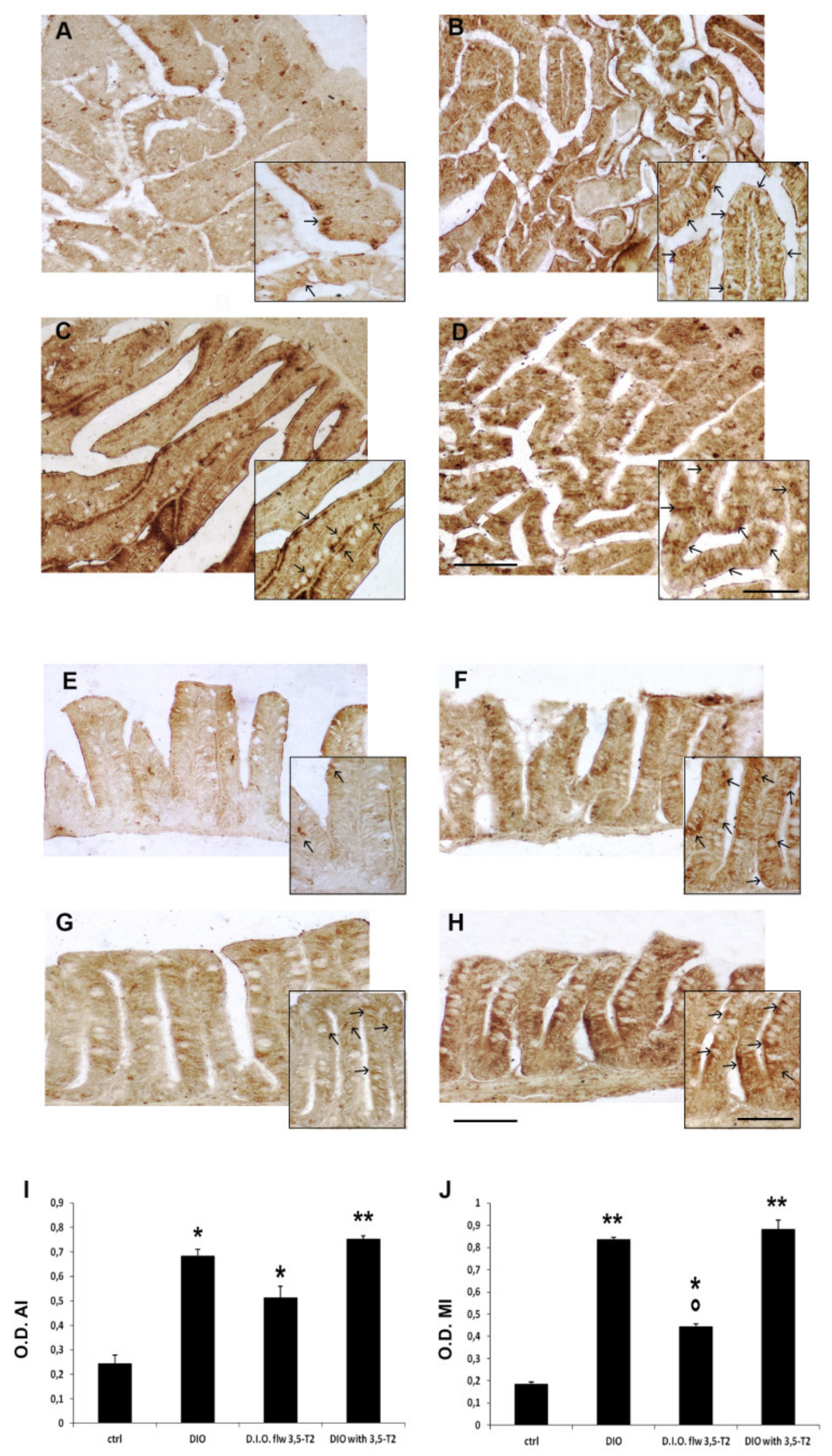
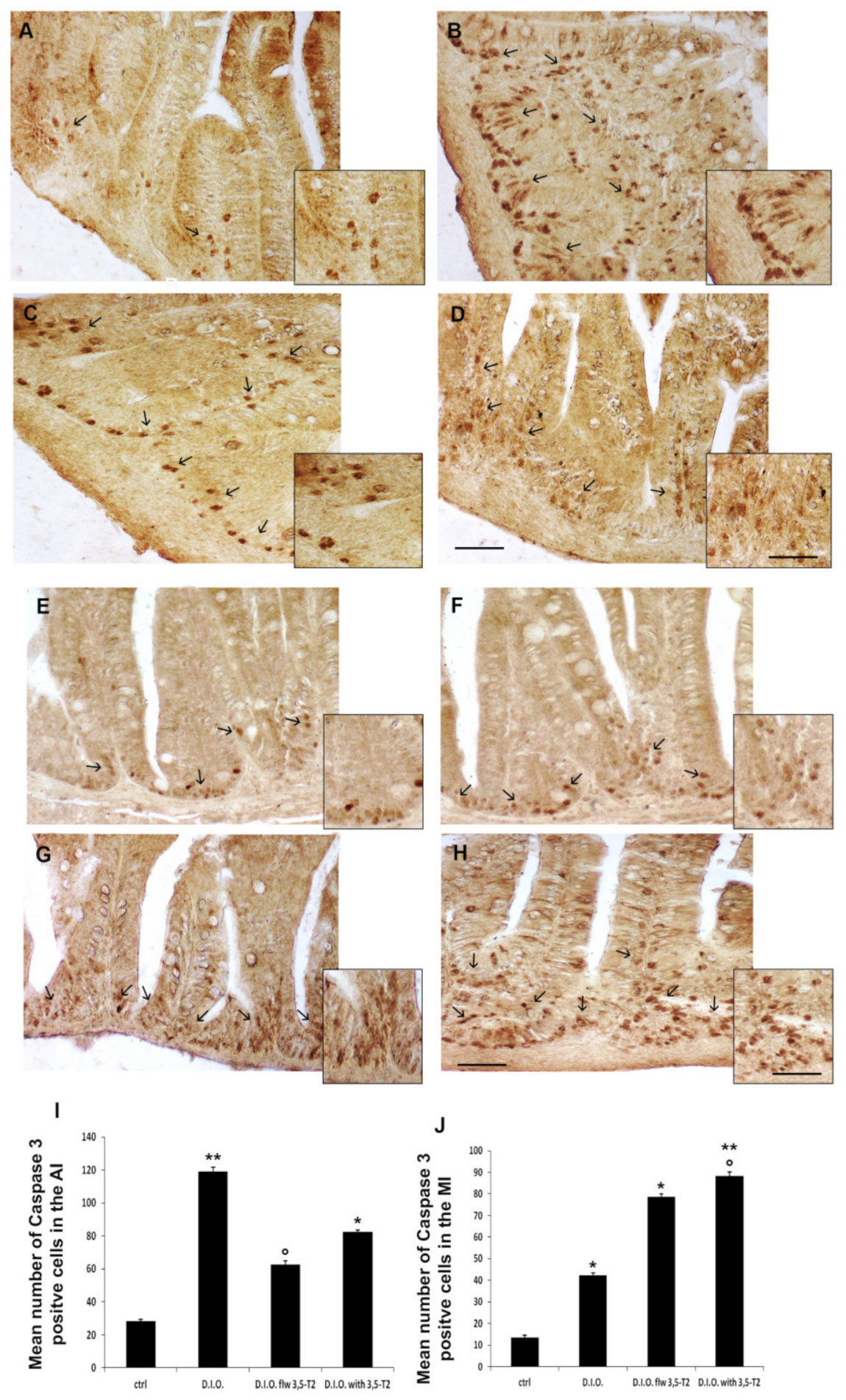
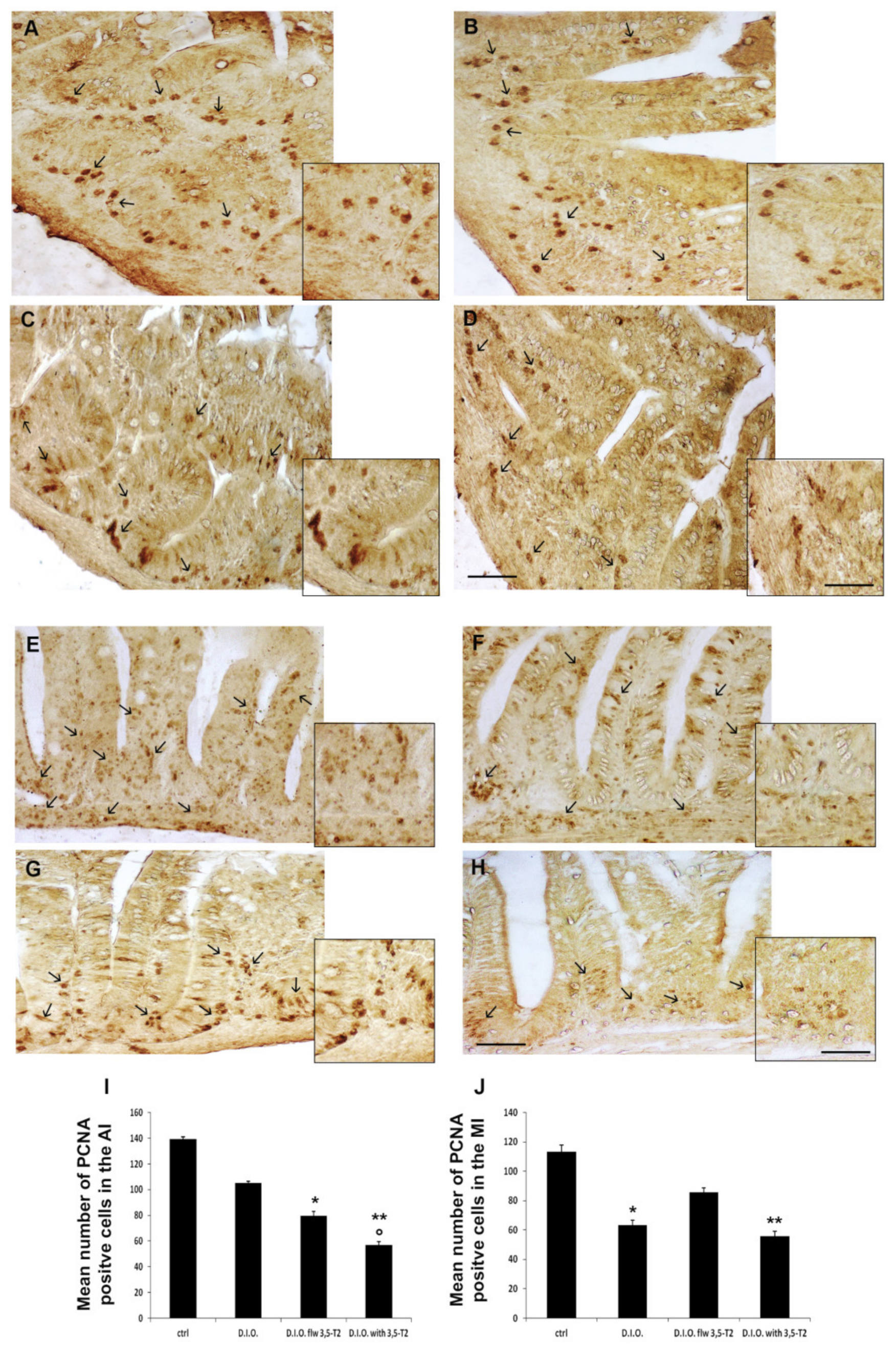
3.5. The 3,5-T2 Sustained Inflammation Induced by D.I.O. Was Accompanied by Alteration of the Cell Turnover in the Anterior and Mid Intestine
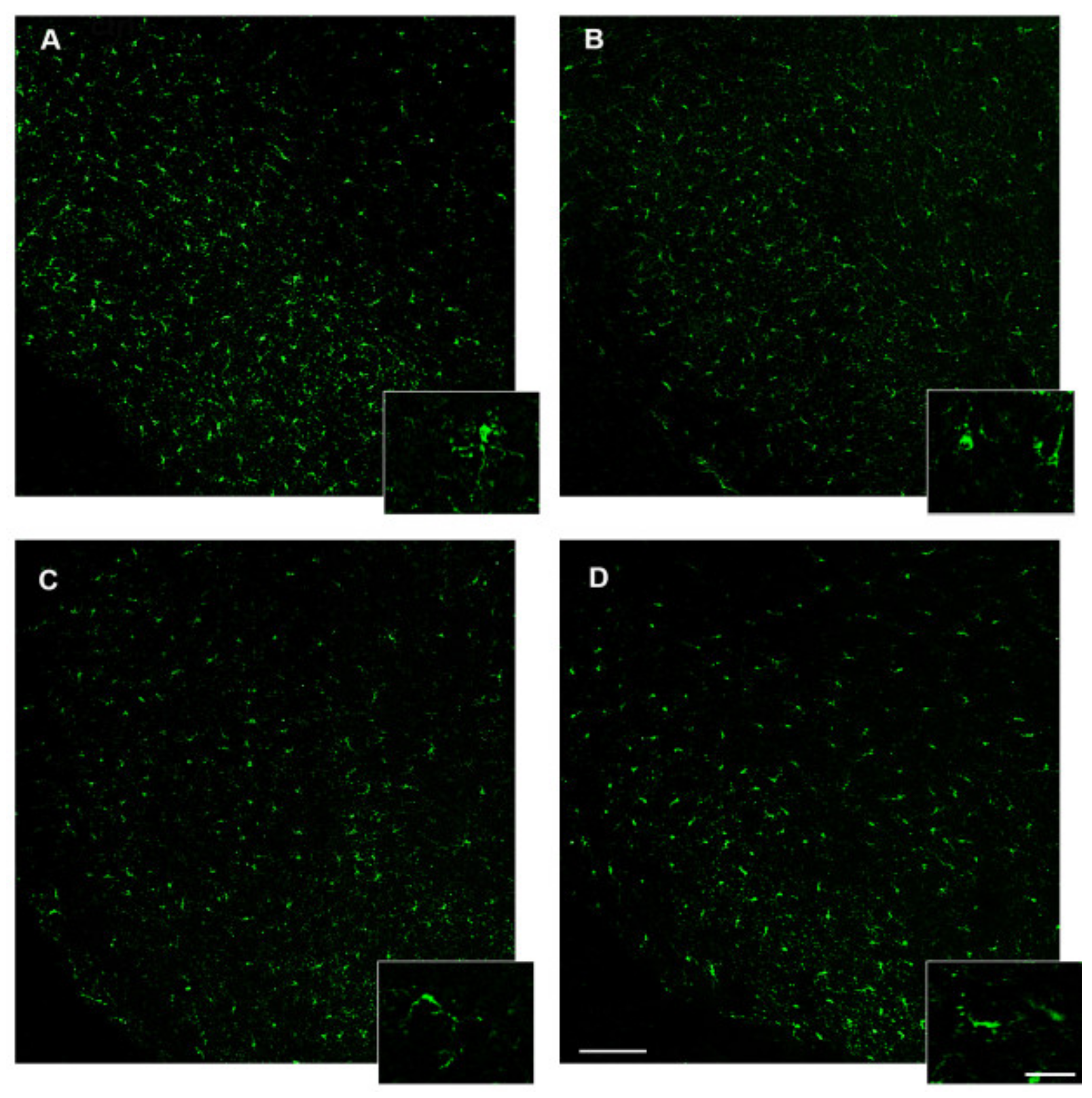
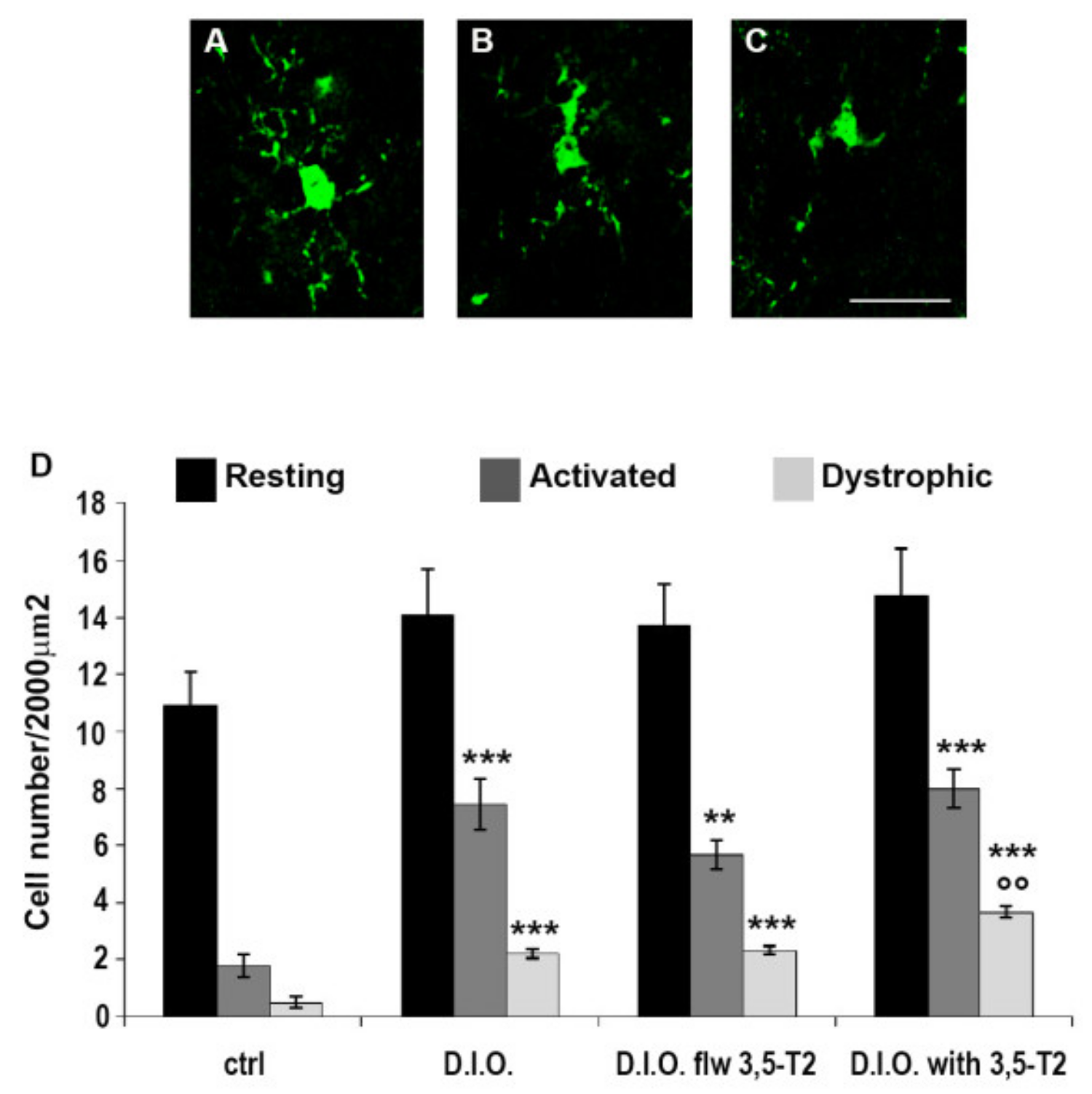
3.6. 3,5-T2 Sustained or Increased Brain Inflammation Induced by D.I.O.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Araujo, J.R.; Tomas, J.; Brenner, C.; Sansonetti, P.J. Impact of high-fat diet on the intestinal microbiota and small intestinal physiology before and after the onset of obesity. Biochimie 2017, 141, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory mechanisms linking obesity and metabolic disease. J. Clin. Invest. 2017, 127, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Arias-Jayo, N.; Abecia, L.; Alonso-Sáez, L.; Ramirez-Garcia, A.; Rodriguez, A.; Pardo, M.A. High-Fat diet consumption induces microbiota dysbiosis and intestinal inflammation in zebrafish. Microb. Ecol. 2018, 76, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Nishimura, Y.; Zang, L.; Hirano, M.; Shimada, Y.; Wang, Z.; Umemoto, N.; Kuroyanagi, J.; Nishimura, N.; Tanaka, T. Diet-induced obesity in zebrafish shares common pathophysiological pathways with mammalian obesity. BMC Physiol. 2010, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Kieffer, T.J.; Habener, J.F. The adipoinsular axis: Effects of leptin on pancreatic β-cells. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E1–E14. [Google Scholar] [CrossRef]
- Bessesen, D.H. Update on obesity. J. Clin. Endocrinol. Metab. 2008, 93, 2027–2034. [Google Scholar] [CrossRef] [Green Version]
- Casper, R.C.; Sullivan, E.L.; Tecott, L. Relevance of animal models to human eating disorders and obesity. Psychopharmacology 2008, 199, 313–329. [Google Scholar] [CrossRef] [Green Version]
- Carnovali, M.; Luzi, L.; Terruzzi, I.; Banfi, G.; Mariotti, M. Metabolic and bone effects of high-fat diet in adult zebrafish. Endocrine 2018, 61, 317–326. [Google Scholar] [CrossRef]
- Wannamethee, S.G.; Tchernova, J.; Whincup, P.; Lowe, G.D.; Kelly, A.; Rumley, A.; Wallace, A.M.; Sattar, N. Plasma leptin: Associations with metabolic, inflammatory and haemostatic risk factors for cardiovascular disease. Atherosclerosis 2007, 19, 418–426. [Google Scholar] [CrossRef]
- Rainone, V.; Schneider, L.; Saulle, I.; Ricci, C.; Biasin, M.; Al-Daghri, N.M.; Giani, E.; Zuccotti, G.V.; Clerici, M.; Trabattoni, D. Upregulation of inflammasome activity and increased gut permeability are associated with obesity in children and adolescents. Int. J. Obes. 2016, 40, 1026–1033. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Bournat, J.C.; Brown, C.W. Mitochondrial dysfunction in obesity. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 446–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cioffi, F.; Senese, R.; Lanni, A.; Goglia, F. Thyroid hormones and mitochondria: With a brief look at derivatives and analogues. Mol. Cell. Endocrinol. 2013, 379, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Goglia, F. The effects of 3,5-diiodothyronine on energy balance. Front. Physiol. 2014, 5, 528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanni, A.; Moreno, M.; Goglia, F. Mitochondrial actions of thyroid hormone. Compr. Physiol. 2016, 6, 1591–1607. [Google Scholar] [CrossRef]
- Senese, R.; de Lange, P.; Petito, G.; Moreno, M.; Goglia, F.; Lanni, A. 3,5-Diiodothyronine: A Novel Thyroid Hormone Metabolite and Potent Modulator of Energy Metabolism. Front. Endocrinol. 2018, 9, 427. [Google Scholar] [CrossRef]
- Lanni, A.; Moreno, M.; Lombardi, A.; Goglia, F. Calorigenic effect of diiodothyronines in the rat. J. Physiol. 1996, 494 (Pt 3), 831–837. [Google Scholar] [CrossRef]
- Silvestri, E.; Cioffi, F.; De Matteis, R.; Senese, R.; de Lange, P.; Coppola, M.; Salzano, A.A.; Scaloni, A.; Ceccarelli, M.; Goglia, F.; et al. 3,5-Diiodo-L-thyronine affects structural and metabolic features of skeletal muscle mitochondria in high-fat-diet fed rats producing a co-adaptation to the glycolytic fiber phenotype. Front. Physiol. 2018, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Coppola, M.; Cioffi, F.; Moreno, M.; Goglia, F.; Silvestri, E. 3,5-diiodo-L-thyronine: A Possible Pharmacological Agent? Curr. Drug. Deliv. 2016, 13, 330–338. [Google Scholar] [CrossRef]
- Jonas, W.; Lietzow, J.; Wohlgemuth, F.; Hoefig, C.S.; Wiedmer, P.; Schweizer, U.; Köhrle, J.; Schürmann, A. 3,5-Diiodo-L-Thyronine (3,5-T2) Exerts Thyromimetic Effects on Hypothalamus-Pituitary-Thyroid Axis, Body Composition, and Energy Metabolism in Male Diet-Induced Obese Mice. Endocrinology 2015, 156, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Teixeira, S.; Filgueira, C.; Sieglaff, D.H.; Benod, C.; Villagomez, R.; Minze, L.J.; Zhang, A.; Webb, P.; Nunes, M.T. 3,5-diiodothyronine (3,5-T2) reduces blood glucose independently of insulin sensitization in obese mice. Acta Physiol. 2017, 220, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Glinni, D.; Moreno, M.; Cioffi, F.; Silvestri, E.; Goglia, F. Thyroid hormone analogues and derivatives: Actions in fatty liver. World J. Hepatol. 2014, 6, 114–129. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, L.; Lossi, L.; Merighi, A.; de Girolamo, P. Anatomical features for the adequate choice of experimental animal models in biomedicine: I. Fishes. Ann. Anat. 2016, 205, 75–84. [Google Scholar] [CrossRef]
- Pappalardo, A.; Porreca, I.; Caputi, L.; De Felice, E.; Schulte-Merker, S.; Zannini, M.; Sordino, P. Thyroid development in zebrafish lacking Taz. Mech. Dev. 2015, 3, 268–278. [Google Scholar] [CrossRef]
- Porreca, I.; De Felice, E.; Fagman, H.; Di Lauro, R.; Sordino, P. Zebrafish bcl2l is a survival factor in thyroid development. Dev. Biol. 2012, 15, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Olvera, A.; Martyniuk, C.J.; Buisine, N.; Jimenez-Jacinto, V.; Sanchez-Flores, A.; Sachs, L.M.; Orozco, A. Differential transcriptome regulation by 3,5-T2 and 3′,3,5-T3 in brain and liver uncovers novel roles for thyroid hormones in tilapia. Sci. Rep. 2017, 7, 15043. [Google Scholar] [CrossRef] [Green Version]
- Little, A.G.; Seebacher, F. Thyroid hormone regulates cardiac performance during cold acclimation in zebrafish (Danio rerio). J. Exp. Biol. 2014, 217 (Pt 5), 718–725. [Google Scholar] [CrossRef] [Green Version]
- Navarrete-Ramirez, P.; Luna, M.; Valverde, R.C.; Orozco, A. 3,5-di-iodothyronine stimulates tilapia growth through an alternate isoform of thyroid hormone receptor beta1. J. Mol. Endocrinol. 2014, 52, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Imperatore, R.; D’Angelo, L.; Safari, O.; Motlagh, H.A.; Piscitelli, F.; de Girolamo, P.; Cristino, L.; Varricchio, E.; Di Marzo, V.; Paolucci, M. Overlapping Distribution of Orexin and Endocannabinoid Receptors and Their Functional Interaction in the Brain of Adult Zebrafish. Front. Neuroanat. 2018, 30, 12–62. [Google Scholar] [CrossRef] [Green Version]
- García, G.C.; Jeziorski, M.C.; Valverde, R.C.; Orozco, A. Effects of iodothyronines on the hepatic outer-ring deiodinating pathway in killifish. Gen. Comp. Endocrinol. 2004, 135, 201–209. [Google Scholar] [CrossRef]
- García, G.C.; López-Bojorquez, L.; Nuñez, J.; Valverde, R.C.; Orozco, A. 3,5-Diiodothyronine in vivo maintains euthyroidal expression of type 2 iodothyronine deiodinase, growth hormone, and thyroid hormone receptor beta1 in the killifish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R877–R883. [Google Scholar] [CrossRef] [PubMed]
- Mania, M.; Maruccio, L.; Russo, F.; Abbate, F.; Castaldo, L.; D’Angelo, L.; de Girolamo, P.; Guerrera, M.C.; Lucini, C.; Madrigrano, L.; et al. Expression and distribution of leptin and its receptors in the digestive tract of DIO (diet-induced obese) zebrafish. Ann. Anat. 2017, 212, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Imperatore, R.; D’Angelo, L.; De Girolamo, P.; Cristino, L.; Paolucci, M. Identification of Orexin and Endocannabinoid Receptors in Adult Zebrafish Using Immunoperoxidase and Immunofluorescence Methods. J. Vis. Exp. 2019, 148. [Google Scholar] [CrossRef] [PubMed]
- Varricchio, E.; Russo, F.; Coccia, E.; Turchini, G.; Francis, D.; De Girolamo, P.; Paolucci, M. Immunohistochemical and immunological detection of ghrelin and leptin in rainbow trout oncorhynchus mykiss and murray cod maccullochella peelii peelii as affected by different dietary fatty acids. Microsc. Res. Tech. 2012, 75, 771–780. [Google Scholar] [CrossRef]
- Streit, W.J.; Xue, Q.S. Life and death of microglia. J. Neuroimmun. Pharmacol. 2009, 4, 371–379. [Google Scholar] [CrossRef]
- Antonelli, A.; Fallahi, P.; Ferrari, S.M.; Di Domenicantonio, A.; Moreno, M.; Lanni, A.; Goglia, F. 3,5-diiodo-L-thyronine increases resting metabolic rate and reduces body weight without undesirable side effects. J. Biol. Regul. Homeost. Agents 2011, 25, 655–660. [Google Scholar]
- Moreno, M.; de Lange, P.; Lombardi, A.; Silvestri, E.; Lanni, A.; Goglia, F. Metabolic effects of thyroid hormone derivatives. Thyroid 2008, 18, 239–253. [Google Scholar] [CrossRef]
- Cioffi, F.; Lanni, A.; Goglia, F. Thyroid hormones, mitochondrial bioenergetics and lipid handling. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 402–407. [Google Scholar] [CrossRef]
- Hernandez, A. 3,5-Diiodo-L-Thyronine (T2) in Dietary Supplements: What Are the Physiological Effects? Endocrinology 2015, 156, 5–7. [Google Scholar] [CrossRef]
- Brugman, S. The zebrafish as a model to study intestinal inflammation. Dev. Comp. Immunol. 2016, 64, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Du, J.; Lam, S.H.; Mathavan, S.; Matsudaira, P.; Gong, Z. Morphological and molecular evidence for functional organization along the rostrocaudal axis of the adult zebrafish intestine. BMC Genomics 2010, 11, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldsmith, J.R.; Jobin, C. Think small: Zebrafish as a model system of human pathology. J. Biomed. Biotechnol. 2012, 2012, 817341. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Ryan, P.M.; Cryan, J.F.; Dinan, T.G.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Gut microbiota, obesity and diabetes. Postgrad. Med. J. 2016, 92, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Vatner, D.F.; Snikeris, J.; Popov, V.; Perry, R.J.; Rahimi, Y.; Samuel, V.T. 3 3,5 Diiodo-L-Thyronine (T2) does not prevent hepatic steatosis or insulin resistance in fat-fed sprague dawley rats. PLoS ONE 2015, 20, e0140837. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57(6), 1470–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landgraf, K.; Schuster, S.; Meusel, A.; Garten, A.; Riemer, T.; Schleinitz, D.; Kiess, W.; Körner, A. Short-term overfeeding of zebrafish with normal or high-fat diet as a model for the development of metabolically healthy versus unhealthy obesity. BMC Physiol. 2017, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.; Stephens, W.Z.; Burns, A.R.; Stagaman, K.; David, L.A.; Bohannan, B.J.; Guillemin, K.; Rawls, J.F. Ontogenetic differences in dietary fat influence microbiota assembly in the zebrafish gut. MBio 2015, 6, e00687-15. [Google Scholar] [CrossRef] [Green Version]
- Zang, L.; Maddison, L.A.; Chen, W. Zebrafish as a Model for Obesity and Diabetes. Front. Cell Dev. Biol. 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Cole, M.A.; Murray, A.J.; Cochlin, L.E.; Heather, L.C.; McAleese, S.; Knight, N.S.; Sutton, E.; Jamil, A.A.; Parassol, N.; Clarke, K. A high fat diet increases mitochondrial fatty acid oxidation and uncoupling to decrease efficiency in rat heart. Basic Res. Cardiol. 2011, 106, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Ørgensen, W.; Rud, K.A.; Mortensen, O.H.; Frandsen, L.; Grunnet, N.; Quistorff, B. Your mitochondria are what you eat: A high-fat or a high-sucrose diet eliminates metabolic flexibility in isolated mitochondria from rat skeletal muscle. Physiol. Rep. 2017, 5, e13207. [Google Scholar] [CrossRef]
- De Lange, P.; Cioffi, F.; Senese, R.; Moreno, M.; Lombardi, A.; Silvestri, E.; De Matteis, R.; Lionetti, L.; Mollica, M.P.; Goglia, F.; et al. Nonthyrotoxic prevention of diet-induced insulin resistance by 3,5-diiodo-L-thyronine in rats. Diabetes 2011, 60, 2730–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grasselli, E.; Voci, A.; Demori, I.; Canesi, L.; Goglia, F.; Lanni, A.; Gallo, G.; Vergani, L. 3,5-Diiodo-L-thyronine modulates the expression of genes of lipid metabolism in a rat model of fatty liver. J. Endocrinol. 2012, 212, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, A.; Senese, R.; Busiello, R.A.; Cioffi, F.; Goglia, F.; Lanni, A. 3,5-diiodo-L-thyronine activates brown adipose tissue thermogenesis in hypothyroid rats. PLoS ONE 2015, 10(2), e0116498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavallo, A.; Priore, P.; Gnoni, G.V.; Papa, S.; Zanotti, F.; Gnoni, A. 3,5-Diiodo-L-thyronine administration to hypothyroid rats rapidly enhances fatty acid oxidation rate and bioenergetic parameters in liver cells. PLoS ONE 2013, 8, e52328. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, I.J.; Huang, L.S.; Huggins, L.A.; Yu, S.; Nagareddy, P.R.; Scanlan, T.S.; Ehrenkranz, J.R. Thyroid hormone reduces cholesterol via a non-LDL receptor-mediated pathway. Endocrinology 2012, 153, 5143–5149. [Google Scholar] [CrossRef] [Green Version]
- Al-Asmakh, M.; Anuar, F.; Zadjali, F.; Rafter, J.; Pettersson, S. Gut microbial communities modulating brain development and function. Gut Microbes 2012, 3, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Goglia, F. Biological effects of 3,5-diiodothyronine (T(2)). Biochemistry 2005, 70, 164–172. [Google Scholar] [CrossRef]
- Koerner, D.; Schwartz, H.L.; Surks, M.I.; Oppenheimer, J.H. Binding of selected iodothyronine analogues to receptor sites of isolated rat hepatic nuclei. High correlation between structural requirements for nuclear binding and biological activity. J. Biol. Chem. 1975, 250, 6417–6423. [Google Scholar]
- Lietzow, J.; Golchert, J.; Homuth, G.; Völker, U.; Jonas, W.; Köhrle, J. 3,5-T2 alters murine genes relevant for xenobiotic, steroid, and thyroid hormone metabolism. J. Mol. Endocrinol. 2016, 56, 311–323. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imperatore, R.; Tunisi, L.; Mavaro, I.; D’Angelo, L.; Attanasio, C.; Safari, O.; Motlagh, H.A.; De Girolamo, P.; Cristino, L.; Varricchio, E.; et al. Immunohistochemical Analysis of Intestinal and Central Nervous System Morphology in an Obese Animal Model (Danio rerio) Treated with 3,5-T2: A Possible Farm Management Practice? Animals 2020, 10, 1131. https://doi.org/10.3390/ani10071131
Imperatore R, Tunisi L, Mavaro I, D’Angelo L, Attanasio C, Safari O, Motlagh HA, De Girolamo P, Cristino L, Varricchio E, et al. Immunohistochemical Analysis of Intestinal and Central Nervous System Morphology in an Obese Animal Model (Danio rerio) Treated with 3,5-T2: A Possible Farm Management Practice? Animals. 2020; 10(7):1131. https://doi.org/10.3390/ani10071131
Chicago/Turabian StyleImperatore, Roberta, Lea Tunisi, Isabella Mavaro, Livia D’Angelo, Chiara Attanasio, Omid Safari, Hamidreza Ahmadniaye Motlagh, Paolo De Girolamo, Luigia Cristino, Ettore Varricchio, and et al. 2020. "Immunohistochemical Analysis of Intestinal and Central Nervous System Morphology in an Obese Animal Model (Danio rerio) Treated with 3,5-T2: A Possible Farm Management Practice?" Animals 10, no. 7: 1131. https://doi.org/10.3390/ani10071131
APA StyleImperatore, R., Tunisi, L., Mavaro, I., D’Angelo, L., Attanasio, C., Safari, O., Motlagh, H. A., De Girolamo, P., Cristino, L., Varricchio, E., & Paolucci, M. (2020). Immunohistochemical Analysis of Intestinal and Central Nervous System Morphology in an Obese Animal Model (Danio rerio) Treated with 3,5-T2: A Possible Farm Management Practice? Animals, 10(7), 1131. https://doi.org/10.3390/ani10071131