Genetic Characterization of the “Chusca Lojana”, a Creole Goat Reared in Ecuador, and Its Relationship with Other Goat Breeds

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and DNA Extraction
2.2. Genotyping Using STR Markers
2.3. Genetic Diversity Within-Breed
2.4. Genetic Diversity Inter-Breed
2.5. Genetic Structure of Chusca Lojana Goat
3. Results
3.1. Genetic Diversity Within-Breed
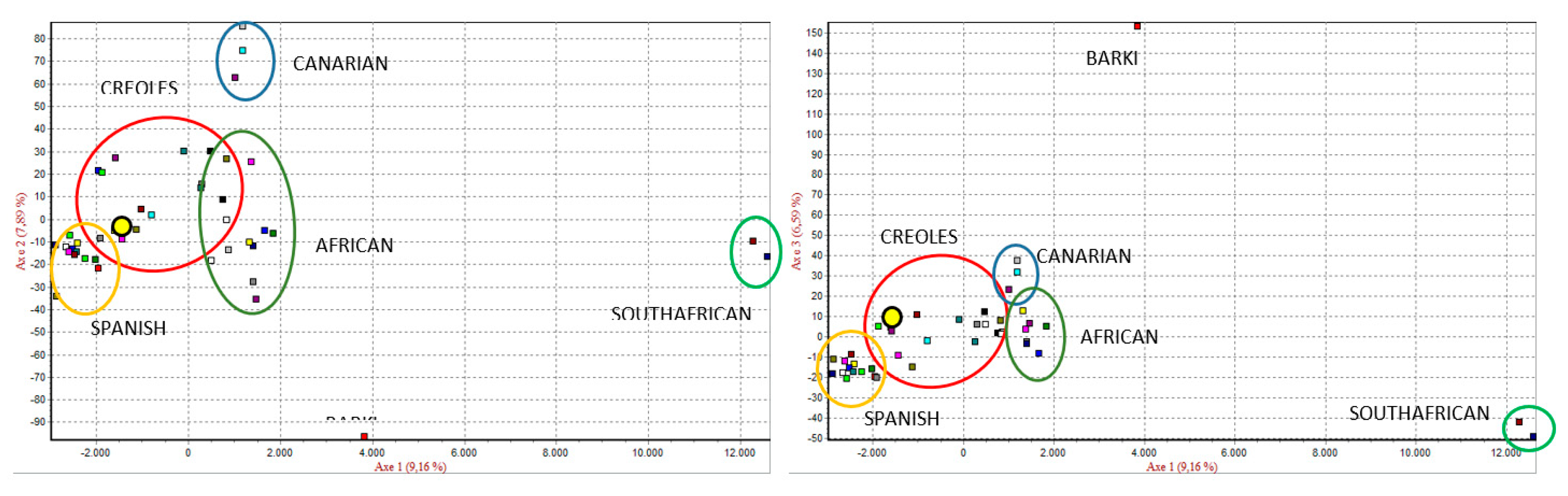
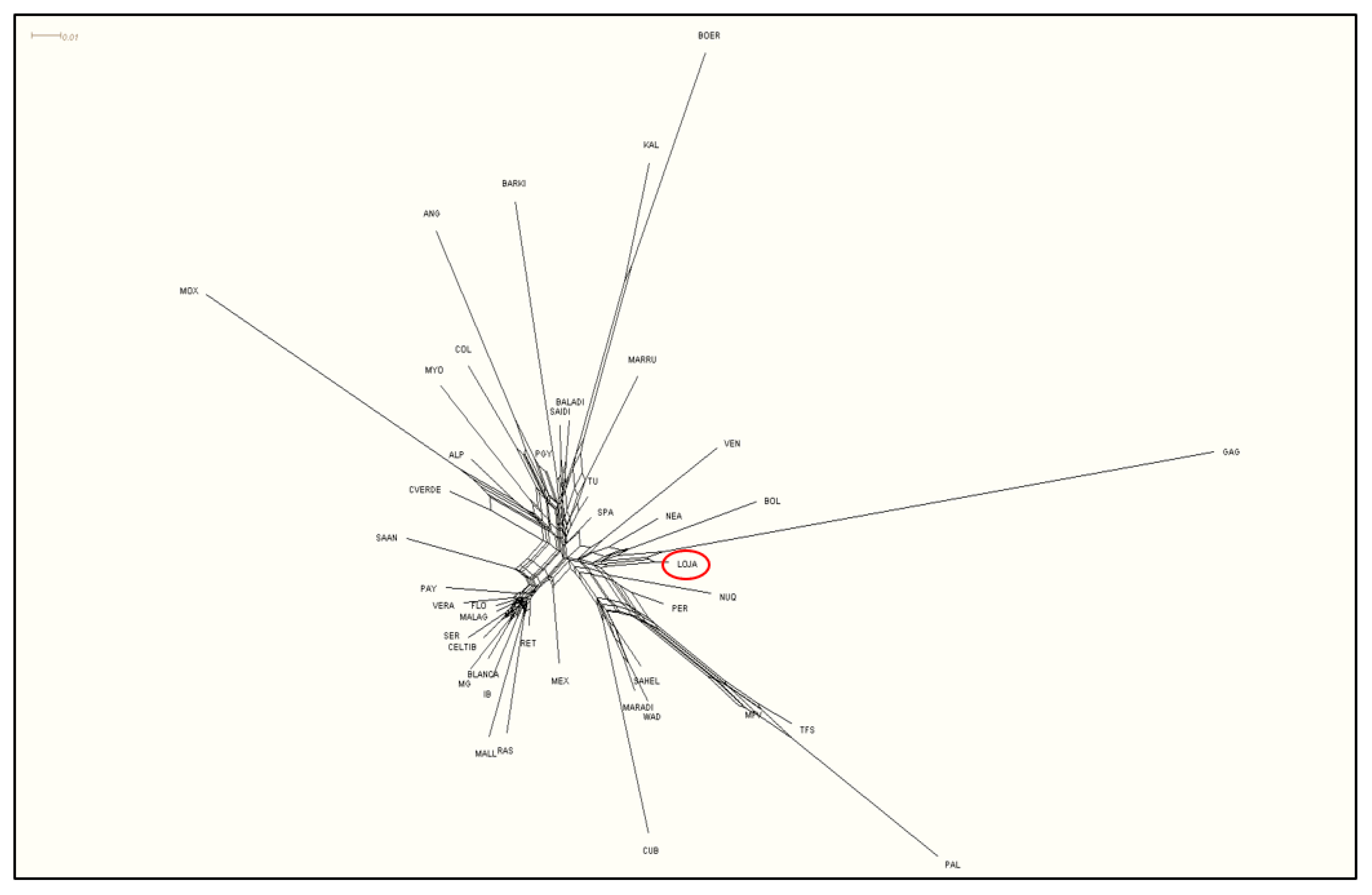
3.2. Genetic Structure of the Chusca Lojana Goat
3.3. Genetic Diversity Inter-Breed
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Notes
Funding
Acknowledgments
Conflicts of Interest
References
- Mittermeier, R.A.; Myers, N.; Gil, P.R.; Mittermeier, C.G. Hotspots: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; Agrupación Sierra Madre: Mexico City, Mexico, 1999; Volume 3, pp. 39–40. [Google Scholar]
- Ladle, R.J.; Jepson, P.; Araújo, M.B.; Whittaker, R.J. Dangers of crying wolf over risk of extinctions. Nature 2004, 428, 799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tollefson, J. One Million Species Face Extinction. Nature 2019, 569, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations. The State of the World’s Biodiversity for Food and Agriculture; Bélanger, J., Pilling, D., Eds.; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2019; p. 572. [Google Scholar]
- Hanotte, O.; Dessie, T.; Kemp, S. Time to Tap Africa’s Livestock Genomes. Science 2010, 328, 1640–1641. [Google Scholar] [CrossRef]
- Ginja, C.; Cortes, O.; Gama, L.T.; Delgado, J.V.; Amills, M.; De Sousa, C.B.; Canon, J.; Capote, J.; Dunner, S.; Ferrando, A.; et al. Conservation of Goat Populations from Southwestern Europe Based on Molecular Diversity Criteria. In Sustainable Goat Production in Adverse Environments; Simões, J., Gutierrez, C., Eds.; Springer International Publishing: Basel, Switzerland, 2017; Volume 1, pp. 509–533. [Google Scholar]
- Taberlet, P.; Valentini, A.; Rezaei, H.R.; Naderi, S.; Pompanon, F.; Negrini, R.; Ajmone-Marsan, P. Are cattle, sheep, and goats endangered species? Mol. Ecol. 2008, 17, 275–284. [Google Scholar] [CrossRef]
- Ajmone-Marsan, P.; Colli, L.; Han, J.L.; Achilli, A.; Lancioni, H.; Joost, S.; Crepaldi, P.; Pilla, F.; Stella, A.; Taberlet, P.; et al. The characterization of goat genetic diversity: Towards a genomic approach. Small Rumin. Res. 2014, 121, 58–72. [Google Scholar] [CrossRef]
- Dubeuf, J.-P.; Morand-Fehr, P.; Rubino, R. Situation, changes and future of goat industry around the world. Small Rumin. Res. 2004, 51, 165–173. [Google Scholar] [CrossRef]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Chelex 100 as a Medium for Simple Extraction of DNA for PCR-Based Typing from Forensic Material. Biotechniques 2013, 54, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Park, S.D.E. Trypanotolerance in West African Cattle and the Population Genetic Effects of Selection. Ph.D. Thesis, University of Dublin, Dublin, Ireland, September 2001. [Google Scholar]
- Yeh, F.; Boylet, J. Population Genetic Analysis of Codominant and Dominant Markers and Quantitative Traits. Belg. J. Bot. 1997, 129, 157–163. [Google Scholar]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX 4.05, logiciel sous Windows TM pour la génétique des populations. Laboratoire Génome, Populations, Interactions 2004. (In French) [Google Scholar]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Guo, S.W.; Thompson, E.A. Performing the Exact Test of Hardy-Weinberg Proportion for Multiple Alleles. Biometrics 1992, 48, 361. [Google Scholar] [CrossRef]
- Wright, S. The Theory of Gene Frequencies. In Evolution and the Genetics of Populations; University of Chicago Press: Chicago, MI, USA, 1969; Volume 2, pp. 291–293. [Google Scholar]
- Reynolds, J.; Weir, B.S.; Cockerham, C.C. Estimation of the Coancestry Coefficient: Basis for a Short-Term Genetic Distance. Genetics 1983, 105, 767–779. [Google Scholar]
- Langella, O. Populations 1.2.28. 1999. Available online: http://bioinformatics.org/populations/index.html (accessed on 26 September 2018).
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2005, 23, 254–267. [Google Scholar] [CrossRef]
- Bowcock, A.M.; Ruiz-Linares, A.; Tomfohrde, J.; Minch, E.; Kidd, J.R.; Cavalli-Sforza, L.L. High resolution of human evolutionary trees with polymorphic microsatellites. Nature 1994, 368, 455–457. [Google Scholar] [CrossRef]
- Page, R.D.M. Tree View: An Application to Display Phylogenetic Trees on Personal Computers. Bioinformatics 1996, 12, 357–358. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar]
- Alderson, G.L.H. Conservation of breeds and maintenance of biodiversity: Justification and methodology for the conservation of Animal Genetic Resources. Arch. Zootec. 2018, 67, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Ginja, C.; Gama, L.T.; Martínez, A.; Sevane, N.; Martin-Burriel, I.; Lanari, M.R.; Revidatti, M.A.; Aranguren-Méndez, J.A.; Bedotti, D.O.; Ribeiro, M.N.; et al. Genetic diversity and patterns of population structure in Creole goats from the Americas. Anim. Genet. 2017, 48, 315–329. [Google Scholar] [CrossRef]
- Canon, J.; García, D.; García-Atance, M.A.; Obexer-Ruff, G.; Lenstra, J.A.; Ajmone-Marsan, P.; Dunner, S.; The ECOGENE Consortium. Geographical partitioning of goat diversity in Europe and the Middle East. Anim. Genet. 2006, 37, 327–334. [Google Scholar] [CrossRef]
- Martínez, A.M.; Gama, L.T.; Bermejo, J.V.D.; Cañón, J.; Amills, M.; De Sousa, C.B.; Ginja, C.; Zaragoza, P.; Manunza, A.; Landi, V.; et al. The Southwestern fringe of Europe as an important reservoir of caprine biodiversity. Genet. Sel. Evol. 2015, 47, 86. [Google Scholar] [CrossRef] [Green Version]
- Sevane, N.; Cortés, O.; Gama, L.T.; Martínez, A.; Zaragoza, P.; Amills, M.; Bedotti, D.O.; De Sousa, C.B.; Cañon, J.; Dunner, S.; et al. Dissection of ancestral genetic contributions to Creole goat populations. Animal 2018, 12, 2017–2026. [Google Scholar] [CrossRef] [Green Version]
- Periasamy, K.; Vahidi, S.M.F.; Silva, P.; Faruque, M.O.; Naqvi, A.N.; Basar, M.; Cao, J.; Zhao, S.; Thuy, L.T.; Pichler, R.; et al. Mapping molecular diversity of indigenous goat genetic resources of Asia. Small Rumin. Res. 2017, 148, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Chacón, Y.E.; La, O.M.; Velásquez, F.J.; Pérez, J.E.; Delgado, V.; Cos, Y.; Martínez, A. Validación de los estándares raciales de la cabra criolla cubana para su registro internacional (Validation of the Racial Standards of the Cuban Creole Goat for Its International Registration). REDVET Revista Electrónica de Veterinaria 2012, 13, 1–8. (In Spanish) [Google Scholar]
- Bulut, Z.; Kurar, E.; Ozsensoy, Y.; Altunok, V.; Nizamlioglu, M. Genetic Diversity of Eight Domestic Goat Populations Raised in Turkey. BioMed Res. Int. 2016, 2016, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Elbeltagy, A.R.; Naga, A.M.A.; Hassen, H.; Solouma, G.M.; Rischkowsky, B.; Mwacharo, J.M. Genetic diversity and structure of goats within an early livestock dispersal area in Eastern North Africa. Afr. J. Biotechnol. 2016, 15, 431–441. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed/Population | Acronym | Origin | n | |
|---|---|---|---|---|
| 1 | Chusca Lojana | LOJ | Ecuador | 145 |
| 2 | Galapagos goat | GAG | Galapagos Islands (Ecuador) | 24 |
| 3 | Spanish goat | SPA | USA | 64 |
| 4 | Myotonic | MYO | USA | 43 |
| 5 | Mexicana | MEX | Mexico | 70 |
| 6 | Creole from Cuba | CUB | Cuba | 40 |
| 7 | Creole from Colombia | COL | Colombia | 24 |
| 8 | Creole from Venezuela | VEN | Venezuela | 45 |
| 9 | Creole from Perú | PER | Peru | 61 |
| 10 | Moxotó | MOX | Brazil | 40 |
| 11 | Creole from Bolivia | BOL | Bolivia | 40 |
| 12 | Creole from Paraguay | PGY | Paraguay | 84 |
| 13 | North-east creole | NEA | Argentina | 40 |
| 14 | Neuquina | NUQ | Argentina | 51 |
| 15 | Blanca de Rasquera | RAS | Spain | 56 |
| 16 | Retinta | RET | Spain | 15 |
| 17 | Verata | VERA | Spain | 30 |
| 18 | Blanca Andaluza | BLANCA | Spain | 40 |
| 19 | Celtibérica | CELTIB | Spain | 40 |
| 20 | Malagueña | MALAG | Spain | 40 |
| 21 | Murciano-Granadina | MG | Spain | 40 |
| 22 | Florida | FLO | Spain | 50 |
| 23 | Payoya | PAY | Spain | 36 |
| 24 | Negra Serrana | SER | Spain | 42 |
| 25 | Pitiusa | IB | Islas Baleares (Spain) | 79 |
| 26 | Mallorquina | MALL | Islas Baleares (Spain) | 70 |
| 27 | Majorera | MFV | Islas Canarias (Spain) | 64 |
| 28 | Palmera | PAL | Islas Canarias (Spain) | 32 |
| 29 | Tinerfeña | TF2 | Islas Canarias (Spain) | 70 |
| 30 | Cabo Verde | CVERDE | Cabo Verde | 37 |
| 31 | Barki | BARKI | Egypt | 44 |
| 32 | Baladi | BALADI | Egypt | 31 |
| 33 | Saidi | SAIDI | Egypt | 34 |
| 34 | Morocco | MOR | Marruecos | 24 |
| 35 | Tunisian Local Goat | TU | Tunisia | 58 |
| 36 | Maradi | MARADI | Nigeria | 47 |
| 37 | West African Dwarf | WAD | Nigeria | 52 |
| 38 | Sahel | SAHEL | Nigeria | 46 |
| 39 | Kalahari Goat | KAL | South Africa | 47 |
| 40 | Boer | BOER | South Africa | 46 |
| 41 | Saanen | SAAN | International | 36 |
| 42 | Alpina | ALP | International | 37 |
| 43 | Anglo-Nubiana | ANG | International | 41 |
| Microsatellites | MNA | Ae | He | Ho | PIC | FIS | FIS IC | HWEd |
|---|---|---|---|---|---|---|---|---|
| BM1258 | 12 | 5.04 | 0.804 | 0.755 | 0.78 | 0.06116 | (−0.02427–0.13910) | NS |
| BM1329 | 8 | 5.99 | 0.836 | 0.741 | 0.81 | 0.11373 | (−0.02560–0.19802) | NS |
| BM1818 | 9 | 4.65 | 0.788 | 0.746 | 0.76 | 0.05246 | (−0.02859–0.13614) | NS |
| BM6506 | 11 | 6.73 | 0.855 | 0.746 | 0.84 | 0.12696 | (0.04177–0.20820) | NS |
| BM6526 | 9 | 5.65 | 0.826 | 0.819 | 0.8 | 0.008 | (−0.06846–0.07866) | NS |
| BM8125 | 7 | 2.35 | 0.576 | 0.559 | 0.53 | 0.03099 | (−0.08580–0.14336) | NS |
| CSRD247 | 7 | 4.61 | 0.786 | 0.729 | 0.76 | 0.07246 | (−0.01127–0.15292) | NS |
| CSRM60 | 8 | 4.43 | 0.777 | 0.748 | 0.74 | 0.03741 | (−0.05035–0.12679) | NS |
| CSSM66 | 20 | 7.02 | 0.861 | 0.69 | 0.84 | 0.1987 | (0.11256–0.27766) | NS |
| ETH010 | 4 | 2.06 | 0.516 | 0.434 | 0.44 | 0.15805 | (0.00882–0.30514) | NS |
| ETH225 | 6 | 1.30 | 0.233 | 0.214 | 0.22 | 0.08139 | (−0.06547–0.23836) | NS |
| HAUT27 | 7 | 3.68 | 0.731 | 0.622 | 0.7 | 0.14921 | (0.05547–0.23469) | NS |
| HSC | 14 | 8.42 | 0.885 | 0.789 | 0.87 | 0.10785 | (0.03307–0.18731) | NS |
| ILSTS008 | 3 | 1.60 | 0.375 | 0.352 | 0.34 | 0.06315 | (−0.06421–0.19046) | NS |
| ILSTS011 | 8 | 1.95 | 0.49 | 0.469 | 0.43 | 0.04338 | (−0.08673–0.17046) | NS |
| ILSTS019 | 7 | 4.42 | 0.776 | 0.681 | 0.74 | 0.12382 | (0.02715–0.22251) | NS |
| ILSTS030 | 9 | 6.01 | 0.837 | 0.667 | 0.81 | 0.20378 | (0.10995–0.29227) | NS |
| ILSTS087 | 7 | 3.16 | 0.686 | 0.667 | 0.64 | 0.02767 | (−0.06163–0.11632) | NS |
| INRA005 | 4 | 2.58 | 0.615 | 0.621 | 0.55 | −0.00962 | (−0.12535–0.09767) | NS |
| INRA006 | 9 | 7.63 | 0.872 | 0.671 | 0.86 | 0.23073 | (0.13451–0.31479) | NS |
| INRA023 | 7 | 5.73 | 0.828 | 0.507 | 0.8 | 0.38867 | (0.28331–0.48339) | *** |
| INRA063 | 5 | 2.50 | 0.601 | 0.462 | 0.52 | 0.23224 | (0.10057–0.35608) | NS |
| INRA172 | 7 | 3.78 | 0.738 | 0.655 | 0.69 | 0.11342 | (0.01843–0.20403) | NS |
| MAF065 | 10 | 4.28 | 0.769 | 0.664 | 0.74 | 0.13723 | (0.03830–0.23089) | NS |
| MAF209 | 2 | 1.52 | 0.341 | 0.269 | 0.28 | 0.21245 | (0.02267–0.38747) | NS |
| McM527 | 9 | 4.20 | 0.765 | 0.669 | 0.73 | 0.12542 | (0.01993–0.21898) | NS |
| MM12 | 14 | 7.46 | 0.869 | 0.813 | 0.85 | 0.06528 | (−0.00962–0.13071) | NS |
| OarFCB011 | 10 | 4.58 | 0.784 | 0.734 | 0.75 | 0.064 | (−0.02090–0.15133) | NS |
| OarFCB020 | 6 | 1.70 | 0.414 | 0.372 | 0.38 | 0.0999 | (−0.04615–0.23000) | NS |
| OarFCB048 | 11 | 4.69 | 0.789 | 0.746 | 0.76 | 0.05456 | (−0.02320–0.13311) | NS |
| OarFCB304 | 12 | 4.77 | 0.793 | 0.738 | 0.76 | 0.06993 | (−0.01207–0.15543) | NS |
| SPS115 | 6 | 1.93 | 0.488 | 0.549 | 0.4 | −0.13716 | (−0.27617–0.00121) | NS |
| SRCRSP05 | 7 | 5.47 | 0.82 | 0.759 | 0.79 | 0.07493 | (−0.01091–0.15903) | NS |
| SRCRSP08 | 6 | 5.07 | 0.772 | 0.643 | 0.73 | 0.16705 | (0.05626–0.26468) | NS |
| SRCRSP23 | 11 | 3.69 | 0.805 | 0.722 | 0.78 | 0.10356 | (0.01086–0.18899) | NS |
| SRCRSP24 | 8 | 4.33 | 0.732 | 0.558 | 0.69 | 0.23787 | (0.11786–0.33931) | ** |
| TGLA053 | 7 | 3.58 | 0.723 | 0.722 | 0.68 | 0.00138 | (−0.09825–0.08853) | NS |
| TGLA122 | 7 | 3.09 | 0.679 | 0.587 | 0.64 | 0.13537 | (0.02766–0.23688) | NS |
| 8 | 4.3 | 0.71 | 0.63 | 0.67 | 0,10986 | (0.08773–0.12549) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguirre-Riofrio, L.; Maza-Tandazo, T.; Quezada-Padilla, M.; Albito-Balcazar, O.; Flores-Gonzalez, A.; Camacho-Enriquez, O.; Martinez-Martinez, A.; Consortium, B.; Delgado-Bermejo, J.V. Genetic Characterization of the “Chusca Lojana”, a Creole Goat Reared in Ecuador, and Its Relationship with Other Goat Breeds. Animals 2020, 10, 1026. https://doi.org/10.3390/ani10061026
Aguirre-Riofrio L, Maza-Tandazo T, Quezada-Padilla M, Albito-Balcazar O, Flores-Gonzalez A, Camacho-Enriquez O, Martinez-Martinez A, Consortium B, Delgado-Bermejo JV. Genetic Characterization of the “Chusca Lojana”, a Creole Goat Reared in Ecuador, and Its Relationship with Other Goat Breeds. Animals. 2020; 10(6):1026. https://doi.org/10.3390/ani10061026
Chicago/Turabian StyleAguirre-Riofrio, Lenin, Teddy Maza-Tandazo, Manuel Quezada-Padilla, Oscar Albito-Balcazar, Alex Flores-Gonzalez, Osvaldo Camacho-Enriquez, Amparo Martinez-Martinez, BioGoat Consortium, and Juan Vicente Delgado-Bermejo. 2020. "Genetic Characterization of the “Chusca Lojana”, a Creole Goat Reared in Ecuador, and Its Relationship with Other Goat Breeds" Animals 10, no. 6: 1026. https://doi.org/10.3390/ani10061026
APA StyleAguirre-Riofrio, L., Maza-Tandazo, T., Quezada-Padilla, M., Albito-Balcazar, O., Flores-Gonzalez, A., Camacho-Enriquez, O., Martinez-Martinez, A., Consortium, B., & Delgado-Bermejo, J. V. (2020). Genetic Characterization of the “Chusca Lojana”, a Creole Goat Reared in Ecuador, and Its Relationship with Other Goat Breeds. Animals, 10(6), 1026. https://doi.org/10.3390/ani10061026