Impact of Maternal High Stocking Density during the Dry Period on Dairy Calf Health, Behaviour, and Welfare


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing and Feeding
2.2. Data Collection and Data Processing
2.2.1. Body Weight and Average Daily Gains
2.2.2. Activity Levels
2.2.3. Training for the Use of an Automatic Milk Feeder
2.2.4. Reactions to a Novel Group Environment, Behaviour for the First Two Weeks in the Group Pen and Reactions to Weaning
2.2.5. Behavioural and Physiological Reactions to Disbudding
2.3. Statistical Analysis
2.3.1. Individual Statistical Model Development
2.3.2. BW, ADG, IgG, Training Count, LP, Reactions to a Novel Group Environment
2.3.3. Behaviour in the First Two Weeks in the Group Pen and around Weaning
2.3.4. Behavioural and Physiological Reactions to Disbudding
3. Results
3.1. BW, ADG, Health and IgG Levels
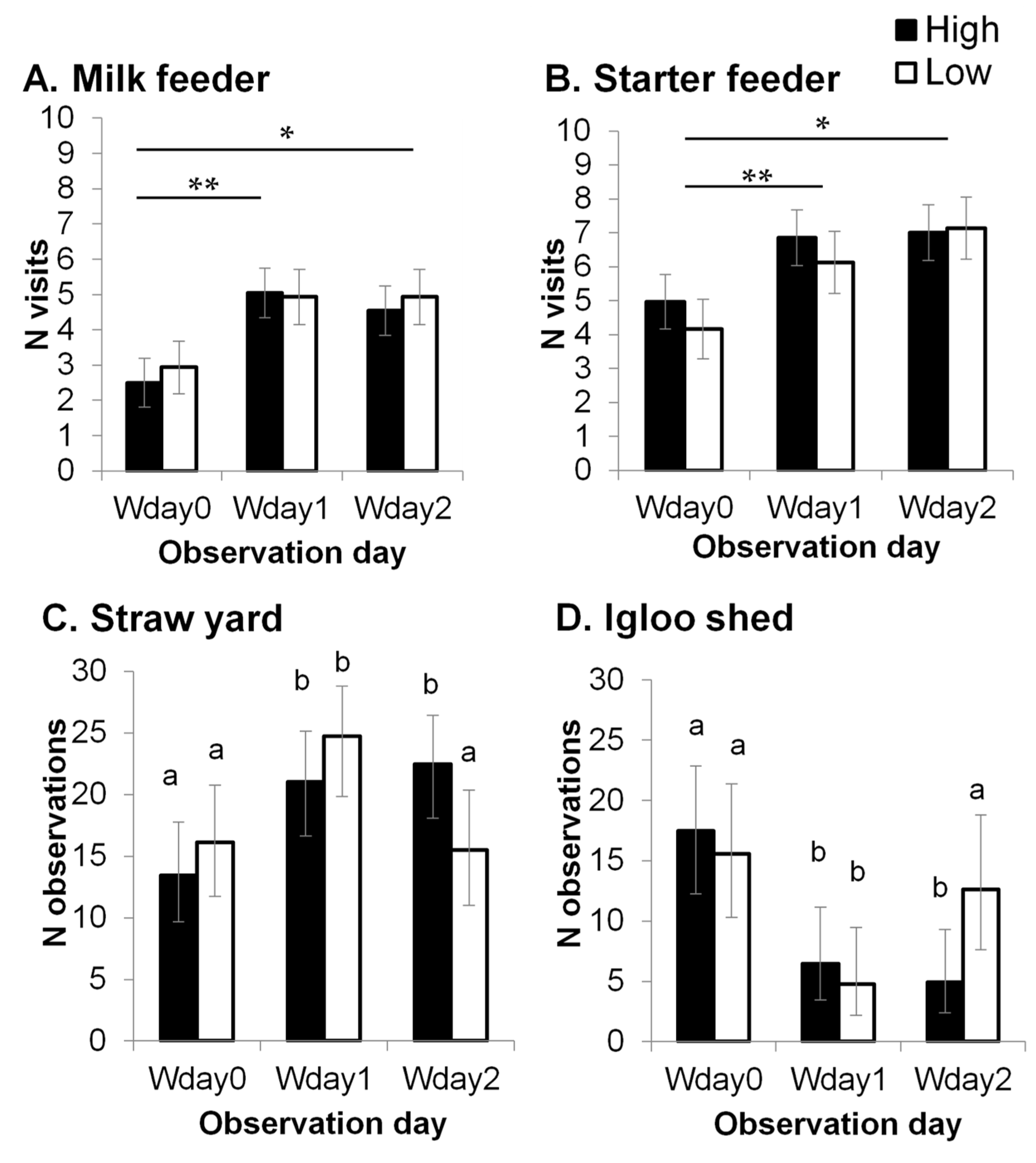
3.2. Activity Levels and Training Count for the Use of an Automatic Milk Feeder
3.3. Reactions to the Group Pen
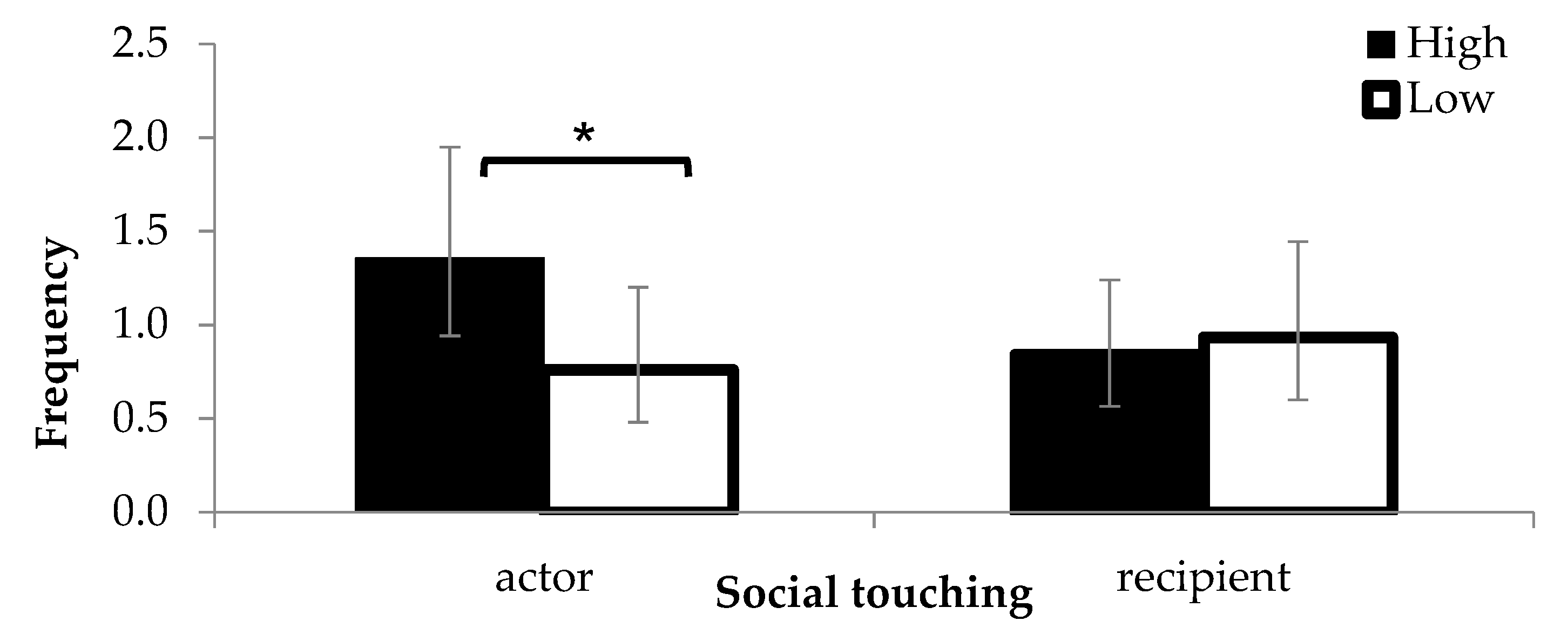
3.4. Behaviour in the First Two Weeks in the Group Pen
3.5. Reactions to Weaning
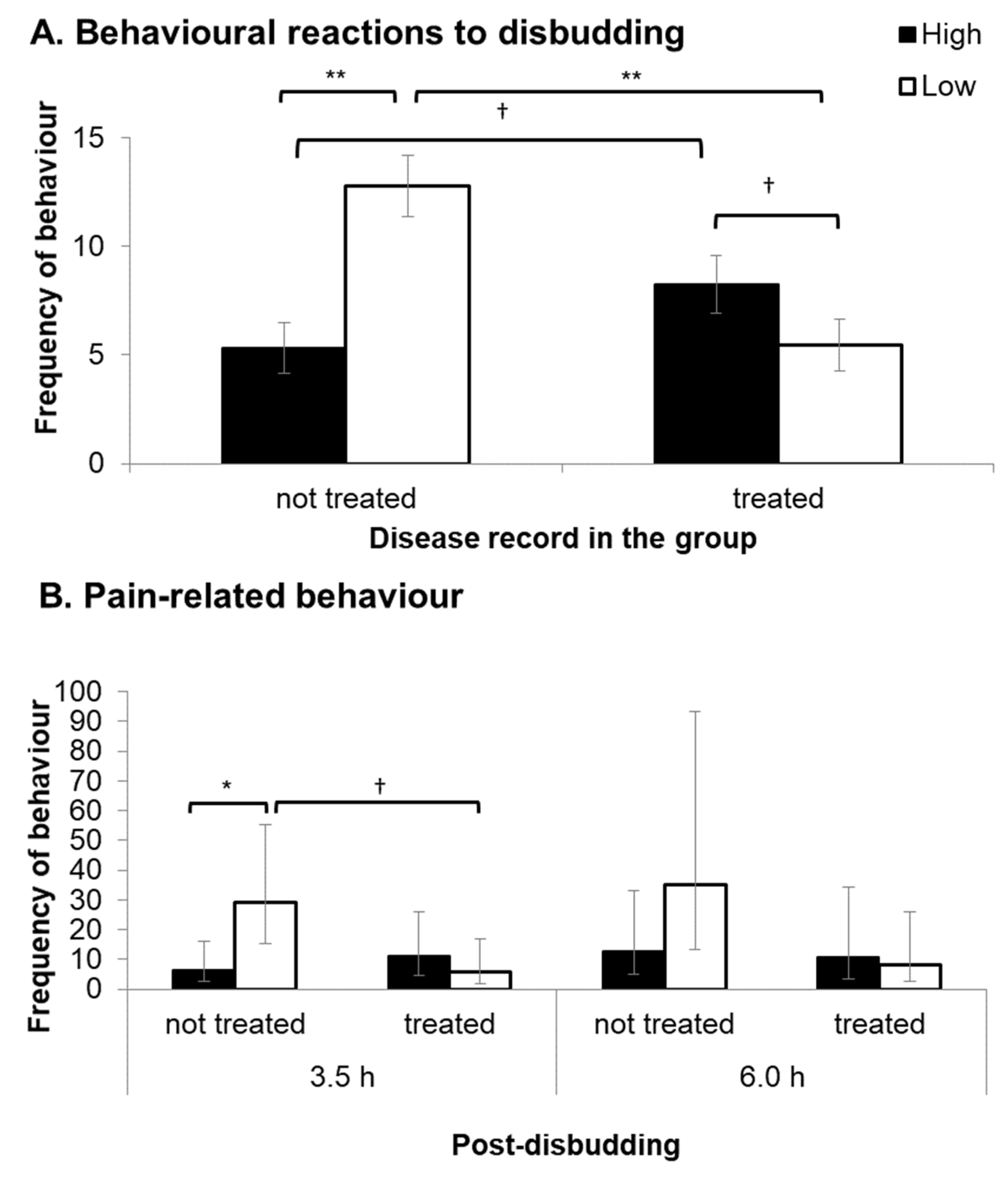
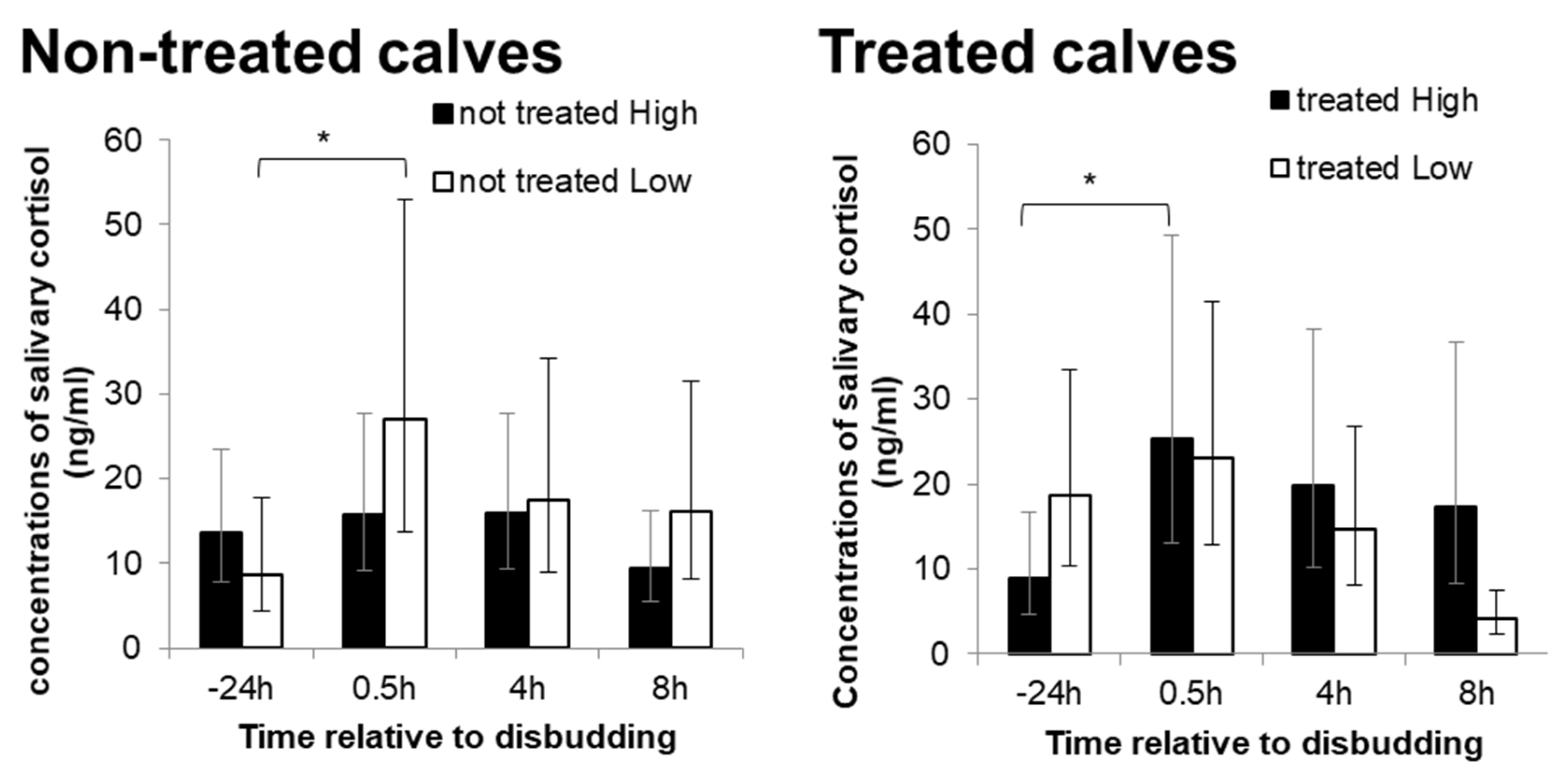
3.6. Reactions to Disbudding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gao, F.; Liu, Y.-C.; Zhang, Z.-H.; Zhang, C.-Z.; Su, H.-W.; Li, S.-L. Effect of prepartum maternal energy density on the growth performance, immunity, and antioxidation capability of neonatal calves. J. Dairy Sci. 2012, 95, 4510–4518. [Google Scholar] [CrossRef]
- Weinstock, M. The long-term behavioural consequences of prenatal stress. Neurosci. Biobehav. Rev. 2008, 32, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, K.D.; Rutherford, K.M.D.; Wallace, J.M.; Brameld, J.M.; Stöger, R.; Alberio, R.; Sweetman, D.; Gardner, D.S.; Perry, V.E.A.; Adam, C.L.; et al. Epigenetics and developmental programming of welfare and production traits in farm animals. Reprod. Fertil. Dev. 2016, 28, 1443–1478. [Google Scholar] [CrossRef] [Green Version]
- Charil, A.; Laplante, D.P.; Vaillancourt, C.; King, S. Prenatal stress and brain development. Brain Res. Rev. 2010, 65, 56–79. [Google Scholar] [CrossRef] [PubMed]
- Brunton, P.J. Effects of maternal exposure to social stress during pregnancy: Consequences for mother and offspring. Reproduction 2013, 146, R175–R189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braastad, B.O. Effects of prenatal stress on behaviour of offspring of laboratory and farmed mammals. Appl. Anim. Behav. Sci. 1998, 61, 159–180. [Google Scholar] [CrossRef]
- Rutherford, K.M.D.; Donald, R.D.; Arnott, G.; Rooke, J.A.; Dixon, L.; Mehers, J.J.M.; Turnbull, J.; Lawrence, A.B. Farm animal welfare: Assessing risks attributable to the prenatal environment. Anim. Welf. 2012, 21, 419–429. [Google Scholar] [CrossRef]
- Arnott, G.; Roberts, D.; Rooke, J.A.; Turner, S.P.; Lawrence, A.B.; Rutherford, K.M.D. Board invited review: The importance of the gestation period for welfare of calves: Maternal stressors and difficult births. J. Anim. Sci. 2012, 90, 5021–5034. [Google Scholar] [CrossRef] [Green Version]
- Otten, W.; Kanitz, E.; Tuchscherer, M. The impact of pre-natal stress on offspring development in pigs. J. Agric. Sci. 2015, 153, 907–919. [Google Scholar] [CrossRef]
- Rooke, J.A.; Arnott, G.; Dwyer, C.M.; Rutherford, K.M.D. The importance of the gestation period for welfare of lambs: Maternal stressors and lamb vigour and wellbeing. J. Agric. Sci. 2015, 153, 497–519. [Google Scholar] [CrossRef]
- Dixon, L.M.; Sparks, N.H.C.; Rutherford, K.M.D. Early experiences matter: A review of the effects of prenatal environment on offspring characteristics in poultry. Poult. Sci. 2016, 95, 489–499. [Google Scholar] [CrossRef]
- Roussel, S.; Hemsworth, P.H.; Boissy, A.; Duvaux-Ponter, C. Effects of repeated stress during pregnancy in ewes on the behavioural and physiological responses to stressful events and birth weight of their offspring. Appl. Anim. Behav. Sci. 2004, 85, 259–276. [Google Scholar] [CrossRef]
- Roussel-Huchette, S.; Hemsworth, P.H.; Boissy, A.; Duvaux-Ponter, C. Repeated transport and isolation during pregnancy in ewes: Differential effects on emotional reactivity and weight of their offspring. Appl. Anim. Behav. Sci. 2008, 109, 275–291. [Google Scholar] [CrossRef]
- Ison, S.H.; D’Eath, R.B.; Robson, S.K.; Baxter, E.M.; Ormandy, E.; Douglas, A.J.; Russell, J.A.; Lawrence, A.B.; Jarvis, S. “Subordination style” in pigs? The response of pregnant sows to mixing stress affects their offspring’s behaviour and stress reactivity. Appl. Anim. Behav. Sci. 2010, 124, 16–27. [Google Scholar] [CrossRef]
- Adin, G.; Gelman, A.; Solomon, R.; Flamenbaum, I.; Nikbachat, M.; Yosef, E.; Zenou, A.; Shamay, A.; Feuermann, Y.; Mabjeesh, S.J.; et al. Effects of cooling dry cows under heat load conditions on mammary gland enzymatic activity, intake of food and water, and performance during the dry period and after parturition. Livest. Sci. 2009, 124, 189–195. [Google Scholar] [CrossRef]
- do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, J.; Bubolz, J.; Dahl, G.E. Heat-stress abatement during the dry period: Does cooling improve transition into lactation? J. Dairy Sci. 2009, 92, 5988–5999. [Google Scholar] [CrossRef]
- do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, M.J.; Bubolz, J.W.; Dahl, G.E. Heat stress abatement during the dry period influences metabolic gene expression and improves immune status in the transition period of dairy cows. J. Dairy Sci. 2011, 94, 86–96. [Google Scholar] [CrossRef]
- Wolfenson, D.; Flamenbaum, I.; Berman, A. Dry period heat stress relief effects on prepartum progesterone, calf birth weight, and milk production. J. Dairy Sci. 1988, 71, 809–818. [Google Scholar] [CrossRef]
- Collier, R.J.; Doelger, S.G.; Head, H.H.; Thatcher, W.W.; Wilcox, C.J. Effects of heat stress during pregnancy on maternal hormone concentrations, calf birth weight and postpartum milk yield of Holstein cows. J. Anim. Sci. 1982, 54, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Monteiro, A.P.A.; Thompson, I.M.; Hayen, M.J.; Dahl, G.E. Effect of late-gestation maternal heat stress on growth and immune function of dairy calves. J. Dairy Sci. 2012, 95, 7128–7136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; Bubolz, J.W.; do Amaral, B.C.; Thompson, I.M.; Hayen, M.J.; Johnson, S.E.; Dahl, G.E. Effect of heat stress during the dry period on mammary gland development. J. Dairy Sci. 2011, 94, 5976–5986. [Google Scholar] [CrossRef] [PubMed]
- Lay, D.C.; Randel, R.D.; Friend, T.H.; Jenkins, O.C.; Neuendorff, D.A.; Bushong, D.M.; Lanier, E.K.; Bjorge, M.K. Effects of prenatal stress on suckling calves. J. Anim. Sci. 1997, 75, 3143–3151. [Google Scholar] [CrossRef]
- Littlejohn, B.P.; Price, D.M.; Banta, J.P.; Lewis, A.W.; Neuendorff, D.A.; Carroll, J.A.; Vann, R.C.; Welsh, T.H.; Randel, R.D. Prenatal transportation stress alters temperament and serum cortisol concentrations in suckling Brahman calves12. J. Anim. Sci. 2016, 94, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, M.; Haskell, M.J.; Macrae, A.I.; Rutherford, K.M.D. Survey of dry cow management on UK commercial dairy farms. Vet. Rec. 2018, 183, 297. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Dahl, G.E. Invited review: Heat stress effects during late gestation on dry cows and their calves. J. Dairy Sci. 2013, 96, 4079–4093. [Google Scholar] [CrossRef] [PubMed]
- Zobel, G.; Weary, D.M.; Leslie, K.E.; von Keyserlingk, M.A.G. Invited review: Cessation of lactation: Effects on animal welfare. J. Dairy Sci. 2015, 98, 8263–8277. [Google Scholar] [CrossRef] [Green Version]
- Huzzey, J.M.; DeVries, T.J.; Valois, P.; von Keyserlingk, M.A.G. Stocking density and feed barrier design affect the feeding and social behavior of dairy cattle. J. Dairy Sci. 2006, 89, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.R.B.; Moraes, J.G.N.; Mendonça, L.G.D.; Scanavez, A.A.; Nakagawa, G.; Fetrow, J.; Endres, M.I.; Chebel, R.C. Effects of weekly regrouping of prepartum dairy cows on metabolic, health, reproductive, and productive parameters. J. Dairy Sci. 2013, 96, 4436–4446. [Google Scholar] [CrossRef]
- Black, R.A.; Whitlock, B.K.; Krawczel, P.D. Effect of maternal exercise on calf dry matter intake, weight gain, behavior, and cortisol concentrations at disbudding and weaning. J. Dairy Sci. 2017, 100, 7390–7400. [Google Scholar] [CrossRef]
- DeVries, T.J.; von Keyserlingk, M.A.G.; Weary, D.M. Effect of feeding space on the inter-cow distance, aggression, and feeding behavior of free-stall housed lactating dairy cows. J. Dairy Sci. 2004, 87, 1432–1438. [Google Scholar] [CrossRef]
- Proudfoot, K.L.; Veira, D.M.; Weary, D.M.; von Keyserlingk, M.A.G. Competition at the feed bunk changes the feeding, standing, and social behavior of transition dairy cows. J. Dairy Sci. 2009, 92, 3116–3123. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, M.; Haskell, M.; Macrae, A.; Rutherford, K. Effects of stocking density during the dry period on dairy cow physiology, metabolism and behaviour. J. Dairy Res. 2019, 86, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Lay, D.C.; Randel, R.D.; Friend, T.H.; Carroll, J.A.; Welsh, T.H.; Jenkins, O.C.; Neuendorff, D.A.; Bushong, D.M.; Kapp, G.M. Effects of prenatal stress on the fetal calf. Domest. Anim. Endocrinol. 1997, 14, 73–80. [Google Scholar] [CrossRef]
- Andreoli, K.M.; Minton, J.E.; Spire, M.F.; Schalles, R.R. Influence of prepartum exposure of beef heifers to winter weather on concentrations of plasma energy-yielding substrates, serum hormones and birth weight of calves. Theriogenology 1988, 29, 631–642. [Google Scholar] [CrossRef]
- Jensen, M.B. Effects of confinement on rebounds of locomotor behaviour of calves and heifers, and the spatial preferences of calves. Appl. Anim. Behav. Sci. 1999, 62, 43–56. [Google Scholar] [CrossRef]
- Krachun, C.; Rushen, J.; de Passillé, A.M. Play behaviour in dairy calves is reduced by weaning and by a low energy intake. Appl. Anim. Behav. Sci. 2010, 122, 71–76. [Google Scholar] [CrossRef]
- Graf, B.; Senn, M. Behavioural and physiological responses of calves to dehorning by heat cauterization with or without local anaesthesia. Appl. Anim. Behav. Sci. 1999, 62, 153–171. [Google Scholar] [CrossRef]
- Faulkner, P.M.; Weary, D.M. Reducing pain after dehorning in dairy calves. J. Dairy Sci. 2000, 83, 2037–2041. [Google Scholar] [CrossRef]
- Heinrich, A.; Duffield, T.F.; Lissemore, K.D.; Millman, S.T. The effect of meloxicam on behavior and pain sensitivity of dairy calves following cautery dehorning with a local anesthetic. J. Dairy Sci. 2010, 93, 2450–2457. [Google Scholar] [CrossRef] [Green Version]
- Haskell, M.J.; Langford, F.M.; Jack, M.C.; Sherwood, L.; Lawrence, A.B.; Rutherford, K.M.D. The effect of organic status and management practices on somatic cell counts on UK dairy farms. J. Dairy Sci. 2009, 92, 3775–3780. [Google Scholar] [CrossRef]
- Stafford, K.J.; Mellor, D.J. Addressing the pain associated with disbudding and dehorning in cattle. Appl. Anim. Behav. Sci. 2011, 135, 226–231. [Google Scholar] [CrossRef]
- Knaepen, L.; Pawluski, J.L.; Patijn, J.; van Kleef, M.; Tibboel, D.; Joosten, E.A. Perinatal maternal stress and serotonin signaling: Effects on pain sensitivity in offspring. Dev. Psychobiol. 2014, 56, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, K.M.D.; Robson, S.K.; Donald, R.D.; Jarvis, S.; Sandercock, D.A.; Scott, E.M.; Nolan, A.M.; Lawrence, A.B. Pre-natal stress amplifies the immediate behavioural responses to acute pain in piglets. Biol. Lett. 2009, 5, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Butkevich, I.P.; Vershinina, E.A. Prenatal stress alters time characteristics and intensity of formalin-induced pain responses in juvenile rats. Brain Res. 2001, 915, 88–93. [Google Scholar] [CrossRef]
- Cramer, M.C.; Stanton, A.L. Associations between health status and the probability of approaching a novel object or stationary human in preweaned group-housed dairy calves. J. Dairy Sci. 2015, 98, 7298–7308. [Google Scholar] [CrossRef] [Green Version]
- Weary, D.M.; Huzzey, J.M.; von Keyserlingk, M.A.G. Board-invited review: Using behavior to predict and identify ill health in animals. J. Anim. Sci. 2009, 87, 770–777. [Google Scholar] [CrossRef] [Green Version]
- Hart, B.L. Biological basis of the behavior of sick animals. Neurosci. Biobehav. Rev. 1988, 12, 123–137. [Google Scholar] [CrossRef]
- Hixson, C.L.; Krawczel, P.D.; Caldwell, J.M.; Miller-Cushon, E.K. Behavioral changes in group-housed dairy calves infected with Mannheimia haemolytica. J. Dairy Sci. 2018, 101, 10351–10360. [Google Scholar] [CrossRef] [Green Version]
- Windeyer, M.C.; Leslie, K.E.; Godden, S.M.; Hodgins, D.C.; Lissemore, K.D.; LeBlanc, S.J. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Prev. Vet. Med. 2014, 113, 231–240. [Google Scholar] [CrossRef]
- Virtala, A.-M.K.; Mechor, G.D.; Gröhn, Y.T.; Erb, H.N. The effect of calfhood diseases on growth of female dairy calves during the first 3 months of life in New York State. J. Dairy Sci. 1996, 79, 1040–1049. [Google Scholar] [CrossRef]
- Weary, D.M.; Jasper, J.; Hötzel, M.J. Understanding weaning distress. Appl. Anim. Behav. Sci. 2008, 110, 24–41. [Google Scholar] [CrossRef]
- de Paula Vieira, A.; Guesdon, V.; de Passillé, A.M.; von Keyserlingk, M.A.G.; Weary, D.M. Behavioural indicators of hunger in dairy calves. Appl. Anim. Behav. Sci. 2008, 109, 180–189. [Google Scholar] [CrossRef]
- Budzynska, M.; Weary, D.M. Weaning distress in dairy calves: Effects of alternative weaning procedures. Appl. Anim. Behav. Sci. 2008, 112, 33–39. [Google Scholar] [CrossRef]
- Jarvis, S.; Moinard, C.; Robson, S.K.; Baxter, E.M.; Ormandy, E.; Douglas, A.J.; Seckl, J.R.; Russell, J.A.; Lawrence, A.B. Programming the offspring of the pig by prenatal social stress: Neuroendocrine activity and behaviour. Horm. Behav. 2006, 49, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Boissy, A.; Bouissou, M.-F. Assessment of individual differences in behavioural reactions of heifers exposed to various fear-eliciting situations. Appl. Anim. Behav. Sci. 1995, 46, 17–31. [Google Scholar] [CrossRef]
- Jensen, M.B.; Vestergaard, K.S.; Krohn, C.C.; Munksgaard, L. Effect of single versus group housing and space allowance on responses of calves during open-field tests. Appl. Anim. Behav. Sci. 1997, 54, 109–121. [Google Scholar] [CrossRef]
- Holm, L.; Jensen, M.B.; Jeppesen, L.L. Calves’ motivation for access to two different types of social contact measured by operant conditioning. Appl. Anim. Behav. Sci. 2002, 79, 175–194. [Google Scholar] [CrossRef]
- Costa, J.H.C.; von Keyserlingk, M.A.G.; Weary, D.M. Invited review: Effects of group housing of dairy calves on behavior, cognition, performance, and health. J. Dairy Sci. 2016, 99, 2453–2467. [Google Scholar] [CrossRef]
- Gaillard, C.; Meagher, R.K.; von Keyserlingk, M.A.G.; Weary, D.M. Social housing improves dairy calves’ performance in two cognitive tests. PLoS ONE 2014, 9, e90205. [Google Scholar] [CrossRef]
- Buchli, C.; Raselli, A.; Bruckmaier, R.; Hillmann, E. Contact with cows during the young age increases social competence and lowers the cardiac stress reaction in dairy calves. Appl. Anim. Behav. Sci. 2017, 187, 1–7. [Google Scholar] [CrossRef]
- de Paula Vieira, A.; de Passillé, A.M.; Weary, D.M. Effects of the early social environment on behavioral responses of dairy calves to novel events. J. Dairy Sci. 2012, 95, 5149–5155. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Definition | Observation |
|---|---|---|
| Milk feeder | At least one front foot of the calf is inside the feeder stall. | 1, 2, 3 |
| Starter feeder (group pen) | The calf’s head is within a half body length from the line between the starter feeder and the straw-bedded area. | 2 |
| Starter feeder (weaning) | The calf’s head is fully across the line between the straw-bedded area and the starter trough, and remained there for more than five seconds. The starter feeder visit is finished when the calf’s head is away from the starter trough for more than ten seconds. | 3 |
| Straw yard | At least one foot of the calf is in the straw-bedded area. | 1, 2, 3 |
| Igloo | Four legs of the calf cross the front line of the igloo. | 1, 2, 3 |
| Water dispenser | The muzzle of the calf touches or is within 5 cm from the water dispenser. | 3 |
| Posture | ||
| Standing inactive | The calf is in an upright position with no leg movement. | 1 |
| Standing | The calf is in an upright position including walking and running. | 2, 3 |
| Lying | The calf lies down with sternal recumbency or lies on its flank. | 1, 2, 3 |
| Walking | The calf makes a forward movement with more than two steps. Two or three hoofs are touching the ground at any time. | 1 |
| Running | The calf makes a rapid forward movement, including instances of jumping, bucking, galloping and trotting [35,36]. | 1 |
| Behaviour | ||
| Explore | The muzzle of the calf is in contact with or within approximately 5 cm from the wall, floor, chain, straw feeder, water dispenser, starter or any other objects in the pen. | 1 |
| Social contact initiated | The muzzle of the calf is in contact with or within approximately 5 cm from the head or body of another calf. | 1 |
| Social contact received | The muzzle of another calf (calves) is (are) in contact or within 5 cm from any parts of the focal calf (lasting 3 s or more). | 1 |
| No contact | The muzzle of the calf is not in contact with any objects in the pen, any parts of another calf’s body or its own body. | 1 |
| Touch | ||
| Social touch actor | The muzzle of the focal calf is in contact with any parts of another calf. Head and neck oriented towards recipient. | 2 |
| Social touch recipient | The muzzle of other calf (calves) is/are in contact with any body parts of the focal calf. | 2 |
| Proximity | ||
| Close | Distance to neighbouring calf (calves) is within one body width. | 2 |
| Not close | Distance to neighbouring calf (calves) is more than one body width. | 2 |
| Event/state | ||
| Human presence | There are human(s) inside or outside of the front face of the pen. | 1, 2, 3 * |
| Measurement | High Stocking | Low Stocking | Test Statistics | p-Value | |
|---|---|---|---|---|---|
| Body weight (kg) | birth | 45.4 ± 1.0 | 46.5 ± 1.1 | W = 0.6 | 0.439 |
| day 7 | 45.1 ± 0.9 | 46.1 ± 1.0 | W = 0.5 | 0.492 | |
| wean | 69.0 ± 1.5 | 69.8 ± 1.7 | W = 0.1 | 0.759 | |
| ADG (g/day) | hutch | −25.6 ± 57.1 | −5.3 ± 62.6 | W = 0.1 | 0.812 |
| group | 578.7 ± 27.1 | 587.1 ± 32.0 | W < 0.1 | 0.844 | |
| pre-weaning | 493.2 ± 25.8 | 489.0 ± 30.3 | W < 0.1 | 0.915 | |
| IgG levels (mg/mL) on day 7 | 7.9 ± 0.8 | 8.1 ± 0.9 | W < 0.1 | 0.911 | |
| LP hutch | day 2 | 0.88 ± 0.02 | 0.90 ± 0.02 | Treatment: F1,34 = 1.0 | 0.338 |
| day 3 | 0.86 ± 0.02 | 0.87 ± 0.02 | Day: F4,125 = 9.4 | <0.001 | |
| day 4 | 0.85 ± 0.02 | 0.85 ± 0.02 | |||
| day 5 | 0.84 ± 0.02 | 0.84 ± 0.02 | |||
| day 6 | 0.84 ± 0.02 | 0.86 ± 0.02 | |||
| LP group | group day 1 * | 0.82 ± 0.02 | 0.80 ± 0.02 | Treatment × Day: F1,33 = 6.0 | 0.019 |
| group day 2 * | 0.81 ± 0.02 | 0.83 ± 0.02 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujiwara, M.; Haskell, M.J.; Macrae, A.I.; Rutherford, K.M.D. Impact of Maternal High Stocking Density during the Dry Period on Dairy Calf Health, Behaviour, and Welfare. Animals 2020, 10, 922. https://doi.org/10.3390/ani10060922
Fujiwara M, Haskell MJ, Macrae AI, Rutherford KMD. Impact of Maternal High Stocking Density during the Dry Period on Dairy Calf Health, Behaviour, and Welfare. Animals. 2020; 10(6):922. https://doi.org/10.3390/ani10060922
Chicago/Turabian StyleFujiwara, Mayumi, Marie J. Haskell, Alastair I. Macrae, and Kenneth M. D. Rutherford. 2020. "Impact of Maternal High Stocking Density during the Dry Period on Dairy Calf Health, Behaviour, and Welfare" Animals 10, no. 6: 922. https://doi.org/10.3390/ani10060922
APA StyleFujiwara, M., Haskell, M. J., Macrae, A. I., & Rutherford, K. M. D. (2020). Impact of Maternal High Stocking Density during the Dry Period on Dairy Calf Health, Behaviour, and Welfare. Animals, 10(6), 922. https://doi.org/10.3390/ani10060922