Transcriptome Profiling Associated with Carcass Quality of Loin Muscles in Crossbred Pigs



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Library Preparation and Data Generation
2.3. Data Analysis
3. Results
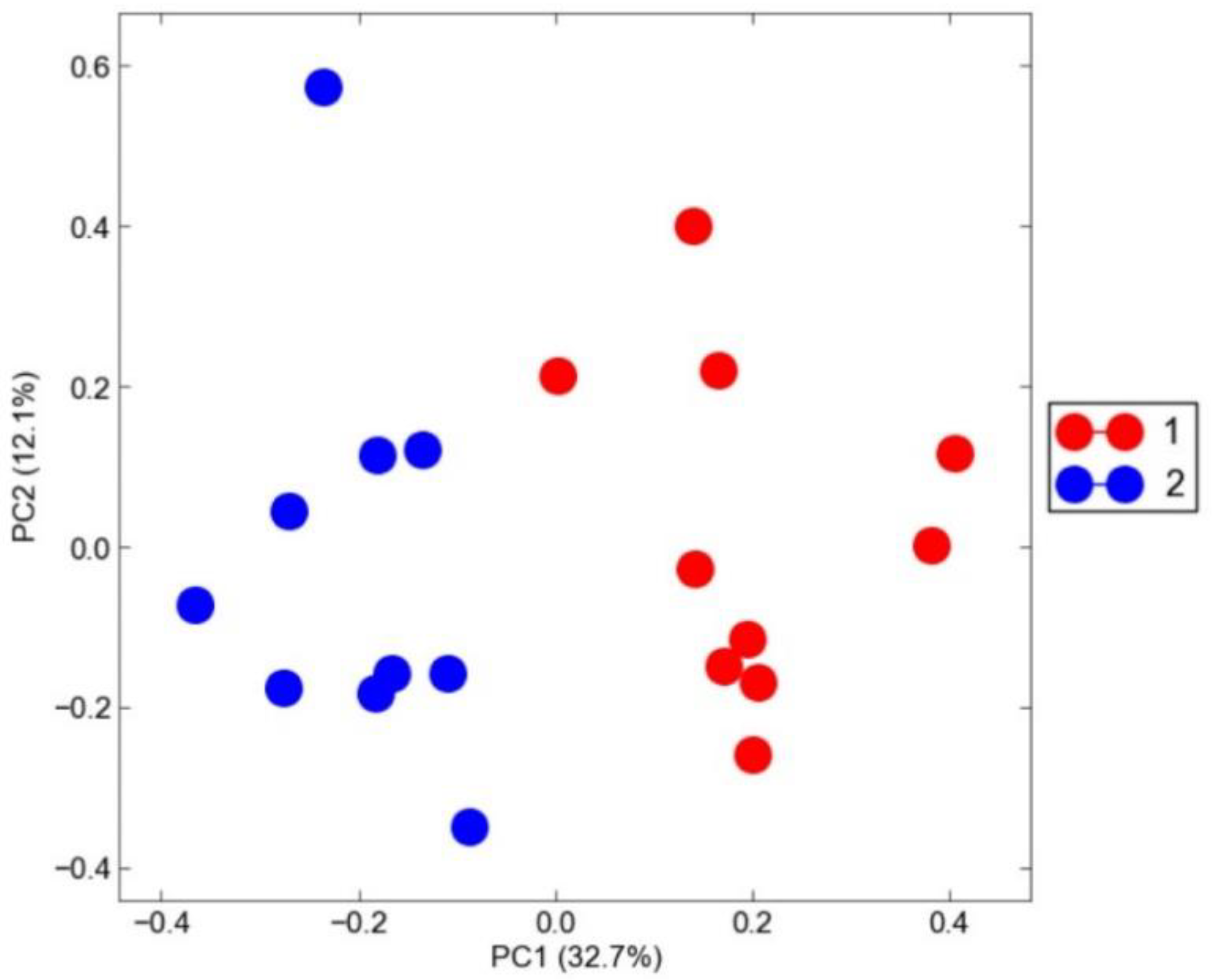
3.1. Pig Loin Muscle Transcriptome Profiling
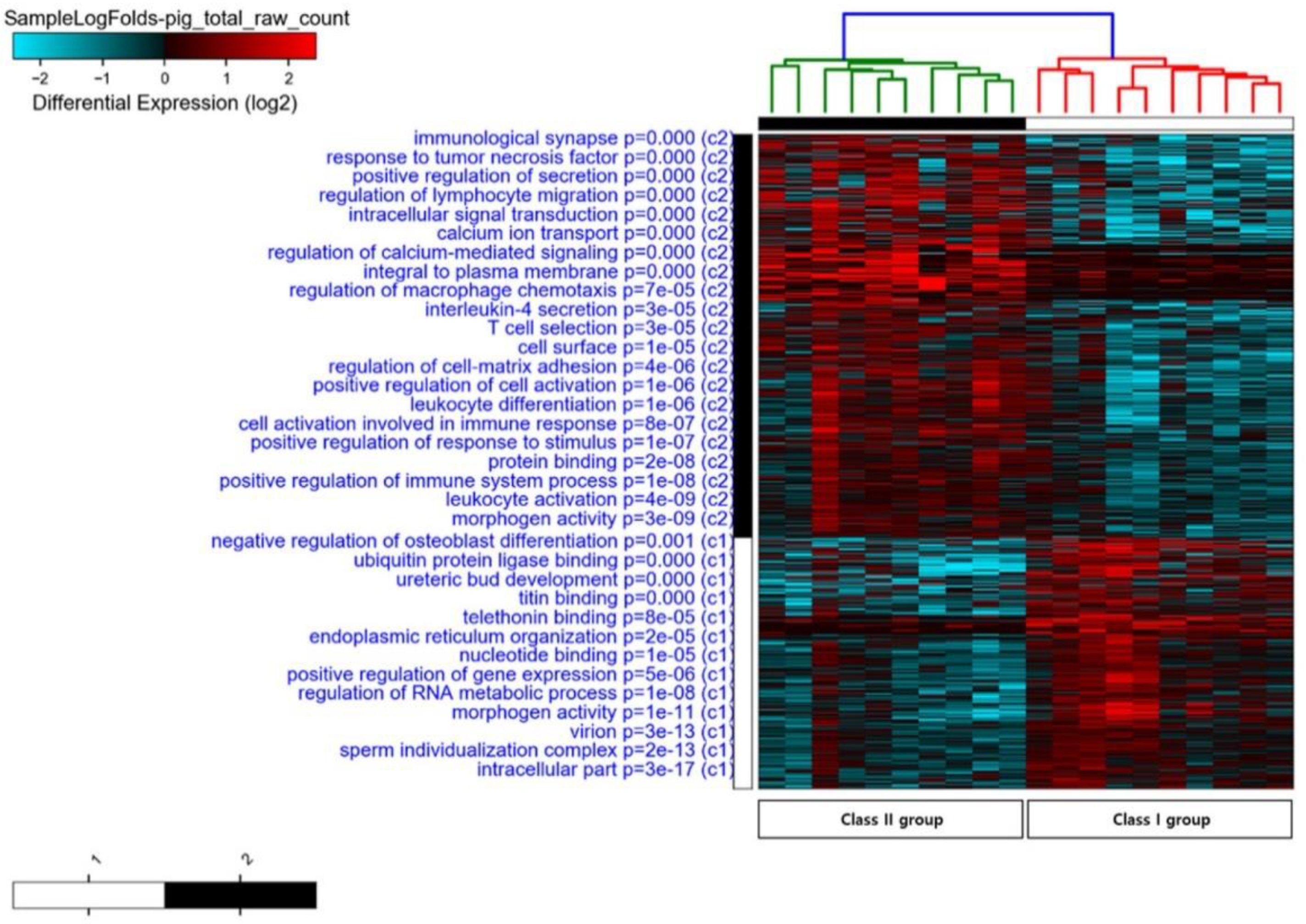
3.2. Putative Candidate Genes Involved in Loin Muscle Carcass Quality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- OECD/FAO. Meat. In OECD-FAO Agricultural Outlook 2019-2028; OECD Publishing: Paris, French, 2019; pp. 166–179. [Google Scholar]
- Oh, S.H.; See, M.T. Pork preference for consumers in China, Japan and South Korea. Asian-Australas. J. Anim. Sci. 2012, 25, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, J.-H.; Yang, H.-S.; Lee, S.-H.; Go, G.-W. Characteristics of pork belly consumption in south korea and their health implication. J. Anim. Sci. Technol. 2015, 57, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Weng, Q.; Dong, C.; Zhang, Z.; Li, R.; Liu, J.; Jiang, A.; Li, Q.; Jia, C.; Wu, W.; et al. A key gene, plin1, can affect porcine intramuscular fat content based on transcriptome analysis. Genes (Basel) 2018, 9, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Zhang, W.; Tume, R.K.; Hudson, N.J.; Huang, F.; Yin, Y.; Zhou, G. Disorder of endoplasmic reticulum calcium channel components is associated with the increased apoptotic potential in pale, soft, exudative pork. Meat Sci. 2016, 115, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Lee, J.H.; Kim, S.W.; Han, J.S.; Kang, K.S.; Kim, S.J.; Park, T.S. Muscle differentiation induced up-regulation of calcium-related gene expression in quail myoblasts. Asian-Australas. J. Anim. Sci. 2018, 31, 1507–1515. [Google Scholar] [CrossRef]
- Hamill, R.M.; McBryan, J.; McGee, C.; Mullen, A.M.; Sweeney, T.; Talbot, A.; Cairns, M.T.; Davey, G.C. Functional analysis of muscle gene expression profiles associated with tenderness and intramuscular fat content in pork. Meat Sci. 2012, 92, 440–450. [Google Scholar] [CrossRef]
- Zhao, S.; Hulsegge, B.; Harders, F.L.; Bossers, R.; Keuning, E.; Hoekman, A.J.; Hoving-Bolink, R.; te Pas, M.F. Functional analysis of inter-individual transcriptome differential expression in pig longissimus muscle. J. Anim. Breed. Genet. 2013, 130, 72–78. [Google Scholar] [CrossRef]
- Rosenberg, H.; Pollock, N.; Schiemann, A.; Bulger, T.; Stowell, K. Malignant hyperthermia: A review. Orphanet J. Rare Dis. 2015, 10, 93. [Google Scholar] [CrossRef] [Green Version]
- Saini, B.L.; Gaur, G.K.; Sahoo, N.R.; Mendiratta, S.K.; Kumar, A.; Naha, B.C.; Baranwal, A.; Yadav, V.; Jaiswal, R.K. Polymorphism distribution of ryr1, prkag3, hfabp, myf-5 and mc4r genes in crossbred pigs. Mol. Biol. Rep. 2018, 45, 1575–1585. [Google Scholar] [CrossRef]
- Ghosh, M.; Sodhi, S.S.; Song, K.D.; Kim, J.H.; Mongre, R.K.; Sharma, N.; Singh, N.K.; Kim, S.W.; Lee, H.K.; Jeong, D.K. Evaluation of body growth and immunity-related differentially expressed genes through deep rna sequencing in the piglets of jeju native pig and berkshire. Anim. Genet. 2015, 46, 255–264. [Google Scholar] [CrossRef]
- Lim, K.S.; Lee, K.T.; Park, J.E.; Chung, W.H.; Jang, G.W.; Choi, B.H.; Hong, K.C.; Kim, T.H. Identification of differentially expressed genes in longissimus muscle of pigs with high and low intramuscular fat content using rna sequencing. Anim. Genet. 2017, 48, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.S.; Feugang, J.M.; Liao, S.F. A nutrigenomics approach using rna sequencing technology to study nutrient-gene interactions in agricultural animals. Curr. Dev. Nutr. 2019, 3, nzz082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ropka-Molik, K.; Zukowski, K.; Eckert, R.; Gurgul, A.; Piorkowska, K.; Oczkowicz, M. Comprehensive analysis of the whole transcriptomes from two different pig breeds using rna-seq method. Anim. Genet. 2014, 45, 674–684. [Google Scholar] [CrossRef] [PubMed]
- KAPE. Available online: https://www.ekape.or.kr/english/contents/list.do?menuId=menu156582&boardInfoNo= (accessed on 6 August 2019).
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of rna-seq experiments with tophat and cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, U.; Kim, B.; Yoon, J. A computational method for detecting copy number variations using scale-space filtering. BMC Bioinform. 2013, 14, 57. [Google Scholar] [CrossRef] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic rna-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Furumichi, M.; Tanabe, M.; Hirakawa, M. Kegg for representation and analysis of molecular networks involving diseases and drugs. Nucleic Acids Res. 2010, 38, D355–D360. [Google Scholar] [CrossRef] [Green Version]
- Hoshijima, M. Mechanical stress-strain sensors embedded in cardiac cytoskeleton: Z disk, titin, and associated structures. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1313–H1325. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Ye, F.; He, L.; Liu, Y.; Zhao, X.; Yin, H.; Li, D.; Xu, H.; Zhu, Q.; Wang, Y. Molecular cloning, expression profiling, and marker validation of the chicken myoz3 gene. Biomed. Res. Int. 2017, 2017, 5930918. [Google Scholar] [CrossRef]
- Kimura, A. Molecular genetics and pathogenesis of cardiomyopathy. J. Hum. Genet. 2016, 61, 41–50. [Google Scholar] [CrossRef]
- Batonnet-Pichon, S.; Behin, A.; Cabet, E.; Delort, F.; Vicart, P.; Lilienbaum, A. Myofibrillar myopathies: New perspectives from animal models to potential therapeutic approaches. J. Neuromuscul. Dis. 2017, 4, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterman-Storer, C.M. The cytoskeleton of skeletal muscle: Is it affected by exercise? A brief review. Med. Sci. Sports Exerc. 1991, 23, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Dransfield, E.; Sosnicki, A. Relationship between muscle growth and poultry meat quality. Poult. Sci. 1999, 78, 743–746. [Google Scholar] [CrossRef]
- Frey, N.; Olson, E.N. Calsarcin-3, a novel skeletal muscle-specific member of the calsarcin family, interacts with multiple z-disc proteins. J. Biol. Chem. 2002, 277, 13998–14004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Deng, C.Y.; Xiong, Y.Z.; Zuo, B. Molecular characterization, expression analysis and association study with meat quality traits of porcine ttid gene. Mol. Biol. Rep. 2013, 40, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Salas, R.C.; Mingala, C.N. Genetic factors affecting pork quality: Halothane and rendement napole genes. Anim. Biotechnol. 2017, 28, 148–155. [Google Scholar] [CrossRef]
- Diniz, W.J.S.; Mazzoni, G.; Coutinho, L.L.; Banerjee, P.; Geistlinger, L.; Cesar, A.S.M.; Bertolini, F.; Afonso, J.; de Oliveira, P.S.N.; Tizioto, P.C.; et al. Detection of co-expressed pathway modules associated with mineral concentration and meat quality in nelore cattle. Front. Genet. 2019, 10, 210. [Google Scholar] [CrossRef]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Velez-Irizarry, D.; Casiro, S.; Daza, K.R.; Bates, R.O.; Raney, N.E.; Steibel, J.P.; Ernst, C.W. Genetic control of longissimus dorsi muscle gene expression variation and joint analysis with phenotypic quantitative trait loci in pigs. BMC Genom. 2019, 20, 3. [Google Scholar] [CrossRef] [Green Version]
- Park, H.Y.; Lee, S.B.; Yoo, H.Y.; Kim, S.J.; Kim, W.S.; Kim, J.I.; Ko, Y.H. Whole-exome and transcriptome sequencing of refractory diffuse large b-cell lymphoma. Oncotarget 2016, 7, 86433–86445. [Google Scholar] [CrossRef] [Green Version]
- Karczewska-Kupczewska, M.; Stefanowicz, M.; Matulewicz, N.; Nikolajuk, A.; Straczkowski, M. Wnt signaling genes in adipose tissue and skeletal muscle of humans with different degrees of insulin sensitivity. J. Clin. Endocrinol. Metab. 2016, 101, 3079–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, J.C.; Zhang, Z.Z.; Wang, W.; McKinnie, S.M.K.; Vederas, J.C.; Oudit, G.Y. Targeting the apelin pathway as a novel therapeutic approach for cardiovascular diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Farkasfalvi, K.; Stagg, M.A.; Coppen, S.R.; Siedlecka, U.; Lee, J.; Soppa, G.K.; Marczin, N.; Szokodi, I.; Yacoub, M.H.; Terracciano, C.M. Direct effects of apelin on cardiomyocyte contractility and electrophysiology. Biochem. Biophys. Res. Commun. 2007, 357, 889–895. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Item | 1 + | 2 | Total | Statistical Test | |
|---|---|---|---|---|---|
| Carcass weight (kg) | 89.2 ± 2.35 | 94.00 ± 4.22 | 91.6 ± 4.13 | t = −3.16 ** | |
| Backfat thickness (mm) | 20.4 ± 2.17 | 27.2 ± 4.59 | 23.8 ± 4.94 | t = −4.24 *** | |
| pH | 5.66 ± 0.06 | 5.60 ± 0.04 | 5.61 ± 0.05 | t = 0.86 NS | |
| CIE color | L * | 55.06 ± 2.35 | 59.79 ± 1.77 | 57.42 ± 3.16 | t = −5.09 *** |
| a * | 9.24 ± 1.09 | 8.15 ± 0.53 | 8.69 ± 1.00 | t = 2.85 ** | |
| b * | 3.00 ± 0.66 | 3.85 ± 0.43 | 3.42 ± 0.70 | t = −3.42 ** | |
| Category | Term ID | Term Description | Observed Gene Count | Background Gene Count | FDR | Matching Genes * |
|---|---|---|---|---|---|---|
| Biological Process | GO:0008104 | Protein localization | 24 | 1966 | 0.0471 | ABRA, ADORA1, AP2A1, ATP6AP1, CAMK2G, DVL2, GRIP1, ITGAL, LIN7A, MSX1, NRXN1, NUBP1, PICK1, PITRM1, PKDCC, PSEN1, RAB3IP, RPS6, RRAGA, SNX13, UBE2D3, ULK1, VAMP8, VPS18 |
| GO:0030001 | Metal ion transport | 7 | 664 | 0.0034 | CACNA1S, CAMK2G, KCND2, KCNQ4, PSEN1, SCN4A, SLC9A4 | |
| GO:0016192 | Vesicle-mediated transport | 9 | 1699 | 0.0137 | ADORA1, ADORA2A, ALB, EEA1, HSP90AB1, LIN7A, MAPK3, PICK1, VAMP8 | |
| Molecular Function | GO:0005524 | ATP binding | 22 | 1462 | 0.0182 | AKT3, ALPK3, APAF1, ATP6AP1, CAMK2G, DYRK1B, EIF4A2, HSP90AB1, IP6K3, LIG1, MAPK3, MTOR, NLRP1, NUBP1, PKDCC, PRKCA, PSMC6, RECQL4, SMC6, SPEG, UBE2D3, ULK1 |
| GO:0005324 | Long-chain fatty acid transporter activity | 2 | 6 | 0.0324 | FABP3, FABP4 | |
| GO:0005504 | Fatty acid binding | 3 | 27 | 0.0324 | ALB, FABP3, FABP4 | |
| GO:0043168 | Anion binding | 31 | 2696 | 0.0324 | AKT3, ALB, ALPK3, APAF1, ATP6AP1, CAMK2G, DYRK1B, EEA1, EIF4A2, FABP3, FABP4, GOT2, GUF1, HSP90AB1, IP6K3, LIG1, MAPK3, MTOR, NLRP1, NUBP1, PKDCC, PRKCA, PSMC6, RECQL4, RRAGA, SMC6, SNX13, SPEG, UBE2D3, ULK1, UXS1 | |
| GO:0019901 | Protein kinase binding | 11 | 599 | 0.0414 | AP2A1, DUSP1, DVL2, HSP90AB1, MTOR, NR3C1, PCNA, PICK1, PRMT1, PTPRJ, RPS6 | |
| GO:0004674 | Protein serine/threonine kinase activity | 7 | 444 | 0.000000000313 | AKT3, CAMK2G, DYRK1B, MAPK3, MTOR, PRKCA, ULK1 | |
| Cellular Component | GO:0030018 | Z disc | 5 | 122 | 0.0153 | ITGB1BP2, MYOT, MYOZ3, PPP2R5A, PSEN1 |
| GO:0034703 | Cation channel complex | 3 | 206 | 0.0394 | CACNA1S, KCNQ4, SCN4A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-M.; Markkandan, K.; Lee, J.-Y.; Kim, G.-W.; Yoo, J.Y. Transcriptome Profiling Associated with Carcass Quality of Loin Muscles in Crossbred Pigs. Animals 2020, 10, 1279. https://doi.org/10.3390/ani10081279
Kim S-M, Markkandan K, Lee J-Y, Kim G-W, Yoo JY. Transcriptome Profiling Associated with Carcass Quality of Loin Muscles in Crossbred Pigs. Animals. 2020; 10(8):1279. https://doi.org/10.3390/ani10081279
Chicago/Turabian StyleKim, Sang-Mo, Kesavan Markkandan, Jong-Young Lee, Gye-Woong Kim, and Jae Young Yoo. 2020. "Transcriptome Profiling Associated with Carcass Quality of Loin Muscles in Crossbred Pigs" Animals 10, no. 8: 1279. https://doi.org/10.3390/ani10081279
APA StyleKim, S. -M., Markkandan, K., Lee, J. -Y., Kim, G. -W., & Yoo, J. Y. (2020). Transcriptome Profiling Associated with Carcass Quality of Loin Muscles in Crossbred Pigs. Animals, 10(8), 1279. https://doi.org/10.3390/ani10081279