Brisket Disease Is Associated with Lower Volatile Fatty Acid Production and Altered Rumen Microbiome in Holstein Heifers

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Design and Sample Collection
2.3. DNA Extraction, 16S rRNA Gene Amplicon Preparation and Sequencing
2.4. Bioinformatics and Statistical Analyses
3. Results
3.1. Mean Pulmonary Arterial Pressure, Blood Oxygen Saturation Breathing Rate and Rumen VFAs of the BD and Healthy Heifer Group (HH) Animals
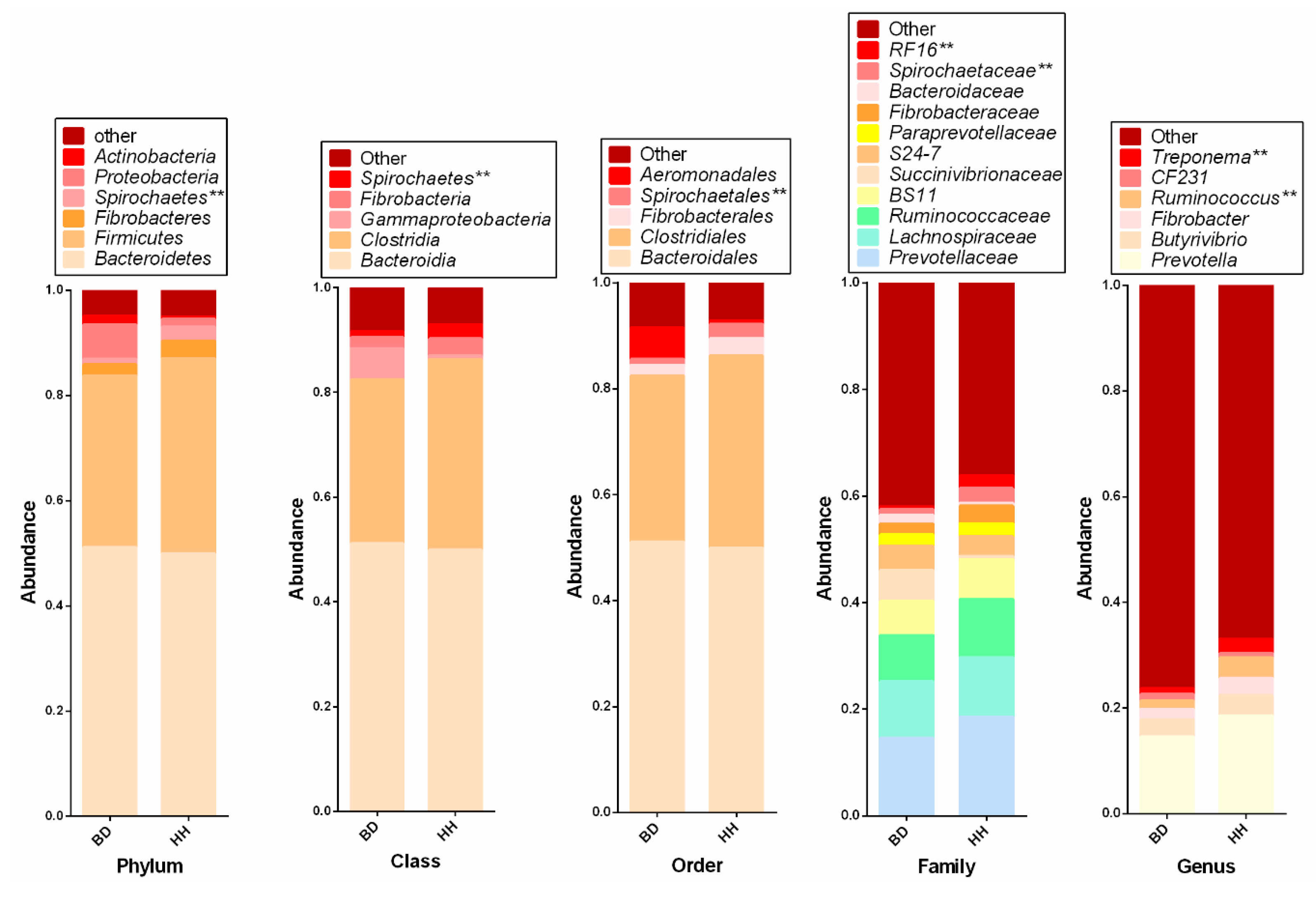
3.2. Changes of Rumen Predominant Microbiota
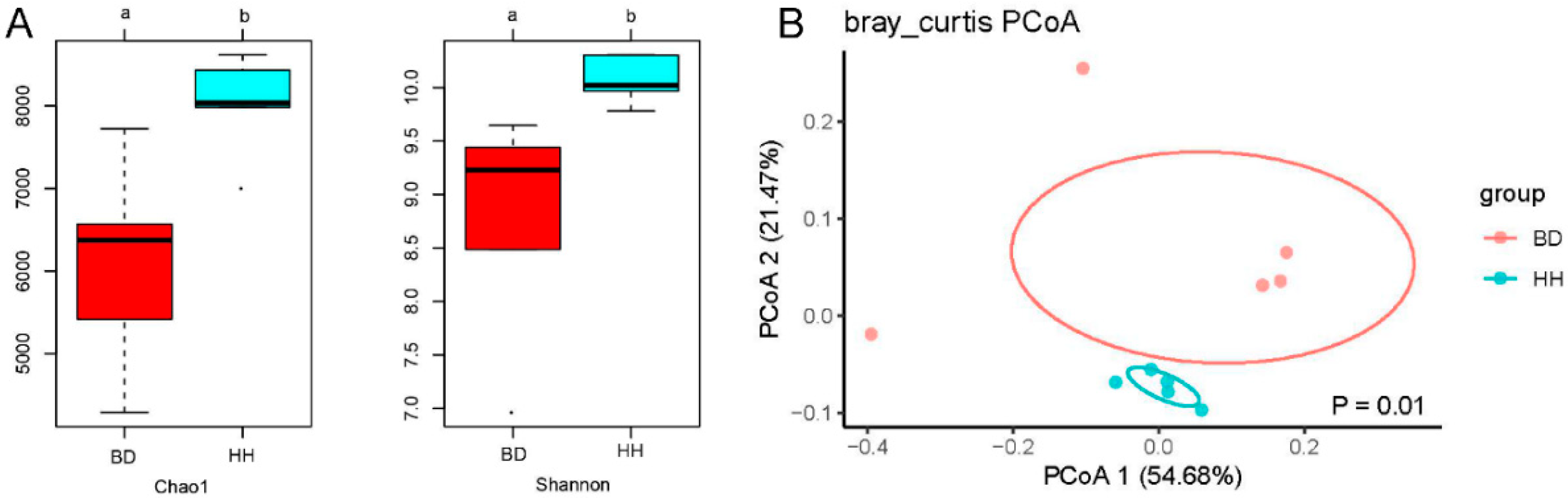
3.3. Alpha and Beta Diversity of the Rumen Microbial Community
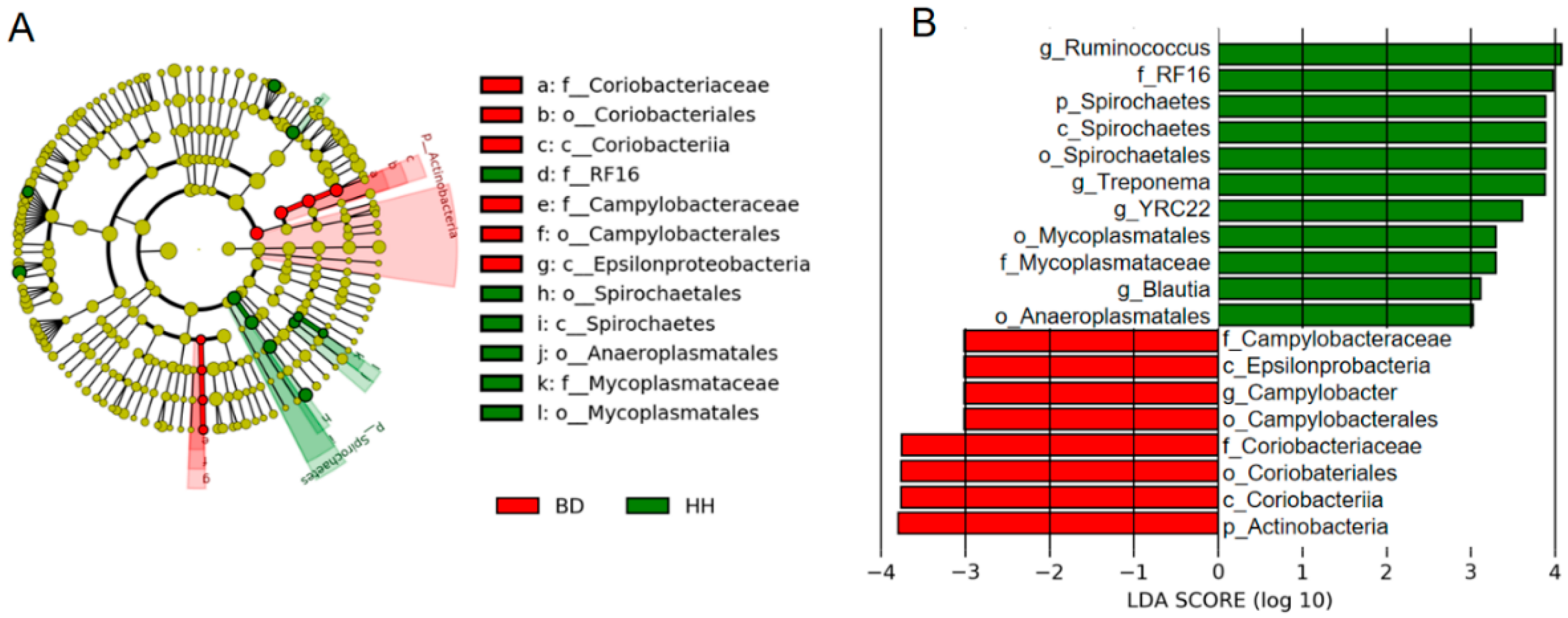
3.4. Microbial Species with Significant Differences
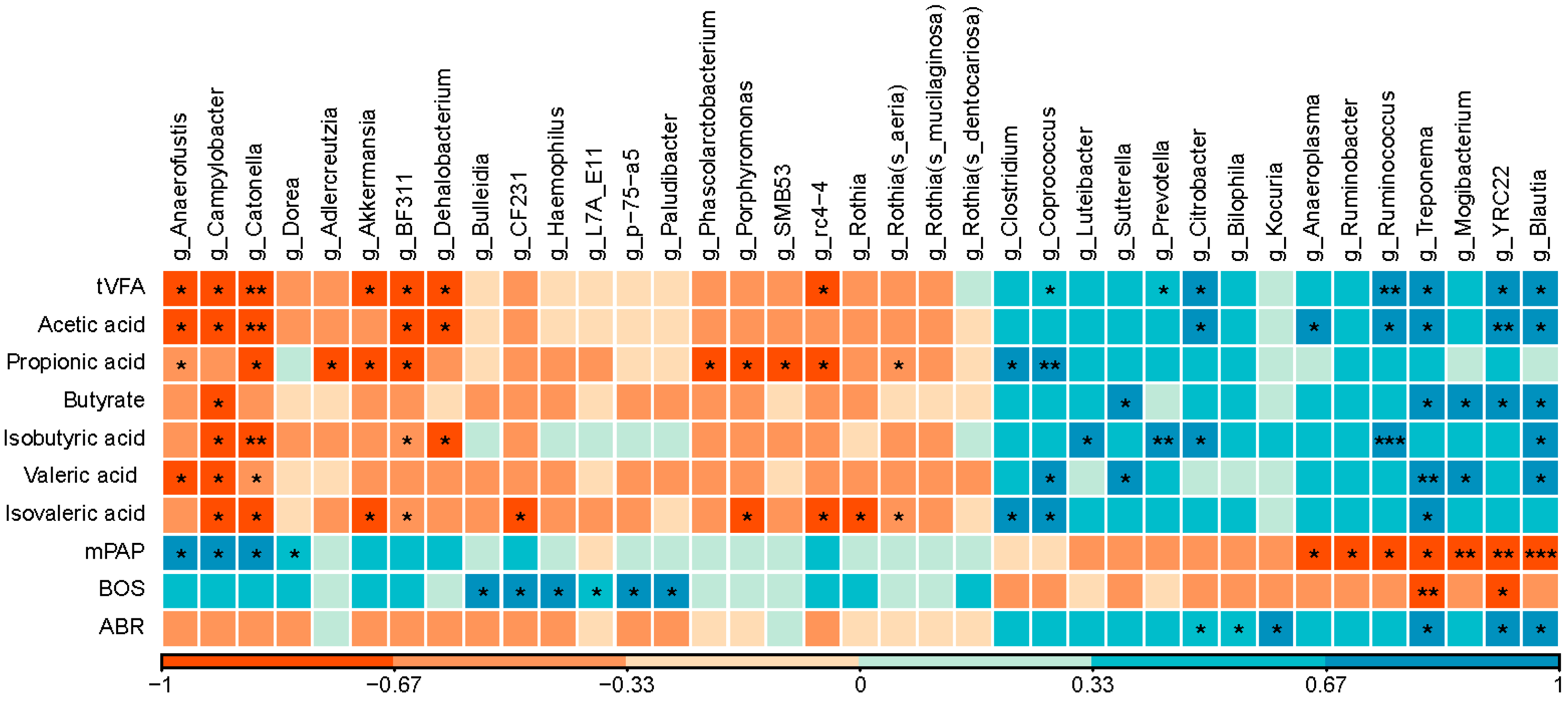
3.5. Correlation between Genera and Physiological Indicators of Heifers
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content |
|---|---|
| Ingredient/diet (g/100 g DM) | |
| Corn silage | 36.40 |
| Alfalfa hay | 24.36 |
| Oat hay | 9.56 |
| Corn grain | 14.91 |
| Soybean meal | 4.47 |
| Rapeseed meal | 2.09 |
| Cotton seed meal | 1.79 |
| Wheat bran | 1.82 |
| Cottonseed protein | 1.49 |
| Jujube powder | 1.04 |
| Premix | 0.30 |
| Limestone | 0.50 |
| Dicalcium phosphate | 0.42 |
| Sodium bicarbonate | 0.75 |
| Salt | 0.24 |
| Chemical composition (g/100 g DM) | |
| Crude protein | 13.49 |
| NEL (MJ/kg) | 6.02 |
| Neutral detergent fiber | 34.11 |
| Acid detergent fiber | 22.96 |
| Ether extract | 3.50 |
| Ash | 6.40 |
| Calcium | 0.73 |
| Total phosphorus | 0.47 |
References
- Pretty, J.N. The sustainable intensification of agriculture. Nat. Resour. Forum 1997, 21, 247–256. [Google Scholar] [CrossRef]
- Hirschler, V. Cardiometabolic risk factors in native populations living at high altitudes. Int. J. Clin. Pract. 2016, 70, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, G.; Paul, V.; Hanah, S.; Bam, J.; Das, P. Effects of climate change on yak production at high altitude. Indian J. Anim. Sci. 2016, 86, 621–626. [Google Scholar]
- Holt, T.N.; Callan, R.J. Pulmonary arterial pressure testing for high mountain disease in cattle. Vet. Clin. N. Am. 2007, 23, 575–596. [Google Scholar] [CrossRef] [PubMed]
- Malherbe, C.R.; Marquard, J.; Legg, D.E.; Cammack, K.M.; O’Toole, D. Right ventricular hypertrophy with heart failure in Holstein heifers at elevation of 1,600 meters. J. Vet. Diagn. Investig. 2012, 24, 867–877. [Google Scholar] [CrossRef] [Green Version]
- Heinonen, I.; Luotolahti, M.; Vuolteenaho, O.; Nikinmaa, M.; Saraste, A.; Hartiala, J.; Koskenvuo, J.; Knuuti, J.; Arjamaa, O. Circulating N-terminal brain natriuretic peptide and cardiac function in response to acute systemic hypoxia in healthy humans. J. Transl. Med. 2014, 12, 189. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Azarfar, A.; Wang, Y.; Cao, Z.; Li, S. N-carbamylglutamate restores nitric oxide synthesis and attenuates high altitude-induced pulmonary hypertension in Holstein heifers ascended to high altitude. J. Anim. Sci. Biotechnol. 2018, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.B.; Rychlik, J.L. Factors that alter rumen microbial ecology. Science 2001, 292, 1119–1122. [Google Scholar] [CrossRef]
- Yohe, T.; Schramm, H.; White, R.; Hanigan, M.; Parsons, C.; Tucker, H.; Enger, B.; Hardy, N.; Daniels, K. Form of calf diet and the rumen. II: Impact on volatile fatty acid absorption. J. Dairy Sci. 2019, 102, 8502–8512. [Google Scholar] [CrossRef]
- Bond, J.; Donaldson, A.; Coumans, J.; Austin, K.; Ebert, D.; Wheeler, D.; Oddy, V. Protein profiles of enzymatically isolated rumen epithelium in sheep fed a fibrous diet. J. Anim. Sci. Biotechnol. 2019, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Roh, S.; Kuno, M.; Hishikawa, D.; Hong, Y.; Katoh, K.; Obara, Y.; Hidari, H.; Sasaki, S. Identification of differentially expressed transcripts in bovine rumen and abomasum using a differential display method. J. Anim. Sci. 2007, 85, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Shabat, S.K.; Sasson, G.; Doron-Faigenboim, A.; Durman, T.; Yaacoby, S.; Berg Miller, M.E.; White, B.A.; Shterzer, N.; Mizrahi, I. Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants. ISME J. 2016, 10, 2958–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudel, P.; Froehlich, K.; Casper, D.P.; St-Pierre, B. Feeding Essential Oils to Neonatal Holstein Dairy Calves Results in Increased Ruminal Prevotellaceae Abundance and Propionate Concentrations. Microorganisms 2019, 7, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, J.J.; Zhang, H.; Yang, D.L.; Zhang, Y.H.; Xiong, B.H.; Jiang, L.S. Illumina sequencing analysis of the ruminal microbiota in high-yield and low-yield lactating dairy cows. PLoS ONE 2018, 13, e0198225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishaq, S.L.; Yeoman, C.J.; Whitney, T.R. Ground Juniperus pinchotii and urea in supplements fed to Rambouillet ewe lambs Part 2: Ewe lamb rumen microbial communities. J. Anim. Sci. 2017, 95, 4587–4599. [Google Scholar] [CrossRef]
- Vlková, E.; Trojanová, I.; Rada, V. Distribution of bifidobacteria in the gastrointestinal tract of calves. Folia Microbiol. 2006, 51, 325–328. [Google Scholar] [CrossRef]
- Qiao, G.H.; Yu, C.Q.; Li, J.H.; Yang, X.; Zhu, X.Q.; Zhou, X.H. Effect of high altitude on nutrient digestibility, rumen fermentation and basal metabolism rate in Chinese Holstein cows on the Tibetan plateau. Anim. Prod. Sci. 2013, 53, 240–246. [Google Scholar] [CrossRef]
- Zhang, F.; Yang, W.; Deng, Z.; Wu, W.; Wu, H.; Chen, J.; Wang, Y.; Yang, Y. Effect of glutamine on change of intestinal microecology in rats exposed to acute high altitude. Chin. J. Microbiol. 2010, 22, 1–4. [Google Scholar]
- Michaux, H.; Nichols, S.; Babkine, M.; Francoz, D. Description of thoracoscopy and associated short-term cardiovascular and pulmonary effects in healthy cattle. Am. J. Vet. Res. 2014, 75, 468–476. [Google Scholar] [CrossRef]
- Cao, Z.; Li, S.; Xing, J.; Ma, M.; Wang, L. Effects of maize grain and lucerne particle size on ruminal fermentation, digestibility and performance of cows in midlactation. J. Anim. Physiol. Anim. Nutr. 2008, 92, 157–167. [Google Scholar] [CrossRef]
- Sun, W.; Qian, X.; Gu, J.; Wang, X.J.; Zhang, L.; Guo, A.Y. Mechanisms and effects of arsanilic acid on antibiotic resistance genes and microbial communities during pig manure digestion. Bioresour. Technol. 2017, 234, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, D.; Chen, H.; Luo, X.; Guan, J.; He, Y.; Zhao, X. Microbial diversity in the rumen, reticulum, omasum, and abomasum of yak on a rapid fattening regime in an agro-pastoral transition zone. J. Microbiol. 2018, 56, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.Q.; Yang, D.Z.; Luo, Y.J.; Li, S.Z.; Liu, F.Y.; Wang, G.S. Over-starvation aggravates intestinal injury and promotes bacterial and endotoxin translocation under high-altitude hypoxic environment. World J. Gastroenterol. 2011, 17, 1584–1593. [Google Scholar] [CrossRef]
- Gozho, G.; Mutsvangwa, T. Influence of carbohydrate source on ruminal fermentation characteristics, performance, and microbial protein synthesis in dairy cows. J. Dairy Sci. 2008, 91, 2726–2735. [Google Scholar] [CrossRef]
- Lei, Y.; Zhang, K.; Guo, M.; Li, G.; Li, C.; Li, B.; Yang, Y.; Chen, Y.; Wang, X. Exploring the spatial-temporal microbiota of compound stomachs in a pre-weaned goat model. Front. Microbiol. 2018, 9, 1846. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Shi, H.; Wang, Y.; Li, S.; Cao, Z.; Ji, S.; He, Y.; Zhang, H. Effect of dietary forage to concentrate ratios on dynamic profile changes and interactions of ruminal microbiota and metabolites in holstein heifers. Front. Microbiol. 2017, 8, 2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, R.; Zou, H.; Wang, Z.; Cao, B.; Peng, Q.; Jing, X.; Wang, Y.; Shao, Y.; Pei, Z.; Zhang, X. Nutritional interventions improved rumen functions and promoted compensatory growth of growth-retarded yaks as revealed by integrated transcripts and microbiome analyses. Front. Microbiol. 2019, 10, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census, C.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [PubMed]
- Rosewarne, C.P.; Cheung, J.L.; Smith, W.J.; Evans, P.N.; Tomkins, N.W.; Denman, S.E.; Cuív, P.Ó.; Morrison, M. Draft genome sequence of Treponema sp. strain JC4, a novel spirochete isolated from the bovine rumen. J. Bacteriol. 2012, 194, 4130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.J.; Zhang, G.N.; Xu, H.J.; Xin, H.S.; Zhang, Y.G. Metagenomic analyses of microbial and carbohydrate-active enzymes in the rumen of holstein cows fed different forage-to-concentrate ratios. Front. Microbiol. 2019, 10, 649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.; Seo, J.; Choi, H.; Yoon, D.; Nam, J.; Kim, H.; Cho, S.; Chang, J. Metagenome analysis of protein domain collocation within cellulase genes of goat rumen microbes. Asian-Australas. J. Anim. Sci. 2013, 26, 1144–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kala, A.; Kamra, D.; Kumar, A.; Agarwal, N.; Chaudhary, L.; Joshi, C. Impact of levels of total digestible nutrients on microbiome, enzyme profile and degradation of feeds in buffalo rumen. PLoS ONE 2017, 12, e0172051. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Sun, R.; Qiao, X.; Xu, C.; Shang, X.; Niu, W. Protective effect of glutamine on intestinal injury and bacterial community in rats exposed to hypobaric hypoxia environment. World J. Gastroenterol. 2014, 20, 4662–4674. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jiang, X.; Yang, C.; Ma, B.; Lei, C.; Xu, C.; Zhang, A.; Yang, X.; Xiong, Q.; Zhang, P. Cecal microbiota of Tibetan Chickens from five geographic regions were determined by 16S rRNA sequencing. Microbiology 2016, 5, 753–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Cilleros, D.; Ramos, S.; Lopez-Oliva, M.E.; Escriva, F.; Alvarez, C.; Fernandez-Millan, E.; Martin, M.A. Cocoa diet modulates gut microbiota composition and improves intestinal health in Zucker diabetic rats. Food Res. Int. 2020, 132, 109058. [Google Scholar] [CrossRef]
- Moreno-Pérez, D.; Bressa, C.; Bailén, M.; Hamed-Bousdar, S.; Naclerio, F.; Carmona, M.; Pérez, M.; González-Soltero, R.; Montalvo-Lominchar, M.G.; Carabaña, C. Effect of a protein supplement on the gut microbiota of endurance athletes: A randomized, controlled, double-blind pilot study. Nutrients 2018, 10, 337. [Google Scholar] [CrossRef] [Green Version]
- Barcenilla, A.; Pryde, S.E.; Martin, J.C.; Duncan, S.H.; Stewart, C.S.; Henderson, C.; Flint, H.J. Phylogenetic relationships of butyrate-producing bacteria from the human gut. Appl. Environ. Microbiol. 2000, 66, 1654–1661. [Google Scholar] [CrossRef] [Green Version]
- Khanna, K.; Mishra, K.P.; Ganju, L.; Kumar, B.; Singh, S.B. High-altitude-induced alterations in gut-immune axis: A review. Int. Rev. Immunol. 2018, 37, 119–126. [Google Scholar] [CrossRef]
- Yang, B.; Le, J.Q.; Wu, P.; Liu, J.X.; Guan, L.; Wang, J.K. Alfalfa intervention alters rumen microbial community development in Hu lambs during early life. Front. Microbiol. 2018, 9, 574. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Guo, W.S.; Meng, Q.X.; Stevenson, D.M.; Weimer, P.J.; Schaefer, D.M. Changes in rumen bacterial community composition in steers in response to dietary nitrate. Appl. Microbiol. Biotechnol. 2013, 97, 8719–8727. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.P.; Meng, Q.X.; Ren, L.P.; Liu, W.; Zhang, X.Z.; Huo, Y.L.; Zhou, Z.M. Effects of nitrate addition on rumen fermentation, bacterial biodiversity and abundance. Asian-Australas. J. Anim. Sci. 2015, 28, 1433–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyström, T.; Ortsäter, H.; Huang, Z.; Zhang, F.; Larsen, F.J.; Weitzberg, E.; Lundberg, J.O.; Sjöholm, Å. Inorganic nitrite stimulates pancreatic islet blood flow and insulin secretion. Free Radic. Biol. Med. 2012, 53, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, M.; Ungerfeld, E.M.; Zhang, X.M.; Long, D.L.; Mao, H.X.; Deng, J.P.; Bannink, A.; Tan, Z.L. Nitrate improves ammonia incorporation into rumen microbial protein in lactating dairy cows fed a low-protein diet. J. Dairy Sci. 2018, 101, 9789–9799. [Google Scholar] [CrossRef] [Green Version]
- Fagan, K.A.; Morrissey, B.; Fouty, B.W.; Sato, K.; Harral, J.W.; Morris, K.G.; Hoedt-Miller, M.; Vidmar, S.; McMurtry, I.F.; Rodman, D.M. Upregulation of nitric oxide synthase in mice with severe hypoxia-induced pulmonary hypertension. Respir. Res. 2001, 2, 306–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonelli, A.R.; Haserodt, S.; Aytekin, M.; Dweik, R.A. Nitric oxide deficiency in pulmonary hypertension: Pathobiology and implications for therapy. Pulm. Circ. 2013, 3, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Finegold, S.M.; Lawson, P.A.; Vaisanen, M.L.; Molitoris, D.R.; Song, Y.; Liu, C.; Collins, M.D. Anaerofustis stercorihominis gen. nov., sp. nov., from human feces. Anaerobe 2004, 10, 41–45. [Google Scholar] [CrossRef]
- Zhang, K.; Fujita, Y.; Chang, L.J.; Qui, Y.G.; Pu, Y.Y.; Wang, S.M.; Shirayama, Y.; Hashimoto, K. Abnormal composition of gut microbiota is associated with resilience versus susceptibility to inescapable electric stress. Transl. Psychiatry 2019, 9, 231. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, M.; Planells, P.; Martinez, E.; Mira, A.; Carda-Dieguez, M. Microbiology of molar-incisor hypomineralization lesions. A pilot study. J. Oral Microbiol. 2020, 12, 1766166. [Google Scholar] [CrossRef]
- Lu, H.F.; Ren, Z.G.; Li, A.; Li, J.Y.; Xu, S.Y.; Zhang, H.; Jiang, J.W.; Yang, J.Z.; Luo, Q.X.; Zhou, K.; et al. Tongue coating microbiome data distinguish patients with pancreatic head cancer from healthy controls. J. Oral Microbiol. 2019, 11, 1563409. [Google Scholar] [CrossRef]
- Shirley, K.; Beckman, D.; Garrick, D. Inheritance of pulmonary arterial pressure in Angus cattle and its correlation with growth. J. Anim. Sci. 2008, 86, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Anand, I.; Harris, E.; Ferrari, R.; Pearce, P.; Harris, P. Pulmonary haemodynamics of the Yak, cattle, and cross breeds at high altitude. Thorax 1986, 41, 696–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]




| Items | HH | BD | p-Value |
|---|---|---|---|
| mPAP (mmHg) | 38.63 ± 1.56 | 74.73 ± 9.41 | 0.0018 |
| Blood oxygen saturation (%) | 88.33 ± 4.13 | 76.4 ± 4.75 | 0.0345 |
| Average breathing rate (breaths/minute) | 33.67 ± 6.08 | 17.2 ± 3.39 | 0.0114 |
| Items | HH | BD | p-Value |
|---|---|---|---|
| Total VFA (mmol/L) | 120.8 ± 4.84 | 77.30 ± 6.23 | 0.0006 |
| Acetic acid (%) | 66.46 ± 0.6839 | 63.19 ± 1.450 | 0.0758 |
| Propionic acid (%) | 17.46 ± 0.4576 | 19.04 ± 2.372 | 0.5303 |
| Butyric acid (%) | 11.22 ± 0.5580 | 10.97 ± 1.141 | 0.8458 |
| Isobutyric acid (%) | 1.483 ± 0.0509 | 1.560 ± 0.1287 | 0.5938 |
| Valeric acid (%) | 1.646 ± 0.0544 | 1.602 ± 0.1484 | 0.7891 |
| Isovaleric acid (%) | 1.720 ± 0.1434 | 1.943 ± 0.1151 | 0.2600 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaowa, N.; Panke-Buisse, K.; Wang, S.; Wang, H.; Cao, Z.; Wang, Y.; Yao, K.; Li, S. Brisket Disease Is Associated with Lower Volatile Fatty Acid Production and Altered Rumen Microbiome in Holstein Heifers. Animals 2020, 10, 1712. https://doi.org/10.3390/ani10091712
Gaowa N, Panke-Buisse K, Wang S, Wang H, Cao Z, Wang Y, Yao K, Li S. Brisket Disease Is Associated with Lower Volatile Fatty Acid Production and Altered Rumen Microbiome in Holstein Heifers. Animals. 2020; 10(9):1712. https://doi.org/10.3390/ani10091712
Chicago/Turabian StyleGaowa, Naren, Kevin Panke-Buisse, Shuxiang Wang, Haibo Wang, Zhijun Cao, Yajing Wang, Kun Yao, and Shengli Li. 2020. "Brisket Disease Is Associated with Lower Volatile Fatty Acid Production and Altered Rumen Microbiome in Holstein Heifers" Animals 10, no. 9: 1712. https://doi.org/10.3390/ani10091712
APA StyleGaowa, N., Panke-Buisse, K., Wang, S., Wang, H., Cao, Z., Wang, Y., Yao, K., & Li, S. (2020). Brisket Disease Is Associated with Lower Volatile Fatty Acid Production and Altered Rumen Microbiome in Holstein Heifers. Animals, 10(9), 1712. https://doi.org/10.3390/ani10091712