Multidrug Antimicrobial Resistance and Molecular Detection of mcr-1 Gene in Salmonella Species Isolated from Chicken

 ,
,  ,
,  ,
,  , , , , , , ,
, , , , , , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Standards
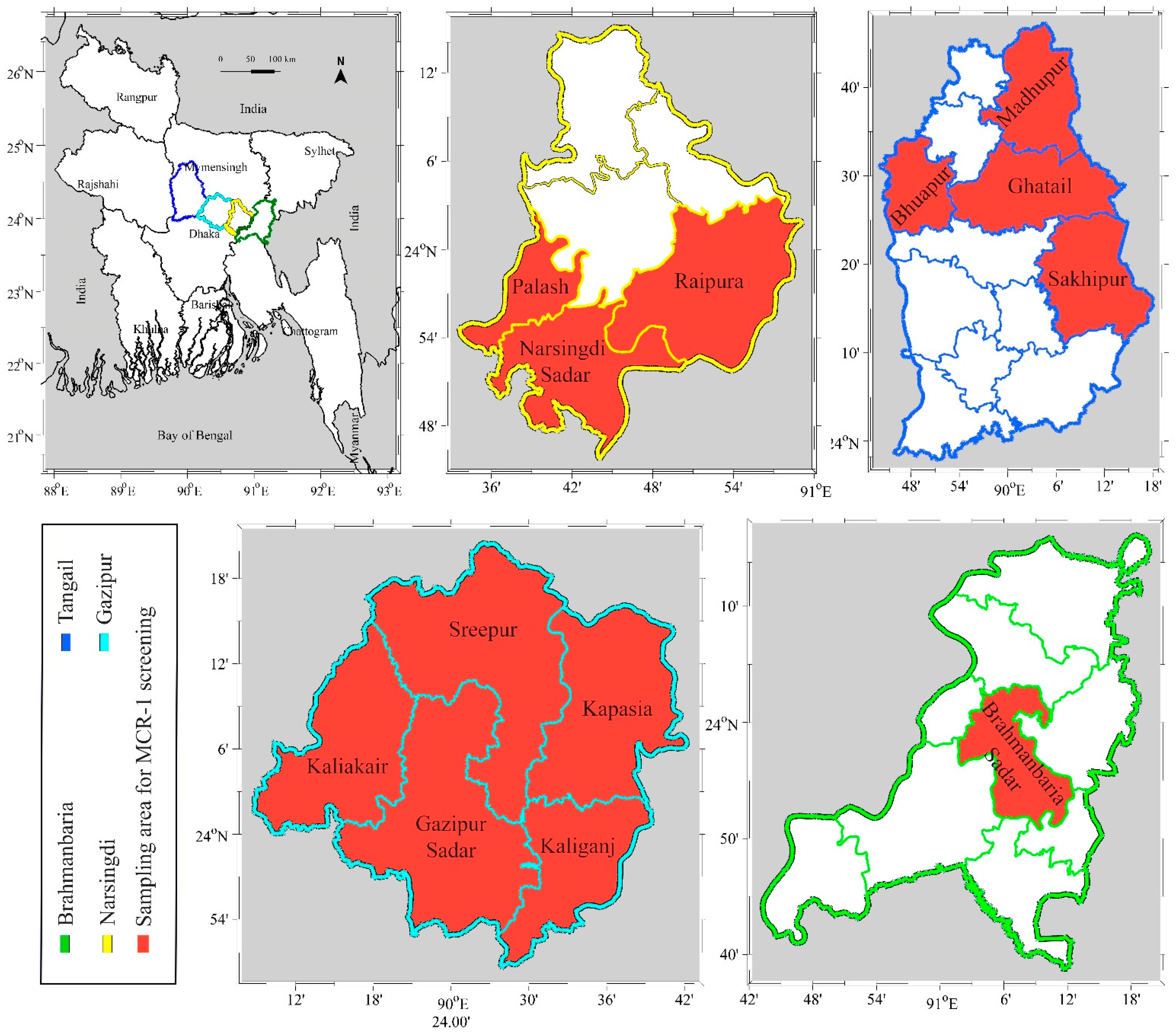
2.2. Study Area and Sampling
2.3. Isolation and Biochemical Identification of Bacterial Isolates
2.4. Antimicrobial Susceptibility Testing
2.5. Bacterial DNA Extraction
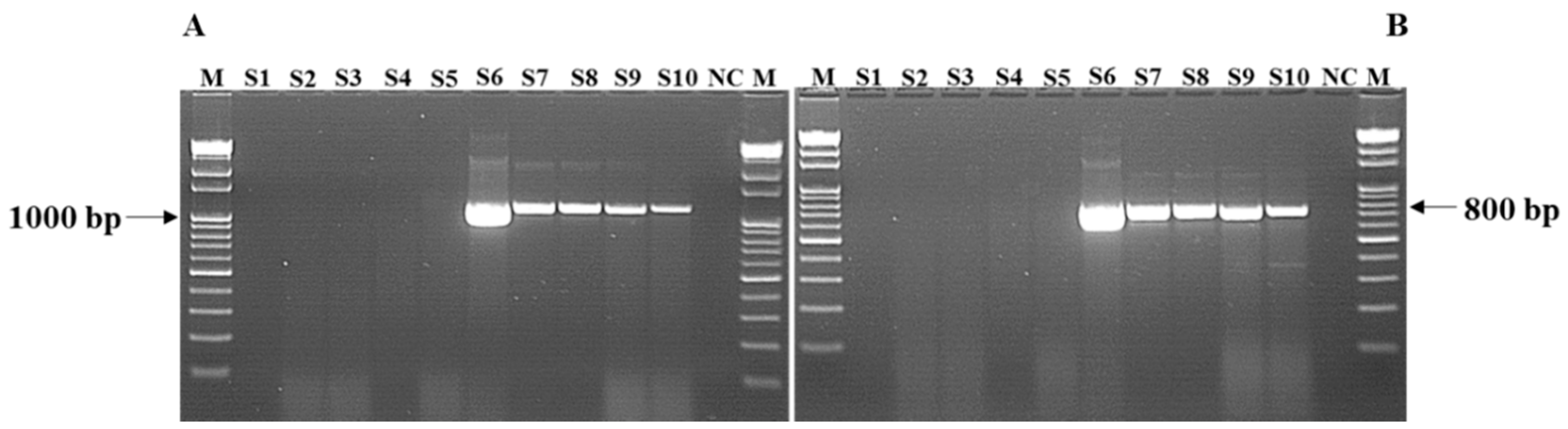
2.6. Polymerase Chain Reaction (PCR) and Gel Electrophoresis
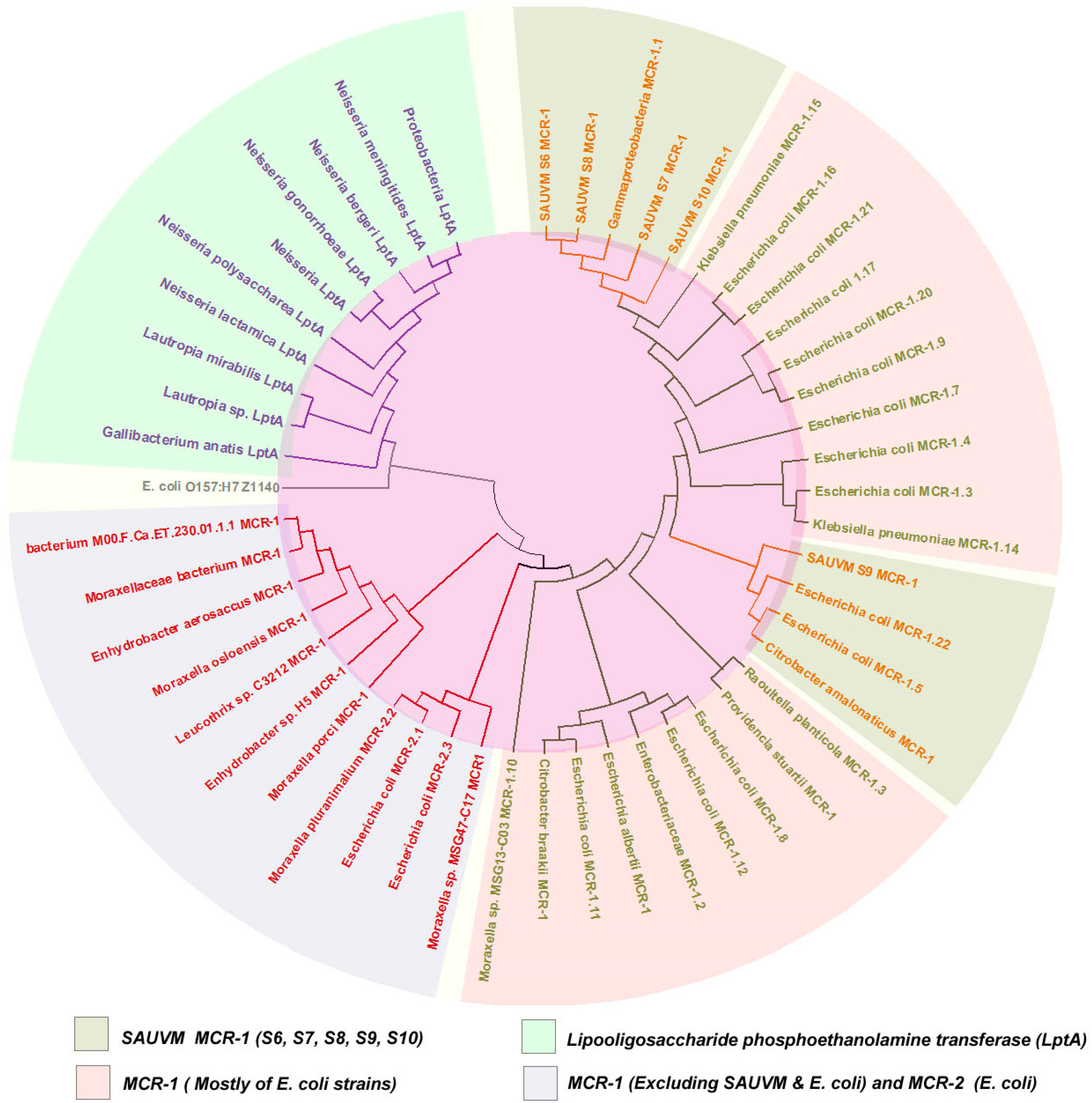
2.7. Sequencing, Multiple Sequence Alignment and Phylogenetic Analysis
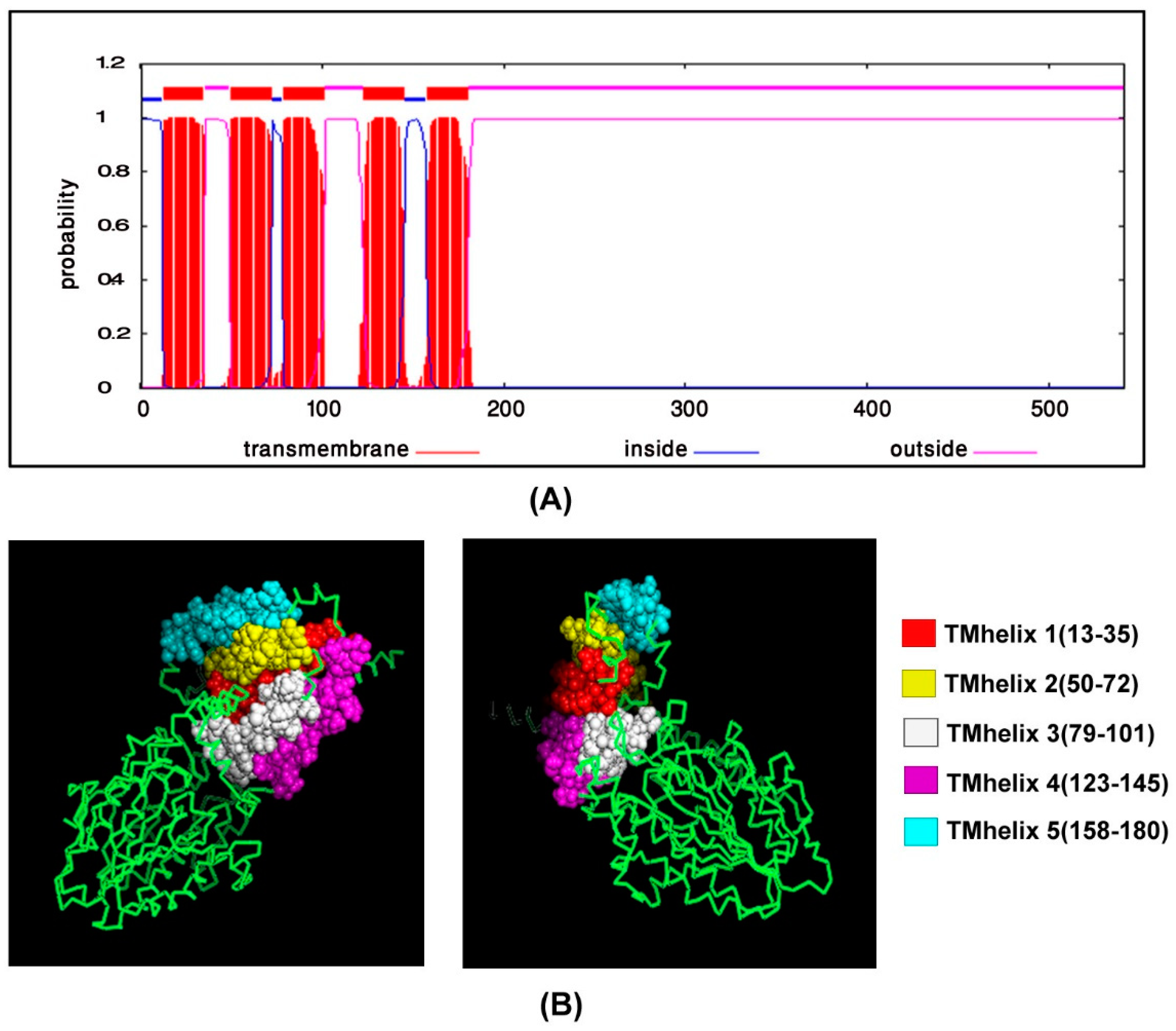
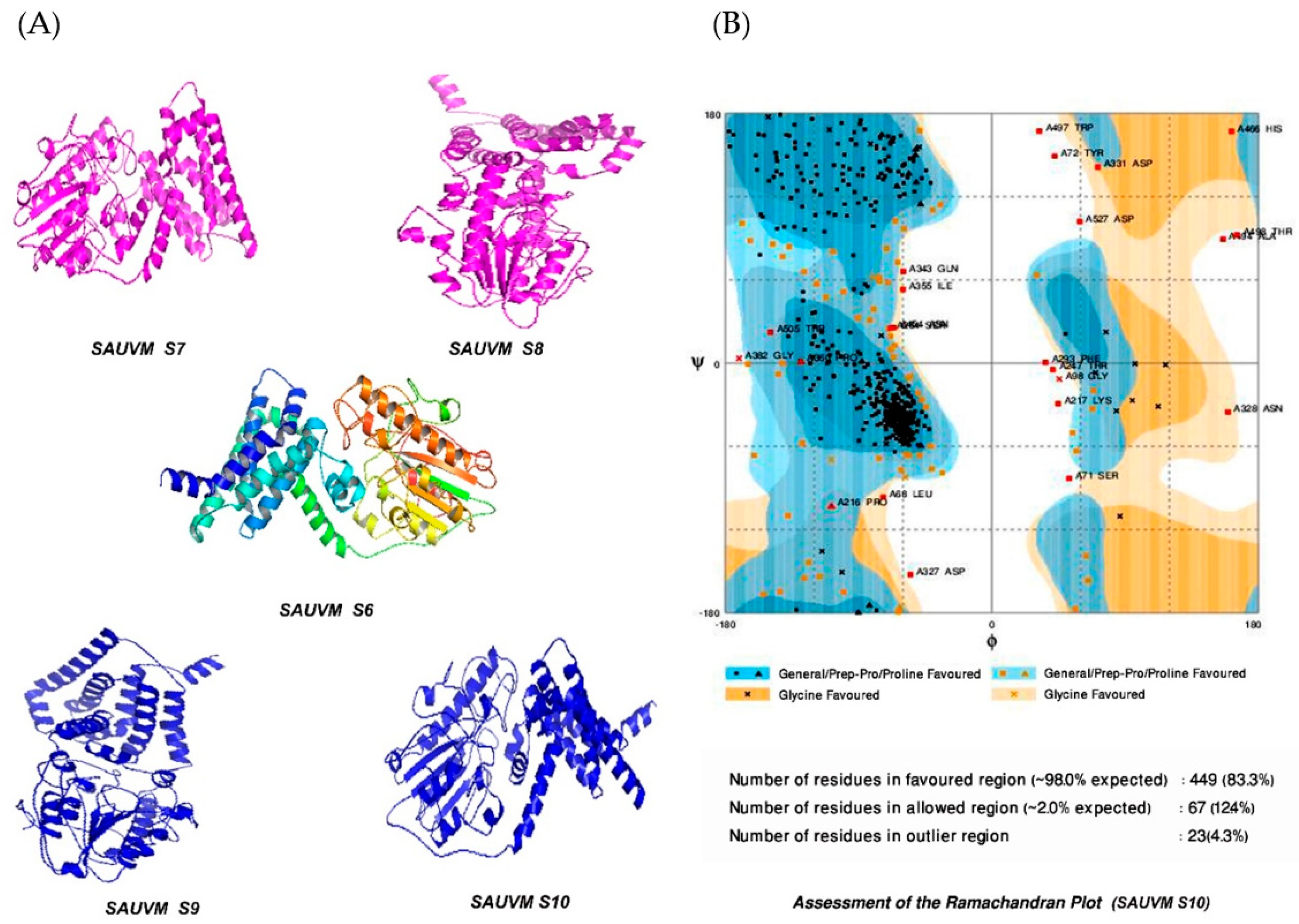
2.8. Transmembrane Topology Analysis, Structural Modelling, Refinement and Validation
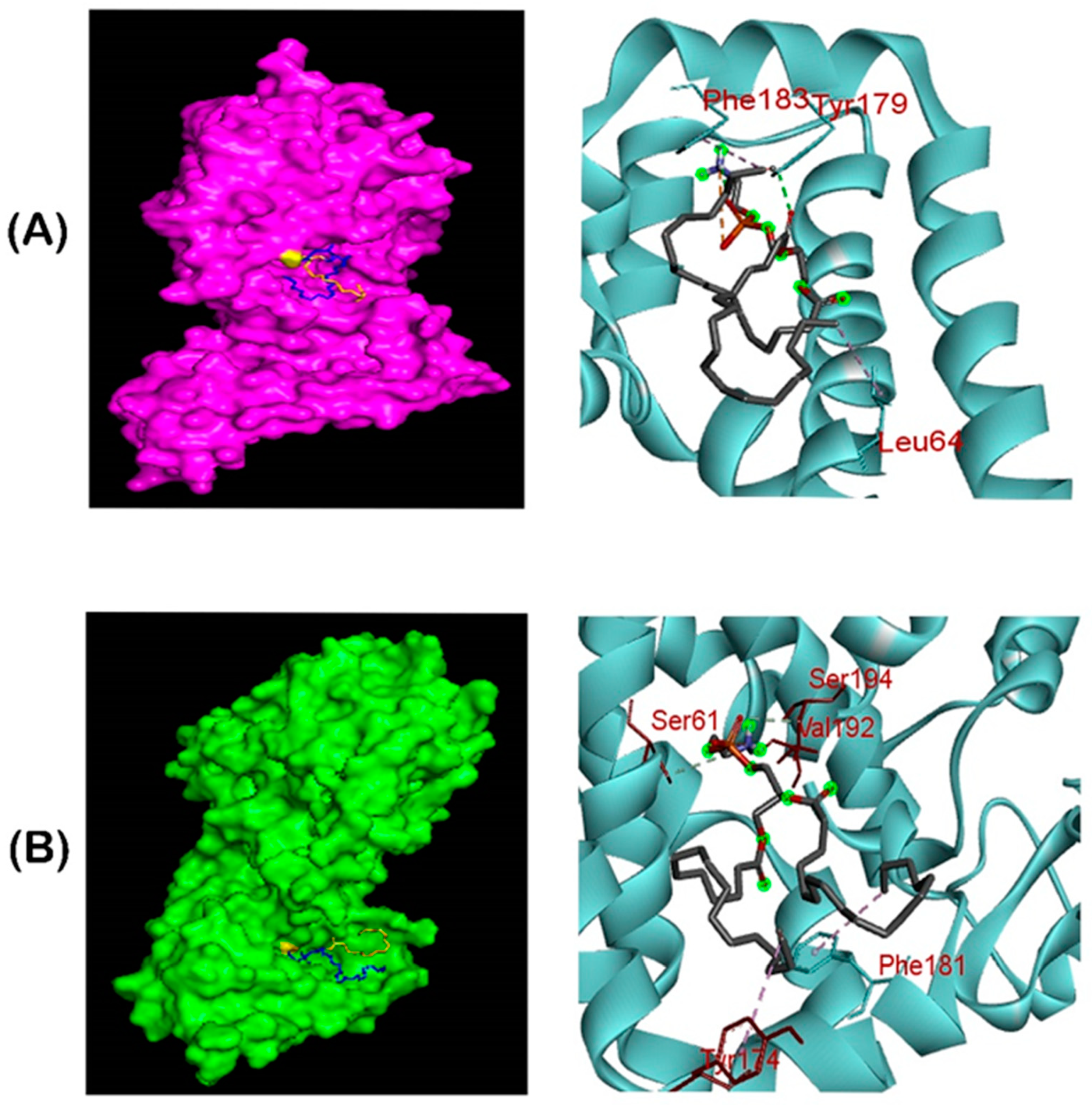
2.9. Molecular Docking of Phosphatidylethanolamine Substrate with mcr-1 and LptA
3. Results
3.1. Confirmation of Salmonella spp.
3.2. Antimicrobial Susceptibility Testing
3.3. Detection of Salmonella Isolates Using PCR and Sequencing of mcr-1 Genes
3.4. Sequence Acquisition, Multiple Sequence Alignment and Phylogenetic Analysis
3.5. Transmembrane Topology Analysis, Structural Modelling, Refinement and Validation
3.6. Molecular Docking of PE Substrate with MCR-1 and LptA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Hamid, M.A.; Rahman, M.A.; Ahmed, S.; Hossain, K.M. Status of Poultry Industry in Bangladesh and the Role of Private Sector for its Development. Asian J. Poult. Sci. 2016, 11, 1–13. [Google Scholar] [CrossRef]
- El-Sharkawy, H.; Tahoun, A.; El-Gohary, A.E.G.A.; El-Abasy, M.; El-Khayat, F.; Gillespie, T.; Kitade, Y.; Hafez, H.M.; Neubauer, H.; El-Adawy, H. Epidemiological, molecular characterization and antibiotic resistance of Salmonella enterica serovars isolated from chicken farms in Egypt. Gut Pathog. 2017, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosby, D.E.; Cox, N.A.; Harrison, M.A.; Wilson, J.L.; Jeff Buhr, R.; Fedorka-Cray, P.J. Salmonella and antimicrobial resistance in broilers: A review. J. Appl. Poult. Res. 2015, 24, 408–426. [Google Scholar] [CrossRef]
- Hoque, M.; Mohiuddin, R.; Khan, M.; Hannan, M.; Alam, M. Outbreak of Salmonella in Poultry of Bangladesh and possible remedy. J. Adv. Biotechnol. Exp. Ther. 2019, 2, 87. [Google Scholar] [CrossRef]
- Kumar, H.; Chen, B.H.; Kuca, K.; Nepovimova, E.; Kaushal, A.; Nagraik, R.; Bhatia, S.K.; Dhanjal, D.S.; Kumar, V.; Kumar, A.; et al. Understanding of colistin usage in food animals and available detection techniques: A review. Animals 2020, 10, 1892. [Google Scholar] [CrossRef]
- Davies, M.; Walsh, T.R. A colistin crisis in India. Lancet. Infect. Dis. 2018, 18, 256–257. [Google Scholar] [CrossRef]
- Al Masud, A.; Rousham, E.K.; Islam, M.A.; Alam, M.U.; Rahman, M.; Al Mamun, A.; Sarker, S.; Asaduzzaman, M.; Unicomb, L. Drivers of Antibiotic Use in Poultry Production in Bangladesh: Dependencies and Dynamics of a Patron-Client Relationship. Front. Vet. Sci. 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Hoque, R.; Ahmed, S.M.; Naher, N.; Islam, M.A.; Rousham, E.K.; Islam, B.Z.; Hassan, S. Tackling antimicrobial resistance in Bangladesh: A scoping review of policy and practice in human, animal and environment sectors. PLoS ONE 2020, 15, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Mund, M.D.; Khan, U.H.; Tahir, U.; Mustafa, B.E.; Fayyaz, A. Antimicrobial drug residues in poultry products and implications on public health: A review. Int. J. Food Prop. 2017, 20, 1433–1446. [Google Scholar] [CrossRef]
- Kadry, M.; Nader, S.M.; Dorgham, S.M.; Kandil, M.M. Molecular diversity of the invA gene obtained from human and egg samples. Vet. World 2019, 12, 1033–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, I. Detection of invA Gene in Isolated Salmonella from Marketed Poultry Meat by PCR Assay. J. Food Process. Technol. 2016, 7, 2. [Google Scholar] [CrossRef]
- Elkenany, R.; Elsayed, M.M.; Zakaria, A.I.; El-Sayed, S.A.E.S.; Rizk, M.A. Antimicrobial resistance profiles and virulence genotyping of Salmonella enterica serovars recovered from broiler chickens and chicken carcasses in Egypt. BMC Vet. Res. 2019, 15, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef]
- Amin, M.B.; Sraboni, A.S.; Hossain, M.I.; Roy, S.; Mozmader, T.A.U.; Unicomb, L.; Rousham, E.K.; Islam, M.A. Occurrence and genetic characteristics of mcr-1-positive colistin-resistant E. coli from poultry environments in Bangladesh. J. Glob. Antimicrob. Resist. 2020, 22, 546–552. [Google Scholar] [CrossRef]
- El Garch, F.; de Jong, A.; Bertrand, X.; Hocquet, D.; Sauget, M. mcr-1-like detection in commensal Escherichia coli and Salmonella spp. from food-producing animals at slaughter in Europe. Vet. Microbiol. 2018, 213, 42–46. [Google Scholar] [CrossRef]
- Sobur, A.; Hasan, M.; Haque, E.; Mridul, A.I.; Noreddin, A.; El Zowalaty, M.E.; Rahman, T. Molecular detection and antibiotyping of multidrug-resistant Salmonella isolated from houseflies in a fish market. Pathogens 2019, 8, 191. [Google Scholar] [CrossRef] [Green Version]
- Gad, A.H.; Abo-Shama, U.H.; Harclerode, K.K.; Fakhr, M.K. Prevalence, serotyping, molecular typing, and antimicrobial resistance of Salmonella isolated from conventional and organic retail ground poultry. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 2018, 45, 493–496. [Google Scholar] [CrossRef]
- CLSI. Clinical and Laboratory Standards Institute: Performance Standards for Antimicrobial Susceptibility Testing Supplement M100S, 26th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Satlin, M.J.; Weinstein, M.P.; Patel, J.; Romney, M.; Kahlmeter, G.; Giske, C.G.; Turnidge, J. Clinical and Laboratory Standards Institute and European Committee on Antimicrobial Susceptibility Testing Position Statements on Polymyxin B and Colistin Clinical Breakpoints. Clin. Infect. Dis. 2020, 71, e523–e529. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.T.; Lubbers, B.V.; Schwarz, S.; Watts, J.L. Applying definitions for multidrug resistance, extensive drug resistance and pandrug resistance to clinically significant livestock and companion animal bacterial pathogens. J. Antimicrob. Chemother. 2018, 73, 1460–1463. [Google Scholar] [CrossRef] [PubMed]
- Dashti, A.A.; Jadaon, M.M.; Abdulsamad, A.M.; Dashti, H.M. Heat treatment of bacteria: A simple method of DNA extraction for molecular techniques. Kuwait Med. J. 2009, 41, 117–122. [Google Scholar]
- Khan, M.N.K.; Das, M.R.; Sabur, M.A.; Rahman, M.M.; Uddin, M.B.; Cho, H.S.; Hossain, M.M.; Sciences, B. Isolation, identification, molecular detection and sensitivity to antibiotics of Salmonella from cattle faeces. Bulg. J. Vet. Med. 2019. [Google Scholar] [CrossRef]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-coffee: A novel method for fast and accurate multiple sequence alignment. Microb. Pathog. 2019, 130, 19–37. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes1. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Zhang, Y. Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys. J. 2011, 101, 2525–2534. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liang, Y.; Zhang, Y. Atomic-level protein structure refinement using fragment-guided molecular dynamics conformation sampling. Structure 2011, 19, 1784–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovell, S.C.; Davis, I.W.; Adrendall, W.B.; de Bakker, P.I.W.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Calpha geometry: Phi, psi and Cbeta deviation. Proteins 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Shoichet, B.K. ZINC--a free database of commercially available compounds for virtual screening. J. Chem. Inf. Model. 2005, 45, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, N.; Addess, K.J.; Bluhm, W.F.; Merino-Ott, J.C.; Townsend-Merino, W.; Zhang, Q.; Knezevich, C.; Xie, L.; Chen, L.; Feng, Z.; et al. The RCSB Protein Databa Bank: A redesigned query system and relational database based on the mmCIF schema. Nucleic Acids Res. 2005, 33, 233–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. In Chemical Biology; Springer Nature Switzerland AG: Cham, Switzerland, 2014; pp. 243–250. ISBN 978-1-4939-2268-0. [Google Scholar]
- Temml, V.; Kaserer, T.; Kutil, Z.; Landa, P.; Vanek, T.; Schuster, D. Pharmacophore modeling for COX-1 and-2 inhibitors with LigandScout in comparison to Discovery Studio. Future Med. Chem. 2014, 6, 1869–1881. [Google Scholar] [CrossRef]
- Parvej, M.S.; Rahman, M.; Uddin, M.F.; Nazir, K.N.H.; Jowel, M.S.; Khan, M.F.R.; Rahman, M.B. Isolation and Characterization of Salmonella enterica Serovar Typhimurium Circulating Among Healthy Chickens of Bangladesh. Turkish J. Agric. Food Sci. Technol. 2016, 4, 519. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Fu, Y.; Xiong, Z.; Ma, Y.; Wei, Y.; Qu, X.; Zhang, H.; Zhang, J.; Liao, M. Highly prevalent multidrug-resistant Salmonella from chicken and pork meat at retail markets in Guangdong, China. Front. Microbiol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Clifford, K.; Desai, D.; da Costa, C.P.; Meyer, H.; Klohe, K.; Winkler, A.; Rahman, T.; Islam, T.; Zaman, M.H. Antimicrobial resistance in livestock and poor quality veterinary medicines. Bull. World Health Organ. 2018, 96, 662–664. [Google Scholar] [CrossRef]
- Wales, A.D.; Davies, R.H. Co-selection of resistance to antibiotics, biocides and heavy metals, and its relevance to foodborne pathogens. Antibiotics 2015, 4, 567–604. [Google Scholar] [CrossRef] [Green Version]
- Faruque, M.O.; Mahmud, S.; Munayem, M.A.; Sultana, R.; Molla, M.T.; Ali, M.F.; Wasim, M.; Sarker, S.; Evamoni, F.Z. Bacteriological Analysis and Public Health Impact of Broiler Meat: A Study on Nalitabari Paurosova, Sherpur, Bangladesh. Adv. Microbiol. 2019, 9, 581–601. [Google Scholar] [CrossRef] [Green Version]
- Mridha, D.; Uddin, M.N.; Alam, B.; Akhter, A.H.M.T.; Islam, S.K.S.; Islam, M.S.; Khan, M.S.R.; Kabir, S.M.L. Identification and characterization of Salmonella spp. From samples of broiler farms in selected districts of Bangladesh. Vet. World 2020, 13, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Wajid, M.; Saleemi, M.K.; Sarwar, Y.; Ali, A. Detection and characterization of multidrug-resistant Salmonella enterica serovar Infantis as an emerging threat in poultry farms of Faisalabad, Pakistan. J. Appl. Microbiol. 2019, 127, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Akhi, M.A.; Das, N.C.; Banik, A.; Abony, M.; Juthi, M.; Uddin, M.E. Detection of Drug-resistant S. aureus from Poultry Samples Collected from Different Areas of Bangladesh. Microbiol. Res. J. Int. 2019, 29, 1–10. [Google Scholar] [CrossRef]
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Schrauwen, E.J.A.; Huizinga, P.; van Spreuwel, N.; Verhulst, C.; Kluytmans-van den Bergh, M.F.Q.; Kluytmans, J.A.J.W. High prevalence of the mcr-1 gene in retail chicken meat in the Netherlands in 2015. Antimicrob. Resist. Infect. Control 2017, 6, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Lima, T.; Domingues, S.; Da Silva, G.J. Plasmid-mediated colistin resistance in Salmonella enterica: A review. Microorganisms 2019, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Yu, L.; Chen, X.; Zhi, C.; Yao, X.; Liu, Y.; Wu, S.; Guo, Z.; Yi, L.; Zeng, Z.; et al. High prevalence of colistin resistance and mcr-1 gene in Escherichia coli isolated from food animals in China. Front. Microbiol. 2017, 8, 1–5. [Google Scholar] [CrossRef]
- Skov, R.L.; Monnet, D.L. Plasmid-mediated colistin resistance (mcr-1 gene): Three months later, the story unfolds. Eurosurveillance 2016, 21, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Carnevali, C.; Morganti, M.; Scaltriti, E.; Bolzoni, L.; Pongolini, S.; Casadei, G. Occurrence of mcr-1 in colistin-resistant Salmonella enterica isolates recovered from humans and animals in Italy, 2012 to 2015. Antimicrob. Agents Chemother. 2016, 60, 7532–7534. [Google Scholar]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a Novel mcr-6 to mcr-9 Multiplex PCR and Assessment of mcr-1 to mcr-9 Occurrence in Colistin-Resistant Salmonella enterica Isolates From Environment, Feed, Animals and Food (2011–2018) in Germany. Front. Microbiol. 2020, 11, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doumith, M.; Godbole, G.; Ashton, P.; Larkin, L.; Dallman, T.; Day, M.; Day, M.; Muller-Pebody, B.; Ellington, M.J.; de Pinna, E.; et al. Detection of the plasmid-mediated mcr-1 gene conferring colistin resistance in human and food isolates of Salmonella enterica and Escherichia coli in England and Wales. J. Antimicrob. Chemother. 2016, 71, 2300–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zając, M.; Sztromwasser, P.; Bortolaia, V.; Leekitcharoenphon, P.; Cavaco, L.M.; Ziȩtek-Barszcz, A.; Hendriksen, R.S.; Wasyl, D. Occurrence and Characterization of mcr-1-Positive Escherichia coli Isolated From Food-Producing Animals in Poland, 2011–2016. Front. Microbiol. 2019, 10, 1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.; Das, T.; Islam, M.Z.; Herrero-Fresno, A.; Biswas, P.K.; Olsen, J.E. High prevalence of mcr-1-encoded colistin resistance in commensal Escherichia coli from broiler chicken in Bangladesh. Sci. Rep. 2020, 10, 18637. [Google Scholar] [CrossRef] [PubMed]
- Ghafur, A.; Shankar, C.; GnanaSoundari, P.; Venkatesan, M.; Mani, D.; Thirunarayanan, M.A.; Veeraraghavan, B. Detection of chromosomal and plasmid-mediated mechanisms of colistin resistance in Escherichia coli and Klebsiella pneumoniae from Indian food samples. J. Glob. Antimicrob. Resist. 2019, 16, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Mohsin, M.; Lei, S.; Srinivas, S.; Wiqar, R.T.; Lin, J.; Feng, Y. Discovery of a mcr-1-bearing plasmid in commensal colistin-resistant Escherichia coli from healthy broilers in Faisalabad, Pakistan. Virulence 2018, 9, 994–999. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.S.; Song, J.; Kim, J.; Shin, J. Increasing prevalence of multidrug-resistant mcr-1-positive Escherichia coli isolates from fresh vegetables and healthy food animals in South Korea. Int. J. Infect. Dis. 2020, 92, 53–55. [Google Scholar] [CrossRef] [Green Version]
- Trung, N.V.; Matamoros, S.; Carrique-Mas, J.J.; Nghia, N.H.; Nhung, N.T.; Chieu, T.T.B.; Mai, H.H.; van Rooijen, W.; Campbell, J.; Wagenaar, J.A.; et al. Zoonotic transmission of mcr-1 colistin resistance gene from small-scale poultry farms, Vietnam. Emerg. Infect. Dis. 2017, 23, 529–532. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Hu, Y.; Li, Z.; Sun, J.; Wang, Q.; Lin, J.; Ye, H.; Liu, F.; Srinivas, S.; Li, D.; et al. Dissemination and Mechanism for the MCR-1 Colistin Resistance. PLoS Pathog. 2016, 12, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wei, W.; Lei, S.; Lin, J.; Srinivas, S.; Feng, Y. An evolutionarily conserved mechanism for intrinsic and transferable polymyxin resistance. MBio 2018, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Name | Primer Sequence | Size (bp) |
|---|---|---|---|
| MCR1 | MCR1-P1F MCR1-P1R | F: CAGTATGGGATTGCGCAATGATT R: TTATCCATCACGCCTTTTGAGTC | 1197 |
| MCR1 | MCR1-P2F MCR1-P2R | F: TGTCGATACCGCCAAATACCAAG R: GGAGTGTGCGGTGGGTTTG | 799 |
| Antimicrobial Agents | Susceptible (S) | Intermediate (I) | Resistance (R) | I + R | |||
|---|---|---|---|---|---|---|---|
| Number of Isolates | % | Number of Isolates | % | Number of Isolates | % | ||
| Penicillins | |||||||
| Ampicillin (AMP, 10 μg) | 70 | 85.37 | 4 | 4.88 | 8 | 9.76 | 14.63 |
| Amoxycillin/clavulanate (AMC, 20/10 µg) | 45 | 54.88 | 3 | 3.66 | 34 | 41.46 | 45.12 |
| Piperacillin/tazobactam (PTZ, 100/10 μg) | 69 | 84.15 | 6 | 7.32 | 8 | 8.54 | 15.85 |
| Aminoglycosides | |||||||
| Amikacin (AMK, 30 μg) | 53 | 64.63 | 7 | 8.54 | 22 | 26.83 | 35.37 |
| Gentamicin (GEN, 10 μg) | 47 | 57.32 | 8 | 9.76 | 27 | 32.93 | 42.68 |
| Cephalosporins | |||||||
| Cefuroxime (CFX, 30 μg) | 46 | 56.10 | 10 | 12.20 | 26 | 31.71 | 43.90 |
| Cefuroxime axetil (CFA, 30 μg) | 44 | 53.66 | 21 | 25.61 | 17 | 20.73 | 46.34 |
| Ceftriaxone (CTR, 30 μg) | 43 | 52.44 | 6 | 7.32 | 33 | 40.24 | 47.56 |
| Cefoperazone/sulbactam (CFS, 75/30 μg) | 42 | 51.22 | 33 | 40.24 | 7 | 8.54 | 48.78 |
| Cefepime (CFP, 30 μg) | 38 | 46.34 | 29 | 35.37 | 15 | 18.29 | 53.66 |
| Carbapenems | |||||||
| Ertapenem (ETP, 10 μg) | 45 | 54.88 | 32 | 39.02 | 5 | 6.10 | 45.12 |
| Imipenem (IMP, 10 μg) | 56 | 68.29 | 5 | 6.10 | 21 | 25.61 | 31.71 |
| Meropenem (MPM, 10 μg) | 48 | 58.54 | 6 | 7.32 | 28 | 34.15 | 41.46 |
| Tetracyclines | |||||||
| Tigecycline (TIG, 15 μg) | 23 | 28.05 | 8 | 9.76 | 51 | 62.20 | 71.95 |
| Fluoroquinolones | |||||||
| Ciprofloxacin (CIP, 5 μg) | 16 | 19.51 | 6 | 7.32 | 60 | 73.17 | 80.49 |
| Nitrofurans | |||||||
| Nitrofurantoin (NIT, 300 µg) | 60 | 73.17 | 8 | 9.76 | 14 | 17.07 | 26.83 |
| Polymyxins | |||||||
| Colistin (COL, 10 μg) | 0 | 0.00 | 6 | 7.32 | 76 | 92.68 | 100.00 |
| Folate Pathway Inhibitors | |||||||
| Trimethoprim/Sulfamethoxazole (SXT, 1.25/23.75 µg) | 23 | 28.05 | 9 | 10.98 | 50 | 60.98 | 71.95 |
| COL | SXT | CIP | TIG | AMC | CTR | GEN * | AMK * | IMP | MPM | NIT | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolate Number | (10 μg) | (25 μg) | (5 μg) | (15 μg) | (30 μg) | (30 μg) | (10 μg) | (30 μg) | (10 μg) | (10 μg) | (300 μg) |
| SAUVM S6 | R | R | R | R | S | S | S | S | R | R | R |
| SAUVM S7 | R | R | R | S | R | R | R | R | S | S | S |
| SAUVM S8 | R | S | R | R | S | R | R | S | S | S | R |
| SAUVM S9 | R | R | R | S | S | S | S | R | R | R | S |
| SAUVM S10 | R | S | R | R | R | S | R | S | S | S | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uddin, M.B.; Hossain, S.M.B.; Hasan, M.; Alam, M.N.; Debnath, M.; Begum, R.; Roy, S.; Harun-Al-Rashid, A.; Chowdhury, M.S.R.; Rahman, M.M.; et al. Multidrug Antimicrobial Resistance and Molecular Detection of mcr-1 Gene in Salmonella Species Isolated from Chicken. Animals 2021, 11, 206. https://doi.org/10.3390/ani11010206
Uddin MB, Hossain SMB, Hasan M, Alam MN, Debnath M, Begum R, Roy S, Harun-Al-Rashid A, Chowdhury MSR, Rahman MM, et al. Multidrug Antimicrobial Resistance and Molecular Detection of mcr-1 Gene in Salmonella Species Isolated from Chicken. Animals. 2021; 11(1):206. https://doi.org/10.3390/ani11010206
Chicago/Turabian StyleUddin, Md Bashir, S.M. Bayejed Hossain, Mahmudul Hasan, Mohammad Nurul Alam, Mita Debnath, Ruhena Begum, Sawrab Roy, Ahmed Harun-Al-Rashid, Md. Shahidur Rahman Chowdhury, Md. Mahfujur Rahman, and et al. 2021. "Multidrug Antimicrobial Resistance and Molecular Detection of mcr-1 Gene in Salmonella Species Isolated from Chicken" Animals 11, no. 1: 206. https://doi.org/10.3390/ani11010206
APA StyleUddin, M. B., Hossain, S. M. B., Hasan, M., Alam, M. N., Debnath, M., Begum, R., Roy, S., Harun-Al-Rashid, A., Chowdhury, M. S. R., Rahman, M. M., Hossain, M. M., Elahi, F., Chowdhury, M. Y. E., Järhult, J. D., El Zowalaty, M. E., & Ahmed, S. S. U. (2021). Multidrug Antimicrobial Resistance and Molecular Detection of mcr-1 Gene in Salmonella Species Isolated from Chicken. Animals, 11(1), 206. https://doi.org/10.3390/ani11010206