The Roles of the miRNAome and Transcriptome in the Ovine Ovary Reveal Poor Efficiency in Juvenile Superovulation


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Superovulation Treatment
2.2. Sample Collection and RNA Extraction
2.3. Library Construction and Sequencing
2.4. Read Filtering and Differential Expression Analysis
2.5. Targeted Relationship between Differentially Expressed miRNAs and mRNAs
2.6. Enrichment Analysis by GO and KEGG
2.7. Real-Time PCR Validation
2.8. Statistical Analysis
3. Results
3.1. Measurement of Follicle Numbers and Serum Hormone in Lamb and Adult Sheep under Superovulation Treatment
3.2. Overview of Sequencing Data
3.3. Differentially Expressed miRNA and mRNA in the Lamb and Adult Libraries
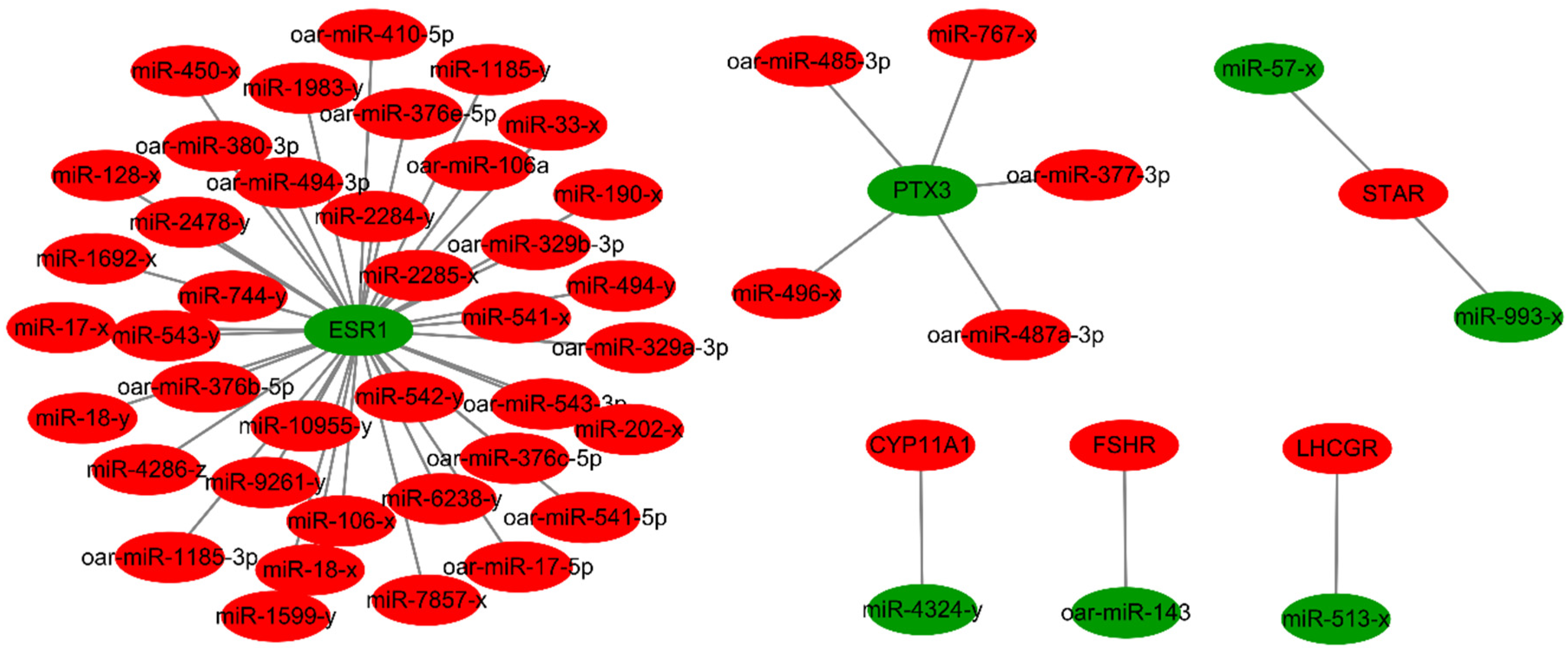
3.4. Analysis of the Targeted Relationship between Differentially Expressed miRNAs and mRNAs
3.5. GO and KEGG Enrichment Analysis for Target Genes of Differentially Expressed miRNAs
3.6. GO and KEGG Enrichment Analysis of Differentially Expressed mRNAs
3.7. Real-Time PCR Validation of mRNA and miRNA Associated with Hormone Receptor and Oocyte Quality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morton, K.M.; Catt, S.L.; Maxwell, W.M.C.; Evans, G. An efficient method of ovarian stimulation and in vitro embryo production from prepubertal lambs. Reprod. Fertil. Dev. 2005, 17, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.; Smith, H.; McGrice, H.A.; Kind, K.L.; van Wettere, W. Towards Improving the Outcomes of Assisted Reproductive Technologies of Cattle and Sheep, with Particular Focus on Recipient Management. Animals (Basel) 2020, 10, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, D.T.; Kotaras, P.J.; Earl, C.R. Advances in production of embryos in vitro from juvenile and prepubertal oocytes from the calf and lamb. Reprod. Fertil. Dev. 1997, 9, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Valasi, I.; Menegatos, I.; Papanikolaou, T.; Goulas, P.; Amiridis, G.S. Oocyte pick-up in juvenile lambs affects neither onset of puberty nor their future fertility. Theriogenology 2006, 66, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Ptak, G.; Loi, P.; Dattena, M.; Tischner, M.; Cappai, P. Offspring from one-month-old lambs: Studies on the developmental capability of prepubertal oocytes. Biol. Reprod. 1999, 61, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function; Elsevier Inc.: Philadelphia, PA, USA, 2004; Volume 116, pp. 281–297. [Google Scholar]
- Azhar, S.; Dong, D.; Shen, W.J.; Hu, Z.; Kraemer, F.B. The role of miRNAs in regulating adrenal and gonadal steroidogenesis. J. Mol. Endocrinol. 2020, 64, R21–R43. [Google Scholar] [CrossRef]
- Hayashi, T.; Hoffman, M.P. Exosomal microRNA communication between tissues during organogenesis. RNA Biol. 2017, 14, 1683–1689. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Guo, X.; Cheng, L.; Chu, P.; Chen, M.; Chen, Y.; Chang, C. An integrated analysis of the circRNA-miRNA-mRNA network reveals novel insights into potential mechanisms of cell proliferation during liver regeneration. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3873–3884. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Wu, Q.; Lu, D.; Yu, J.; Rao, Y.; Kou, Z.; Fang, G.; Liu, W.; Han, H. A systematic study of critical miRNAs on cells proliferation and apoptosis by the shortest path. BMC Bioinform. 2020, 21, 1–14. [Google Scholar] [CrossRef]
- Martin, E.C.; Qureshi, A.T.; Dasa, V.; Freitas, M.A.; Gimble, J.M.; Davis, T.A. MicroRNA regulation of stem cell differentiation and diseases of the bone and adipose tissue: Perspectives on miRNA biogenesis and cellular transcriptome. Biochimie 2016, 124, 98–111. [Google Scholar] [CrossRef]
- Rottiers, V.; Naar, A.M. MicroRNAs in metabolism and metabolic disorders. Nat. Rev. Mol. Cell Biol. 2012, 13, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, N.; Sherman-Baust, C.A.; Wang, T.L.; Davidson, B.; Shih, I.; Zhang, Y.; Wood, W.R.; Becker, K.G.; Morin, P.J. MicroRNA expression and identification of putative miRNA targets in ovarian cancer. PLoS ONE 2008, 3, e2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Ren, G.; Liu, J.; Zhao, Z.; Yu, Y.; Su, R.; Ma, X.; Ni, H.; Lei, W.; Yang, Z. MicroRNA Expression and Regulation in Mouse Uterus during Embryo Implantation. J. Biol. Chem. 2008, 283, 23473–23484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donadeu, F.X.; Schauer, S.N.; Sontakke, S.D. Involvement of miRNAs in ovarian follicular and luteal development. J. Endocrinol. 2012, 215, 323–334. [Google Scholar] [CrossRef]
- Tesfaye, D.; Gebremedhn, S.; Salilew-Wondim, D.; Hailay, T.; Hoelker, M.; Grosse-Brinkhaus, C.; Schellander, K. MicroRNAs: Tiny molecules with a significant role in mammalian follicular and oocyte development. Reproduction 2018, 155, R121–R135. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, M.; Zheng, M.; Hayashi, M.; Lee, J.D.; Yoshino, O.; Lin, S.; Han, J. Impaired microRNA processing causes corpus luteum insufficiency and infertility in mice. J. Clin. Investig. 2008, 118, 1944–1954. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Wang, X.; Yao, G.; Lu, M.; Liang, M.; Sun, Y.; Sun, F. Transactivation of microRNA-320 by microRNA-383 regulates granulosa cell functions by targeting E2F1 and SF-1 proteins. J. Biol. Chem. 2014, 289, 18239–18257. [Google Scholar] [CrossRef] [Green Version]
- Zhai, M.; Xie, Y.; Liang, H.; Lei, X.; Zhao, Z. Comparative profiling of differentially expressed microRNAs in estrous ovaries of Kazakh sheep in different seasons. Gene 2018, 664, 181–191. [Google Scholar] [CrossRef]
- Pokharel, K.; Peippo, J.; Honkatukia, M.; Seppala, A.; Rautiainen, J.; Ghanem, N.; Hamama, T.M.; Crowe, M.A.; Andersson, M.; Li, M.H.; et al. Integrated ovarian mRNA and miRNA transcriptome profiling characterizes the genetic basis of prolificacy traits in sheep (Ovis aries). BMC Genom. 2018, 19, 104. [Google Scholar] [CrossRef] [Green Version]
- Miao, X.; Luo, Q.; Zhao, H.; Qin, X. Ovarian transcriptomic study reveals the differential regulation of miRNAs and lncRNAs related to fecundity in different sheep. Sci. Rep.-UK 2016, 6, 35299. [Google Scholar] [CrossRef]
- Morton, K.M. Developmental capabilities of embryos produced in vitro from prepubertal lamb oocytes. Reprod. Domest. Anim. 2008, 43 (Suppl. 2), 137–143. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, X.; Zhang, J.; Zhong, R.; Zhou, D. Global DNA methylation and related mRNA profiles in sheep oocytes and early embryos derived from pre-pubertal and adult donors. Anim. Reprod. Sci. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lin, J.; Li, X.; Han, B.; Wang, L.; Liu, M.; Huang, J. Transcriptome profile of one-month-old lambs’ granulosa cells after superstimulation. Asian-Australas. J. Anim. Sci. 2017, 30, 20–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths-Jones, S.; Bateman, A.; Marshall, M.; Khanna, A.; Eddy, S.R. Rfam: An RNA family database. Nucleic Acids Res. 2003, 31, 439–441. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef] [PubMed]
- Joachims, T. Svmlight: Support Vector Machine. Available online: http://svmlight.joachims.org/ (accessed on 2 January 2021).
- Turner, D.A. Miranda: A Non-Strict Functional Language with Polymorphic Types. Conference on Functional Programming Languages and Computer Architecture; Springer: Berlin/Heidelberg, Germany, 1985; Volume 201, pp. 1–16. [Google Scholar]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Du, X.; Zhang, L.; Li, X.; Pan, Z.; Liu, H.; Li, Q. TGF-beta signaling controls FSHR signaling-reduced ovarian granulosa cell apoptosis through the SMAD4/miR-143 axis. Cell Death Dis. 2016, 7, e2476. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Chen, C.Z.; Xu, M.Q.; Zhang, L.Q.; Liu, J.B.; Gao, Y.; Jiang, H.; Yuan, B.; Zhang, J.B. MiR-31 and miR-143 affect steroid hormone synthesis and inhibit cell apoptosis in bovine granulosa cells through FSHR. Theriogenology 2019, 123, 45–53. [Google Scholar] [CrossRef]
- Troppmann, B.; Kossack, N.; Nordhoff, V.; Schuring, A.N.; Gromoll, J. MicroRNA miR-513a-3p acts as a co-regulator of luteinizing hormone/chorionic gonadotropin receptor gene expression in human granulosa cells. Mol. Cell. Endocrinol. 2014, 390, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, X.; Zhang, X.; Lu, Y.; Li, L.; Cui, S. miRNA-143 mediates the proliferative signaling pathway of FSH and regulates estradiol production. J. Endocrinol. 2017, 234, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, H.; Sheng, Y.; Wang, Z. MicroRNA-224 delays oocyte maturation through targeting Ptx3 in cumulus cells. Mech. Dev. 2017, 143, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Paramio, M.T.; Izquierdo, D. Current status of in vitro embryo production in sheep and goats. Reprod. Domest. Anim. 2014, 49 (Suppl. 4), 37–48. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Zhang, J.; Li, Y.; Guo, X.; Li, J.; Zhong, R.; Zhang, X. Melatonin-induced demethylation of antioxidant genes increases antioxidant capacity through RORalpha in cumulus cells of prepubertal lambs. Free Radical Biol. Med. 2019, 131, 173–183. [Google Scholar] [CrossRef]
- Van Blerkom, J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 2011, 11, 797–813. [Google Scholar] [CrossRef]
- Ptak, G.; Matsukawa, K.; Palmieri, C.; Della, S.L.; Scapolo, P.A.; Loi, P. Developmental and functional evidence of nuclear immaturity in prepubertal oocytes. Hum. Reprod. 2006, 21, 2228–2237. [Google Scholar] [CrossRef] [Green Version]
- Warzych, E.; Pawlak, P.; Pszczola, M.; Cieslak, A.; Lechniak, D. Prepubertal heifers versus cows-The differences in the follicular environment. Theriogenology 2017, 87, 36–47. [Google Scholar] [CrossRef]
- Wu, Y.; Lin, J.; Han, B.; Wang, L.; Chen, Y.; Liu, M.; Huang, J. Proteomic profiling of follicle fluids after superstimulation in one-month-old lambs. Reprod. Domest. Anim. 2018, 53, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Liu, K.; Zhang, Y.; Qi, Q.; Wang, C.; Guan, H.; Yan, F.; Hou, J. Adult follicular fluid supplementation during in vitro maturation improves the developmental competence of prepubertal lamb oocytes. Theriogenology 2019, 130, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V.; Ovcharenko, D.; Grossmann, R.; Laukova, M.; Mlyncek, M. Identification of microRNAs controlling human ovarian cell steroidogenesis via a genome-scale screen. J. Cell. Physiol. 2009, 219, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Jiajie, T.; Yanzhou, Y.; Hoi-Hung, A.C.; Zi-Jiang, C.; Wai-Yee, C. Conserved miR-10 family represses proliferation and induces apoptosis in ovarian granulosa cells. Sci. Rep.-UK 2017, 7, 41304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, G.; Zhang, L.; Fang, T.; Zhang, Q.; Wu, S.; Jiang, Y.; Sun, H.; Hu, Y. MicroRNA-145 suppresses mouse granulosa cell proliferation by targeting activin receptor IB. FEBS Lett. 2012, 586, 3263–3270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ascari, I.J.; Alves, N.G.; Alves, A.C.; Garcia, I.F.F.; Junqueira, F.B. Resumption of cyclic ovarian activity in postpartum ewes: A review. Semin. Ciên. Agrár. 2016, 37, 1101–1116. [Google Scholar] [CrossRef] [Green Version]
- Satue, K.; Fazio, E.; Medica, P. Can the Presence of Ovarian Corpus Luteum Modify the Hormonal Composition of Follicular Fluid in Mares? Animals (Basel) 2020, 10, 646. [Google Scholar] [CrossRef] [Green Version]
- Ling, Y.H.; Guo, X.F.; Chen, T.; Ding, J.P.; Ma, Y.H.; Chu, M.X.; Di, R.; Zhang, Y.H.; Zhang, X.R. Characterization and analysis of differentially expressed microRNAs in hircine ovaries during the follicular and luteal phases. Anim. Reprod. Sci. 2016, 166, 47–57. [Google Scholar] [CrossRef]
- Scarlet, D.; Walter, I.; Hlavaty, J.; Aurich, C. Expression and immunolocalisation of follicle-stimulating hormone receptors in gonads of newborn and adult female horses. Reprod. Fertil. Dev. 2016, 28, 1340–1348. [Google Scholar] [CrossRef]
- Dierich, A.; Sairam, M.R.; Monaco, L.; Fimia, G.M.; Gansmuller, A.; LeMeur, M.; Sassone-Corsi, P. Impairing follicle-stimulating hormone (FSH) signaling in vivo: Targeted disruption of the FSH receptor leads to aberrant gametogenesis and hormonal imbalance. Proc. Natl. Acad. Sci. USA 1998, 95, 13612–13617. [Google Scholar] [CrossRef] [Green Version]
- Michalovic, L.; Currin, L.; Gutierrez, K.; Bellefleur, A.M.; Glanzner, W.G.; Schuermann, Y.; de Macedo, M.P.; Bohrer, R.C.; Dicks, N.; Lopez, R.; et al. Granulosa cells of prepubertal cattle respond to gonadotropin signaling and upregulate genes that promote follicular growth and prevent cell apoptosis. Mol. Reprod. Dev. 2018, 85, 909–920. [Google Scholar] [CrossRef]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couse, J.F.; Korach, K.S. Estrogen receptor null mice: What have we learned and where will they lead us? Endocr. Rev. 1999, 20, 358–417. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xin, Q.; Wang, X.; Wang, S.; Wang, H.; Zhang, W.; Yang, Y.; Zhang, Y.; Zhang, Z.; Wang, C.; et al. Estrogen receptors in granulosa cells govern meiotic resumption of pre-ovulatory oocytes in mammals. Cell Death Dis. 2017, 8, e2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artini, P.G.; Tatone, C.; Sperduti, S.; D’Aurora, M.; Franchi, S.; Di Emidio, G.; Ciriminna, R.; Vento, M.; Di Pietro, C.; Stuppia, L.; et al. Cumulus cells surrounding oocytes with high developmental competence exhibit down-regulation of phosphoinositol 1,3 kinase/protein kinase B (PI3K/AKT) signalling genes involved in proliferation and survival. Hum. Reprod. 2017, 32, 2474–2484. [Google Scholar] [CrossRef] [PubMed]
- Palma, G.A.; Tortonese, D.J.; Sinowatz, F. Developmental capacity in vitro of prepubertal oocytes. Anat. Histol. Embryol. 2001, 30, 295–300. [Google Scholar] [CrossRef]
- Zhang, X.; Jafari, N.; Barnes, R.B.; Confino, E.; Milad, M.; Kazer, R.R. Studies of gene expression in human cumulus cells indicate pentraxin 3 as a possible marker for oocyte quality. Fertil. Steril. 2005, 83 (Suppl. 1), 1169–1179. [Google Scholar] [CrossRef]
- Kona, S.S.; Praveen, C.V.; Siva, K.A.; Srividya, D.; Padmaja, K.; Rao, V.H. Quantitative expression patterns of GDF9 and BMP15 genes in sheep ovarian follicles grown in vivo or cultured in vitro. Theriogenology 2016, 85, 315–322. [Google Scholar] [CrossRef]
- Wong, A.C.; Ryan, A.F. Mechanisms of sensorineural cell damage, death and survival in the cochlea. Front. Aging Neurosci. 2015, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Boone, D.L.; Tsang, B.K. Caspase-3 in the rat ovary: Localization and possible role in follicular atresia and luteal regression. Biol. Reprod. 1998, 58, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.H.; Chang, C.L.; Wu, H.M.; Chiu, Y.M.; Chen, C.K.; Wang, H.S. Insulin-like growth factor-II (IGF-II), IGF-binding protein-3 (IGFBP-3), and IGFBP-4 in follicular fluid are associated with oocyte maturation and embryo development. Fertil. Steril. 2006, 86, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Liu, H.; Han, Y.; Chen, Y.; Jiang, H. Integrated analysis of miRNA and mRNA expression profiles in 2-, 6-, and 12-month-old Small Tail Han Sheep ovaries reveals that oar-miR-432 downregulates RPS6KA1 expression. Gene 2019, 710, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Christenson, L.K. MicroRNA control of ovarian function. Anim. Reprod. 2010, 7, 129–133. [Google Scholar]
- Ling, Y.H.; Ren, C.H.; Guo, X.F.; Xu, L.N.; Huang, Y.F.; Luo, J.C.; Zhang, Y.H.; Zhang, X.R.; Zhang, Z.J. Identification and characterization of microRNAs in the ovaries of multiple and uniparous goats (Capra hircus) during follicular phase. BMC Genom. 2014, 15, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, M.; Bo, X.; Li, T.; Ma, L.; Zhai, T.; Huang, T. Systematic Analysis of Long Non-Coding RNAs and mRNAs in the Ovaries of Duroc Pigs During Different Follicular Stages Using RNA Sequencing. Int. J. Mol. Sci. 2018, 19, 1722. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, C.; Li, Q.; Li, W.T.; Li, H.; Li, G.X.; Kang, X.T.; Liu, X.J.; Tian, Y.D. Identification of the Key microRNAs and miRNA-mRNA Interaction Networks during the Ovarian Development of Hens. Animals (Basel) 2020, 10, 1680. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2008, 19, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Wu, Y.; Han, B.; Chen, Y.; Wang, L.; Li, X.; Liu, M.; Huang, J. iTRAQ-based proteomic profiling of granulosa cells from lamb and ewe after superstimulation. Theriogenology 2017, 101, 99–108. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of Animals | Average Number of Follicles | FSH (mIU/mL) | LH (mIU/mL) | P4 (10−2 ng/mL) | E2 (pg/mL) |
|---|---|---|---|---|---|---|
| Lamb | 6 | 70.17 ± 5.14 a | 3.65 ± 0.35 | 3.66 ± 0.37 | 1.36 ± 0.24 | 4.43 ± 0.37 |
| Adult | 6 | 20.17 ± 2.65 b | 3.61 ± 0.32 | 3.88 ± 0.10 | 1.33 ± 0.11 | 4.67 ± 0.30 |
| miRNA | Lamb_Mean | Adult_Mean | log2(fc) | p-Value | FDR |
|---|---|---|---|---|---|
| miR-145-x | 7649.92 | 23730.58 | –1.63 | 3.43 × 10−25 | 1.04 × 10−22 |
| miR-144-y | 6678.34 | 286.18 | 4.54 | 3.79 × 10−9 | 9.32 × 10−8 |
| miR-451-x | 4928.68 | 578.77 | 3.09 | 4.28 × 10−4 | 3.33 × 10−3 |
| miR-450-x | 3914.41 | 721.04 | 2.44 | 6.42 × 10−6 | 9.05 × 10−5 |
| miR-486-x | 3379.14 | 327.35 | 3.37 | 1.22 × 10−4 | 1.11 × 10−3 |
| miR-424-x | 2801.34 | 448.96 | 2.64 | 3.14 × 10−7 | 5.34 × 10−6 |
| miR-202-x | 2696.87 | 316.41 | 3.09 | 1.93 × 10−11 | 7.79 × 10−10 |
| oar-miR-10a | 1594.46 | 4637.33 | –1.54 | 2.30 × 10−11 | 9.09 × 10−10 |
| oar-miR-3958-3p | 1497.39 | 392.70 | 1.93 | 4.71 × 10−2 | 1.65 × 10−1 |
| oar-miR-409-3p | 1485.38 | 197.24 | 2.91 | 5.88 × 10−11 | 2.02 × 10−9 |
| Gene | Lamb_Mean | Adult_Mean | log2(fc) | FDR | Gene Classification |
|---|---|---|---|---|---|
| FSHR | 5.93 | 1.49 | 1.99 | 4.63 × 10−7 | HC |
| LHCGR | 84.10 | 16.31 | 2.37 | 1.77 × 10−29 | HC |
| ESR1 | 8.89 | 22.16 | −1.32 | 3.77 × 10−13 | HC |
| CYP11A1 | 251.29 | 41.33 | 2.60 | 3.66 × 10−42 | SHE |
| CYP17A1 | 74.87 | 4.64 | 4.01 | 1.71 × 10−15 | SHE |
| CYP19A1 | 50.23 | 1.37 | 5.20 | 2.46 × 10−14 | SHE |
| STAR | 32.56 | 11.31 | 1.53 | 5.09 × 10−13 | SHE |
| HSD3B | 112.84 | 39.14 | 1.53 | 6.76 × 10−3 | SHE |
| HSD17B1 | 87.11 | 6.59 | 3.73 | 1.15 × 10−7 | SHE |
| PTX3 | 4.02 | 20.19 | −2.33 | 2.75 × 10−40 | ROQ |
| GDF9 | 4.07 | 1.33 | 1.61 | 6.91 × 10−4 | ROQ |
| DNMT1 | 28.33 | 12.30 | 1.20 | 2.21 × 10−39 | ROQ |
| CASP3 | 7.19 | 1.58 | 2.18 | 2.75 × 10−8 | ROQ |
| IGFBP4 | 44.01 | 74.65 | −0.76 | 2.71 × 10−6 | ROQ |
| BCL2 | 3.28 | 5.38 | −0.71 | 8.84 × 10−3 | ROQ |
| BAX | 16.84 | 13.98 | 0.27 | 2.60 × 10−1 | ROQ |
| BAX/BCL2 | 5.13 | 2.60 | 0.98 | 7.75 × 10−3 | NA |
| id | Lamb_Mean | Adult_Mean | log2(fc) | FDR | Target Gene | Target Gene Classification |
|---|---|---|---|---|---|---|
| oar-miR-143 [34] | 70,716.65 | 122,228.40 | −0.79 | 1.98 × 10−11 | FSHR | HC |
| miR-513-x [35] | 1.09 | 2.41 | −1.14 | 2.96 × 10−4 | LHCGR | HC |
| miR-4324-y | 2.84 | 10.16 | −1.84 | 1.20 × 10−16 | CYP11A1 | SHE |
| miR-57-x | 0.05 | 0.42 | −3.13 | 1.47 × 10−3 | STAR | SHE |
| miR-993-x | 0.34 | 0.73 | −1.11 | 7.74 × 10−2 | STAR | SHE |
| oar-miR-485-3p | 60.89 | 5.71 | 3.41 | 4.33 × 10−10 | PTX3 | ROQ |
| oar-miR-487a-3p | 55.39 | 4.55 | 3.61 | 6.36 × 10−11 | PTX3 | ROQ |
| oar-miR-377-3p | 9.39 | 1.46 | 2.69 | 6.53 × 10−3 | PTX3 | ROQ |
| miR-767-x | 3.81 | 0.43 | 3.16 | 1.09 × 10−2 | PTX3 | ROQ |
| miR-496-x | 0.51 | 0.01 | 5.67 | 2.67 × 10−2 | PTX3 | ROQ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Dong, C.; Yang, J.; Li, Y.; Feng, J.; Wang, B.; Zhang, J.; Guo, X. The Roles of the miRNAome and Transcriptome in the Ovine Ovary Reveal Poor Efficiency in Juvenile Superovulation. Animals 2021, 11, 239. https://doi.org/10.3390/ani11010239
Zhang X, Dong C, Yang J, Li Y, Feng J, Wang B, Zhang J, Guo X. The Roles of the miRNAome and Transcriptome in the Ovine Ovary Reveal Poor Efficiency in Juvenile Superovulation. Animals. 2021; 11(1):239. https://doi.org/10.3390/ani11010239
Chicago/Turabian StyleZhang, Xiaosheng, Chunxiao Dong, Jing Yang, Yihai Li, Jing Feng, Biao Wang, Jinlong Zhang, and Xiaofei Guo. 2021. "The Roles of the miRNAome and Transcriptome in the Ovine Ovary Reveal Poor Efficiency in Juvenile Superovulation" Animals 11, no. 1: 239. https://doi.org/10.3390/ani11010239
APA StyleZhang, X., Dong, C., Yang, J., Li, Y., Feng, J., Wang, B., Zhang, J., & Guo, X. (2021). The Roles of the miRNAome and Transcriptome in the Ovine Ovary Reveal Poor Efficiency in Juvenile Superovulation. Animals, 11(1), 239. https://doi.org/10.3390/ani11010239