1. Introduction
Parasitic copepods use a wide range of scleractinian corals as hosts [
1]. Up to date, a total of 363 copepod species representing 99 genera, 19 families and three orders (Cyclopoda, Siphonostomatoida, and Harpactioida) have been recognized as parasites in 148 shallow-water stony corals [
2]. The total included 288 cyclopoids, 68 siphonostomatoids, and seven harpacticoids. These coral-associated copepods exhibit a marked diversity in morphology in accordance with their respective ecological niches. Among the coral-associated copepods, Xarifiidae Humes, 1960 [
3] is a family of endoparasitic copepods living in the gastrovascular cavities of coral polyps and more than 90 valid species have been discovered in Indo-West Pacific coral reefs [
2,
4,
5]. It has been evident that their virulence may be related to their life history strategies,
Symbiodinium density, surface area of host coral colonies, and concentration of nitrate and chlorophyll-
a in the surrounding seawater. Therefore, the potential of using these parasites as bioindicators for predicting the future physiological performance of host corals in response to environmental change can be developed by tracking their abundance and species composition [
2].
According to the previous studies carried out from 1967 to 2010 [
4,
6,
7,
8,
9,
10], 15 species of parasitic copepods have been discovered from a widely distributed scleractinian genus,
Psammocora Dana, 1846 [
11] throughout the Indo-West Pacific, including three highly modified cyclopoids:
Xarifia diminuta Humes and Ho, 1967 [
6],
Xarifia formosa Humes, 1985 [
4],
Xarifia imitans Humes, 1985 [
4]. Recently, Cheng and Lin reported three new species of
Xarifia,
Xarifia conrepta Cheng and Lin 2021 [
12],
Xarifia gracilis Cheng and Lin 2021 [
12], and
Xarifia lata Cheng and Lin 2021 [
12], parasitic in
Psammocora digitata Milne Edwards and Haime 1851 [
13] from Taiwan. Thus, among 92
Xarifia species recorded up to date, six of them have been discovered in
Psammocora corals. All six species can be found in
P.
digitata, while
X. diminuta also occurred in
Psammocora contigua [
14]. (
Table 1).
Obviously, the current knowledge about coral-associated copepods of the widespread
Psammocora corals remains limited. There are ten valid species of
Psammocora around the world [
15], but only two species of
Psammocora corals have been examined for coral-associated copepods.
Psammocora columna Dana, 1846 [
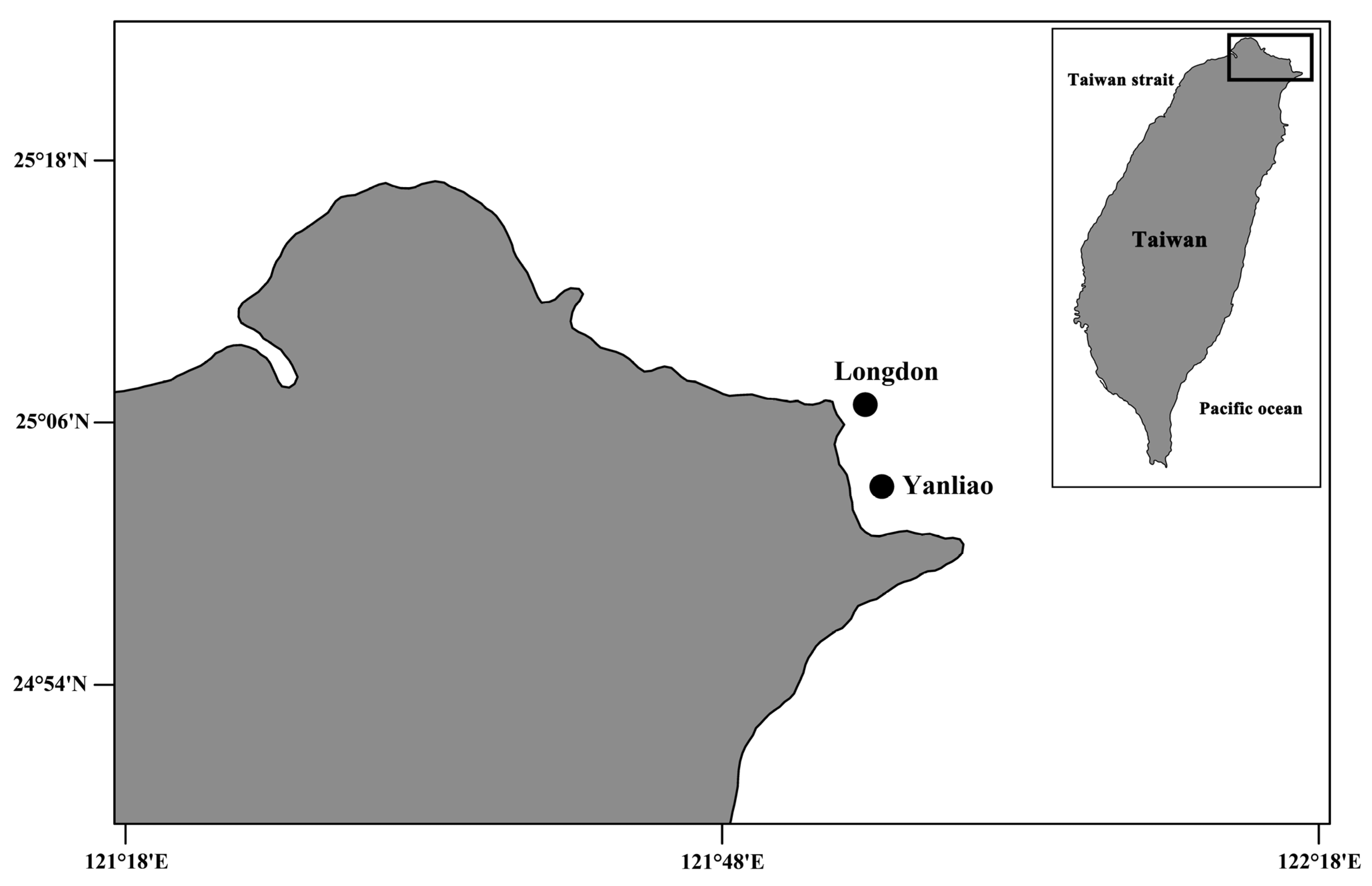
11] is a widely distributed coral and has not been examined for coral-associated copepods yet. Herein, we perform a survey in two colonies of
P.
columna collected from a shallow-water reef on the north coast of Taiwan (
Figure 1), and describe two new species of xarifiid copepods. Combining our findings with previous records, 20 species of parasitic copepods including eight species of
Xarifia have been found in
Psammocora corals.
3. Results
SYSTEMATICS. Family Xarifiidae Humes, 1960. Genus
Xarifia Humes, 1960.
Xarifia yanliaoensis sp. nov. (
Figure 3,
Figure 4 and
Figure 5).
Material examined: Three specimens (two ♀♀ and one ♂) were obtained from washings of a
Psammocora columna colony collected at 5 m depth, at Longdon (25°06′49.5″ N, 121°55′11.8″ E), Gongliao District, New Taipei, Taiwan on 12 August 2010 (
Figure 1 and
Figure 2). The other specimens (two ♀♀ and three ♂♂) were obtained from washings the other colony of the same coral species collected at 9 m depth, at Yanliao (25°03′04.4″ N 121°56′01.7″ E) on 20 August 2010 (
Figure 1 and
Figure 2). Female holotype (NTUM-Inv-10019), male allotype (NTUM-Inv-10020); paratypes (NKUST-Cop-0004) were deposited in National Taiwan University Museum (NTUM), Taipei, Taiwan.
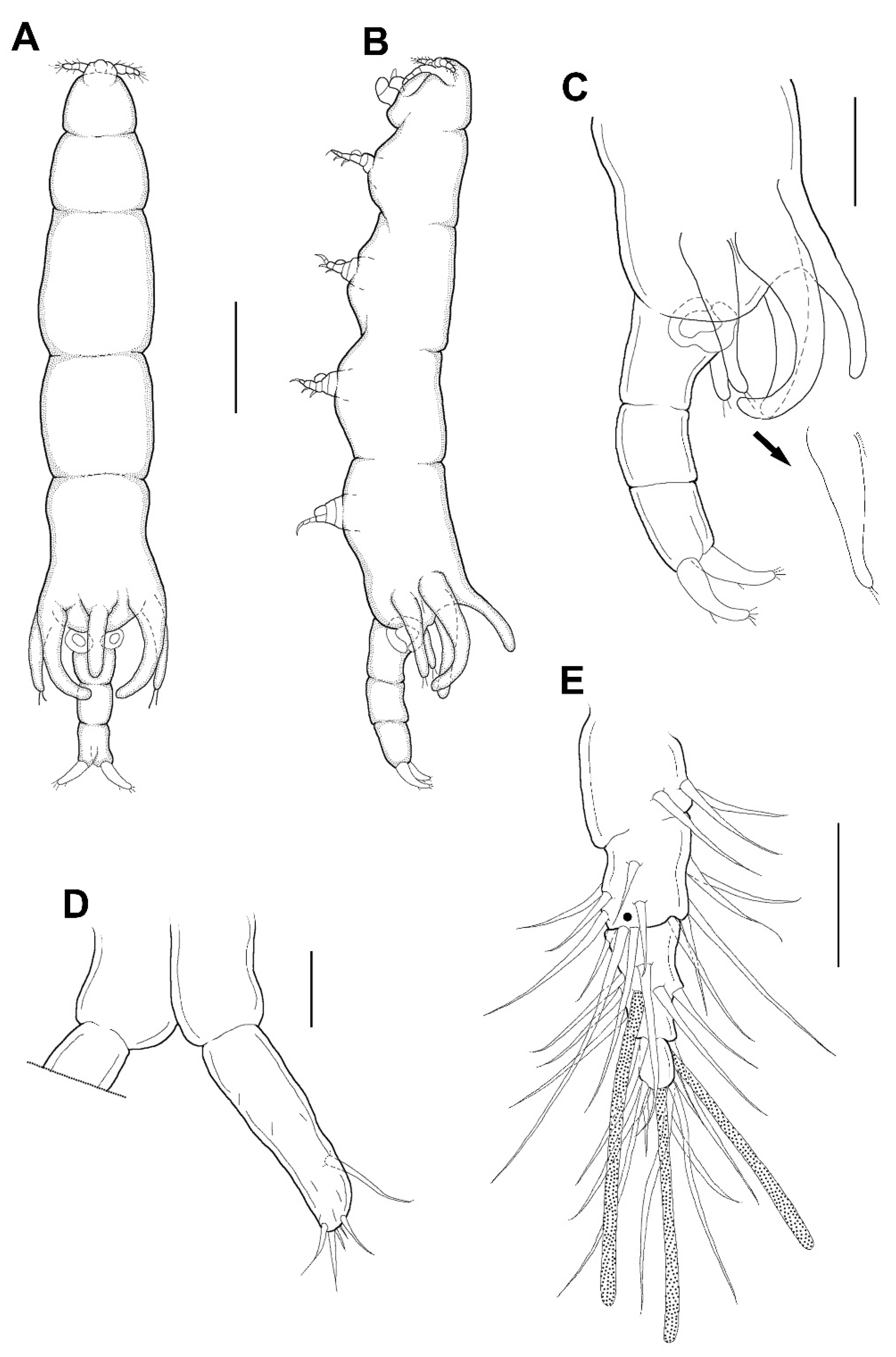
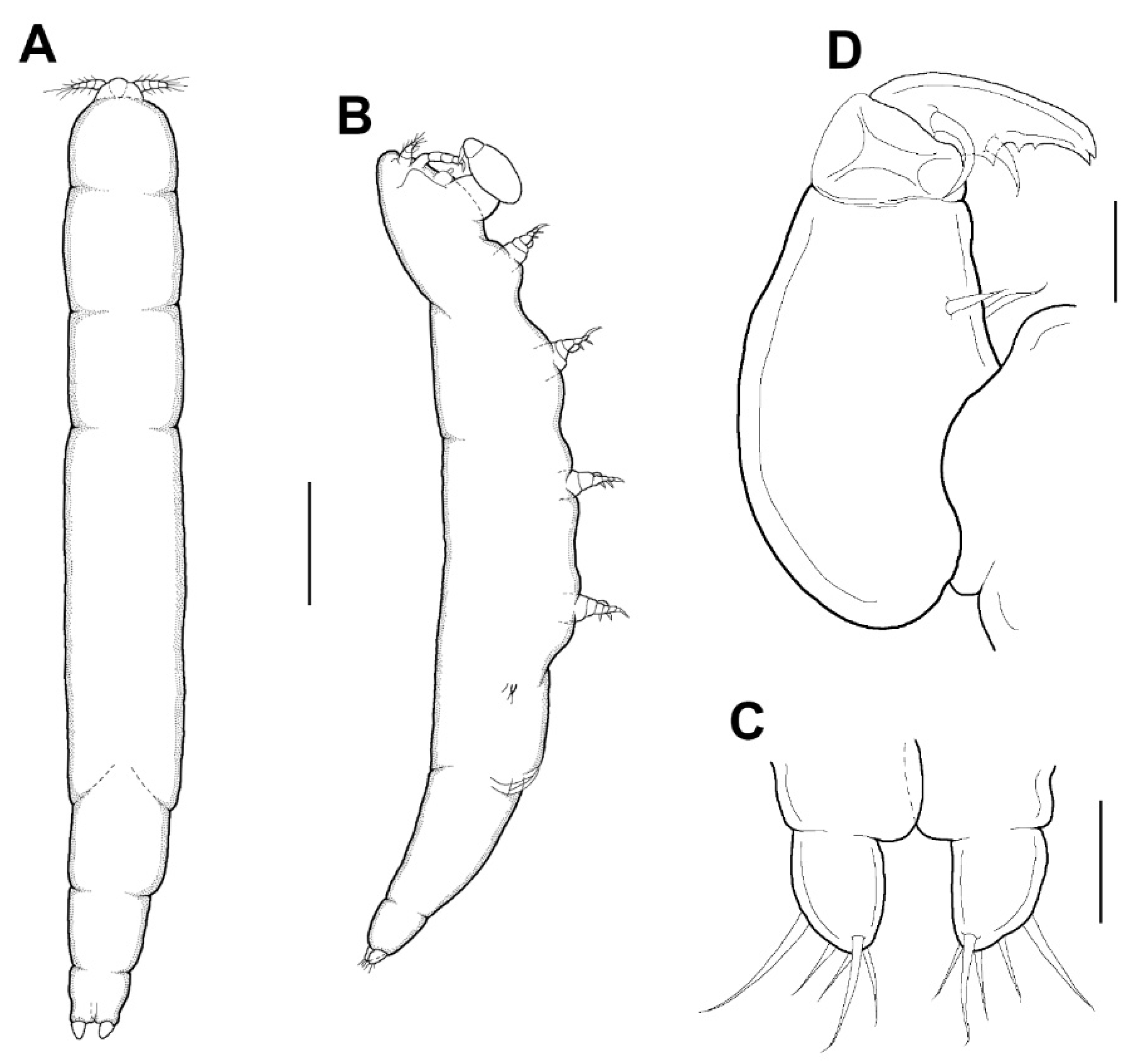
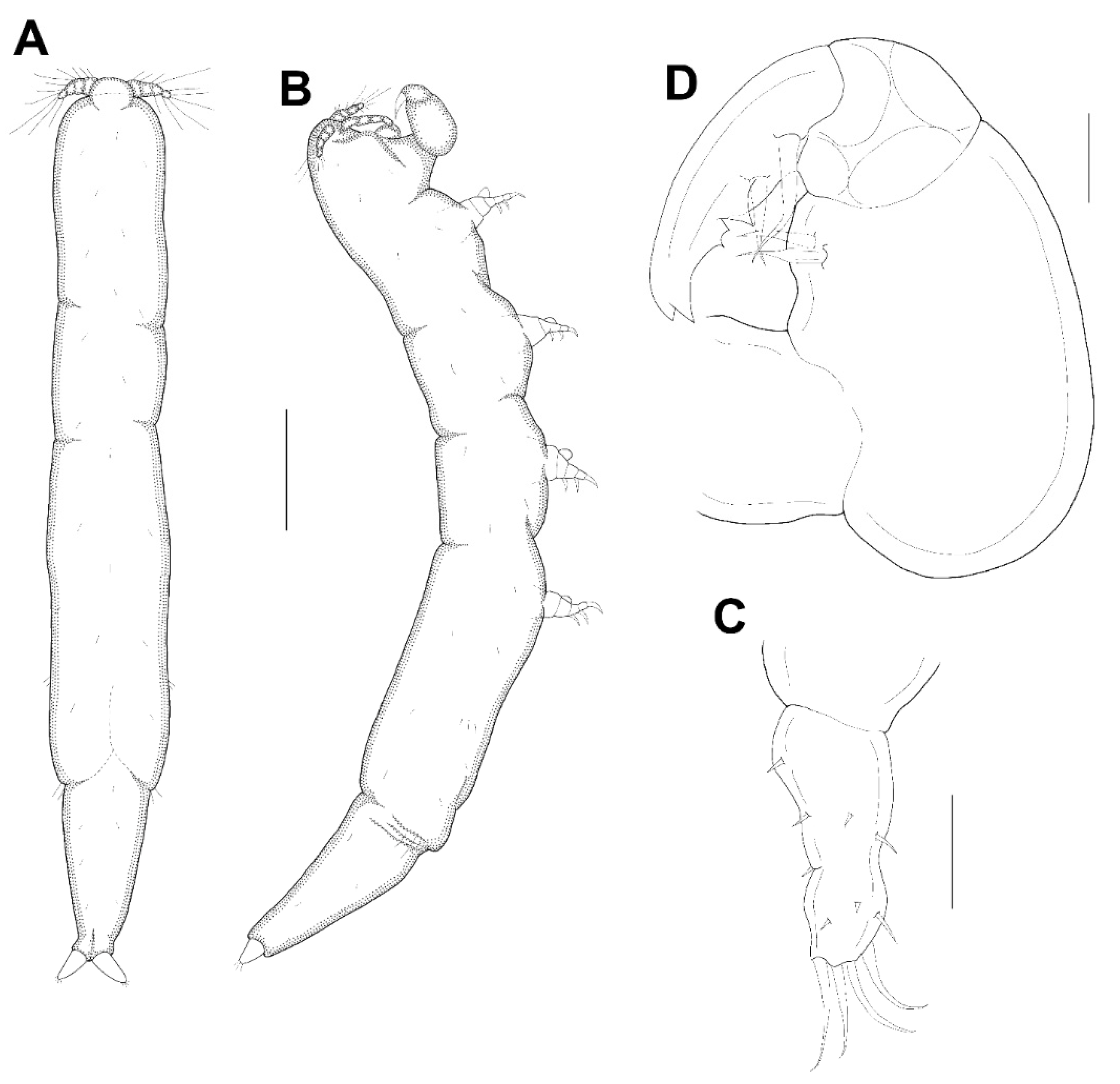
Female: Body (
Figure 3A,B) slender, about 6.16 times longer than wide. Length 1.17 (1.07–1.26) mm and greatest width 0.19 (0.17–0.21) mm based on two specimens in lactic acid before dissection. External segmentation not evident, but prosome with 4 lateral constrictions. Three unequal long posteriorly directed processes on the region dorsal to fifth legs, the middle one slightly smaller (
Figure 3A–C). Genital somite and abdomen (
Figure 3C) 3-segmented, straight or slightly curved dorsally, together occupying ~25% of body. Genital openings dorsolateral (
Figure 3A–C). Caudal ramus (
Figure 3D) elongate, covered with small setules, 107 µm long, 31 µm wide at the base, ratio 3.45:1, bearing 6 setae (1 outer lateral and 5 terminal setae). Body surface smooth (
Figure 3A–C).
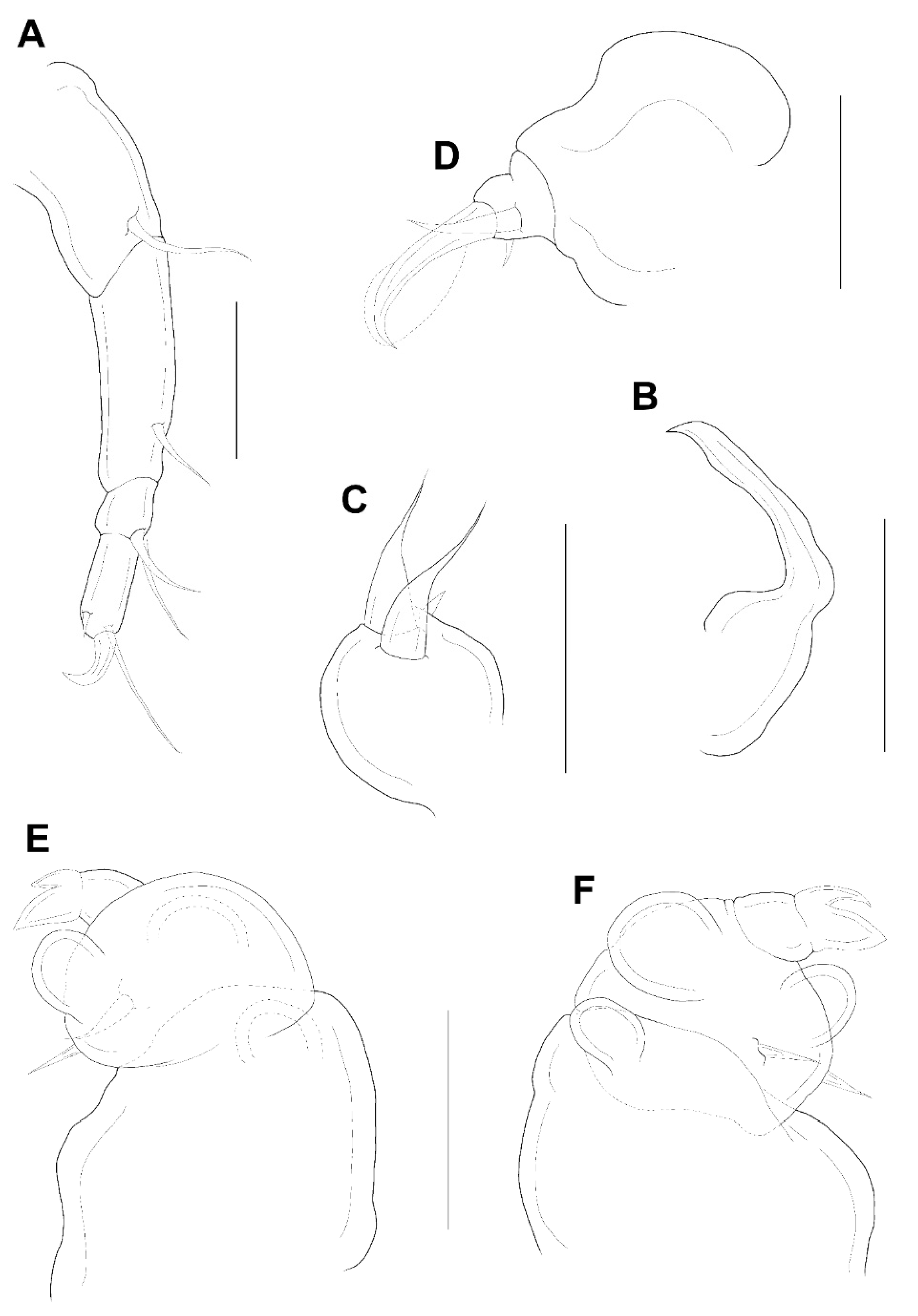
Antennule (
Figure 3E) 5-segmented, width of first to fifth segments: 23.3 × 18.7, 18.7 × 15.6, 12.5 × 10.9, 7.8 × 7.8, and 9.4 × 6.2 μm, respectively; armature formula: 3, 13, 5 + 1 aesthetasc, 2 + 1 aesthetasc, and 7 + 1 aesthetasc from proximal to distal; all setae naked. Antenna (
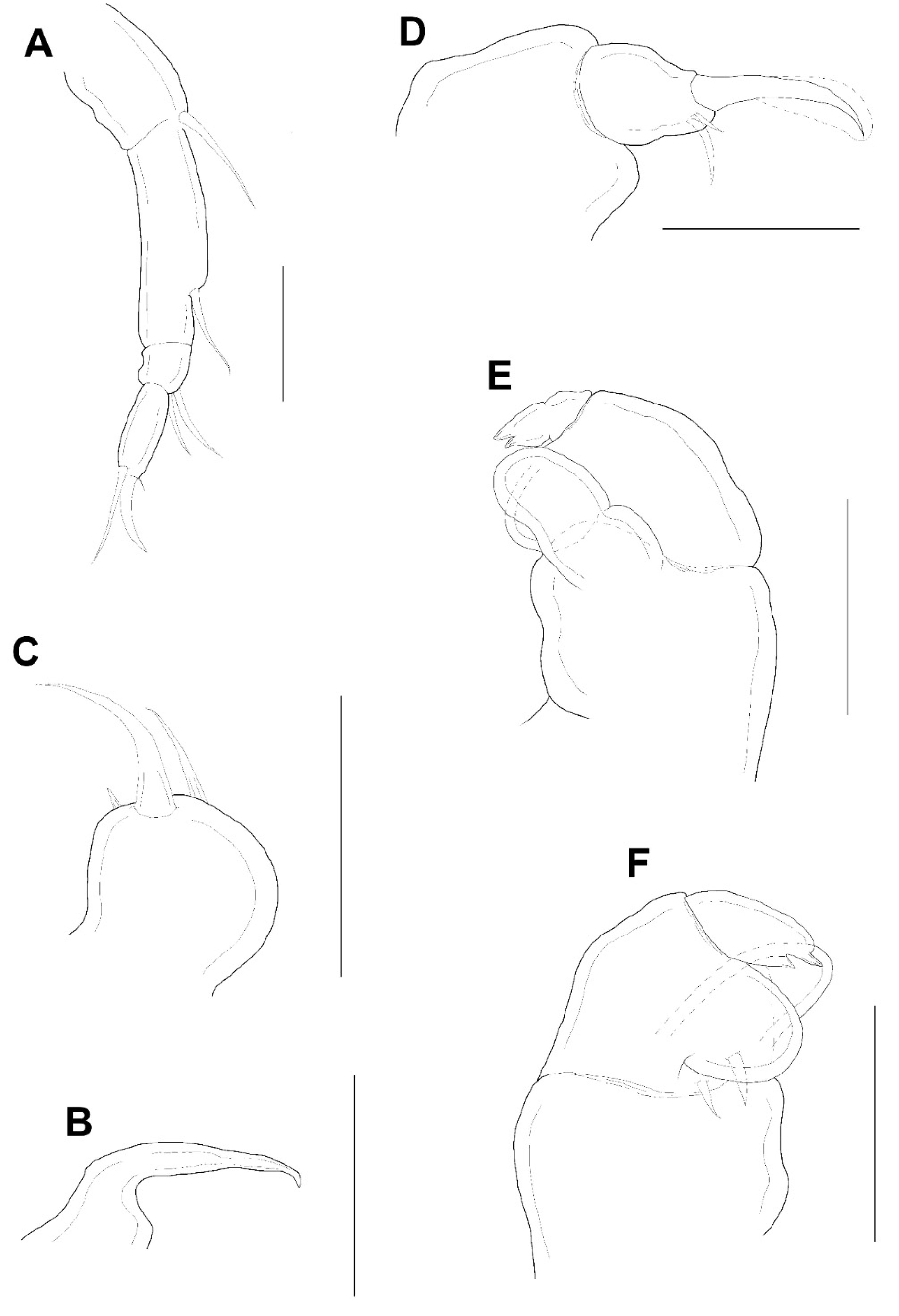
Figure 4A) 4-segmented. Armature formula 1, 1, 2, 2+claw from proximal to distal; terminal claw slightly shorter than final segment. Mandible (
Figure 4B) simple, slender, and smooth. Maxillule (
Figure 4C) with 2 terminal setae and 1 small anterior spiniform process. Maxilla (
Figure 4D) 2-segmented; first segment (syncoxa) unarmed; second segment (basis) drawn out into pointed process having large, slightly curved, claw-like seta with lamella, and bearing with two unequal inner setae. Maxilliped (
Figure 4E,F) 3-segmented; syncoxa (first segment) with large, distal protuberance (the length as the second segment of maxilliped); basis (second segment) having lateral lobe, 2 medial setae proximally; endopod (third segment) small, tipped with 2 spiniform setae.
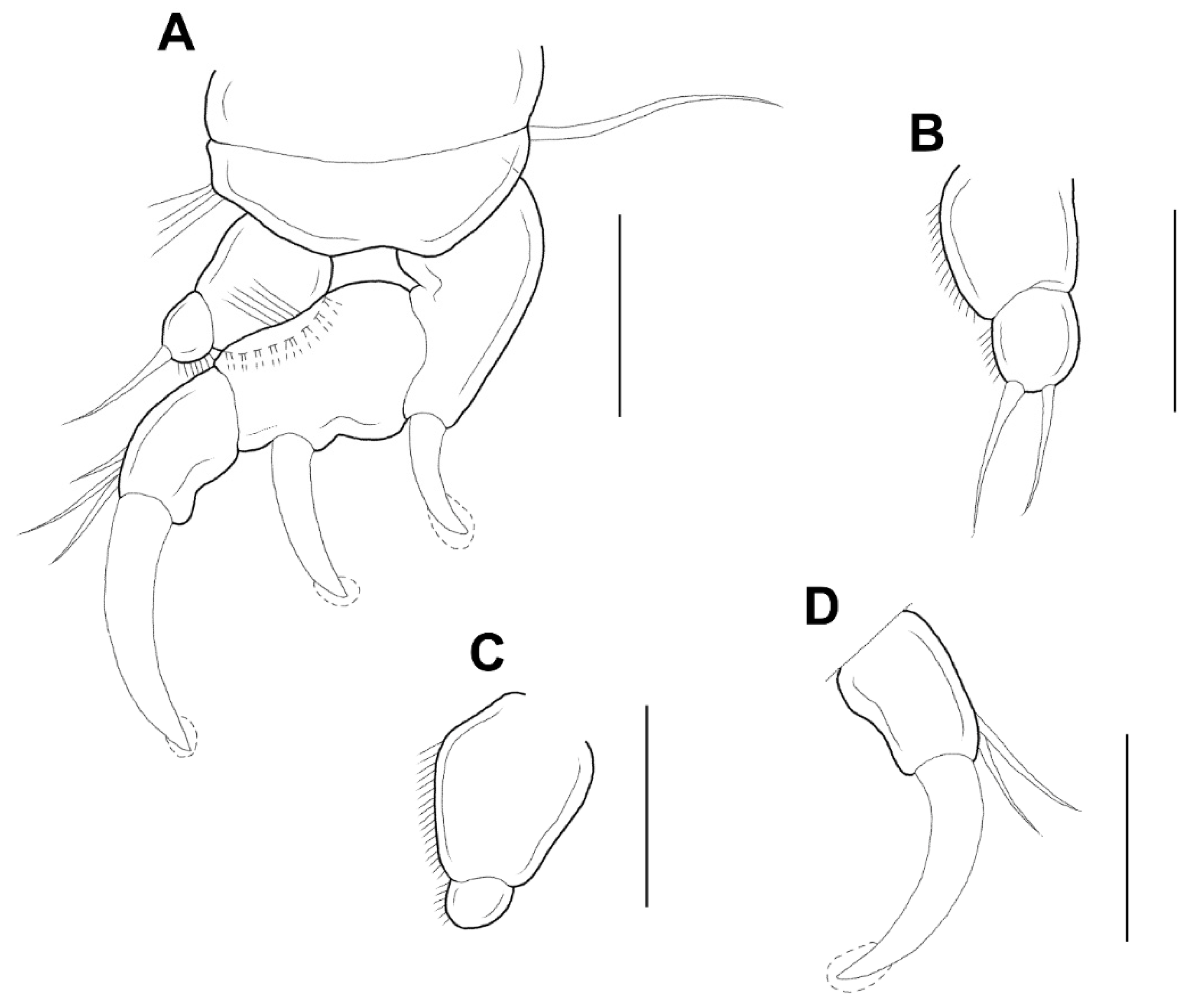
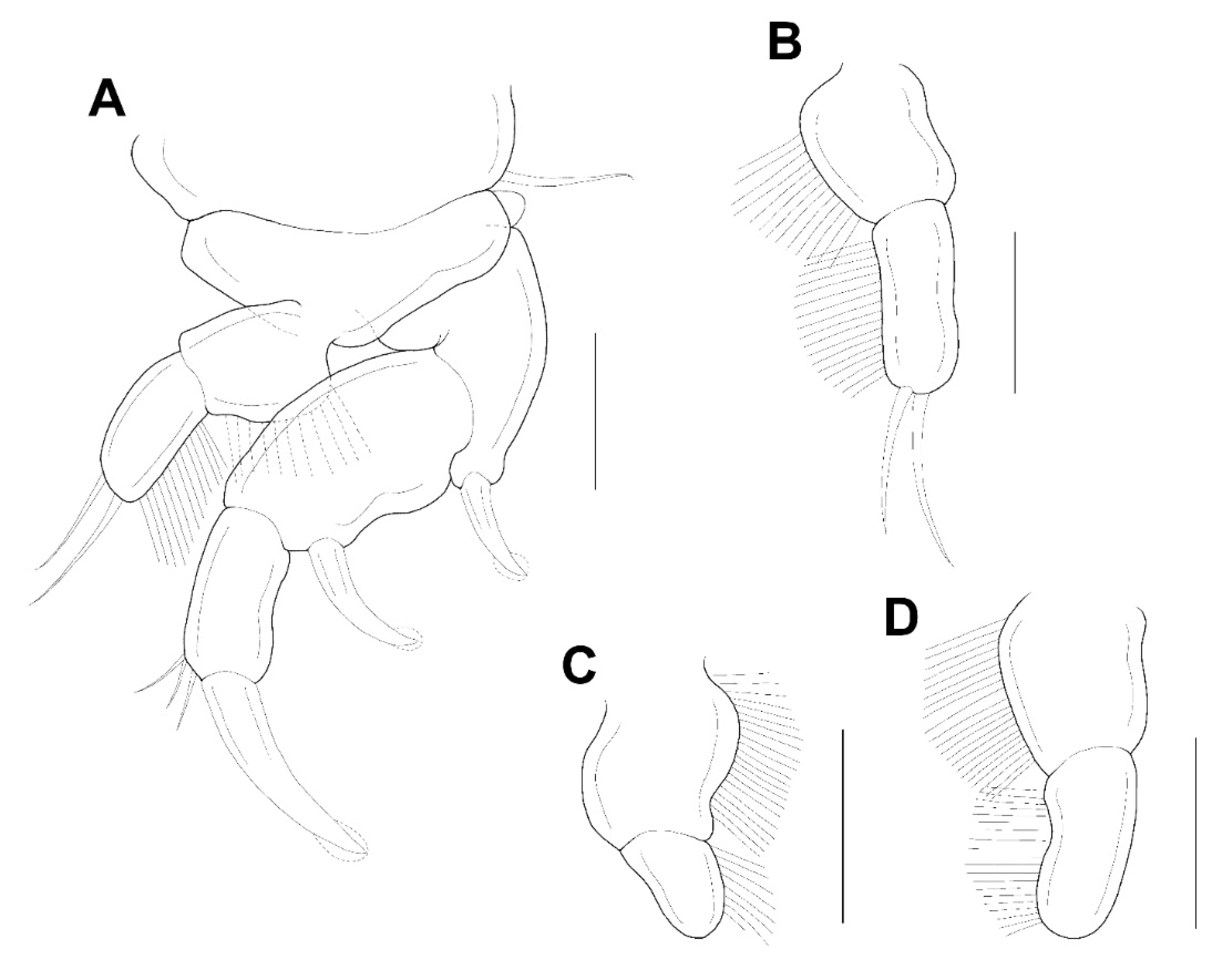
Legs 1–4 (
Figure 5A–D) each with 3-segmented exopod, 2-segmented endopod. The shape and armature of leg 2 as in leg 1, but endopod of leg 2 armed with 2 setae (instead of 1 seta). Exopods of legs 3 and 4 similar to legs 1 and 2, but 2 setae only (instead of 3 setae) on the terminal segment. Endopod of leg 3 unarmed. Endopod of leg 4 bearing with 1 seta. Formula of spines (Roman numerals) and setae (Arabic numerals) as follows:
| | Coxa | Basis | Exopod | Endopod |
| Leg 1 | 0-0 | 1-0 | I-0; I-0; I, 3 | 0-0; 1 |
| Leg 2 | 0-0 | 1-0 | I-0; I-0; I, 3 | 0-0; 2 |
| Leg 3 | 0-0 | 1-0 | I-0; I-0; I, 2 | 0-0; 0 |
| Leg 4 | 0-0 | 1-0 | I-0; I-0; I, 2 | 0-0; 1 |
Leg 5 (
Figure 3A–C) 155.4 μm long, 44.4 μm wide at base, tapering distally, armed with single proximal seta and 2 apical setae.
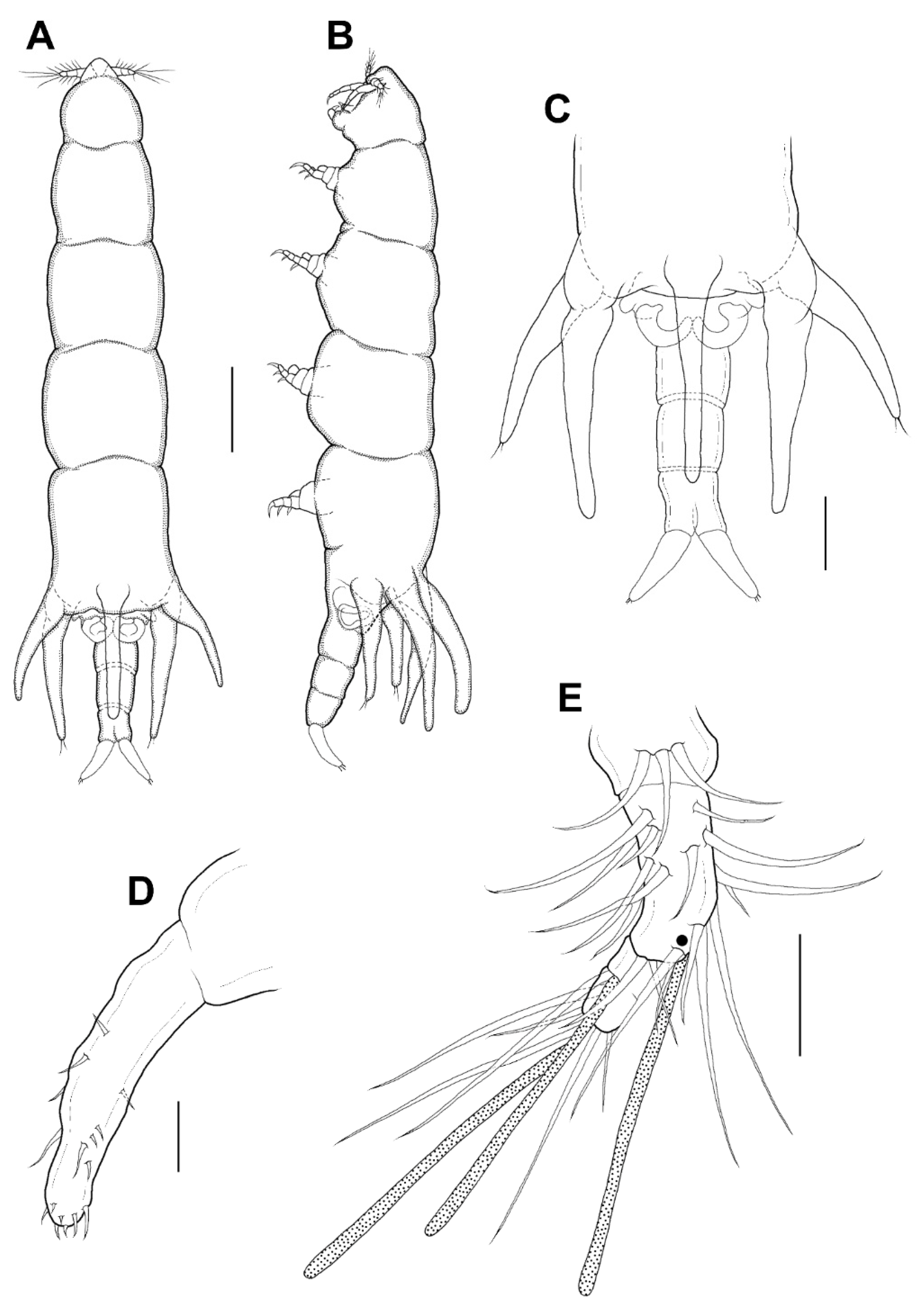
Male: Body (
Figure 6A,B) slender than female, about 8.6 times longer than wide. Length 1.38 (1.23–1.52) mm and greatest width 0.16 (0.15–0.16) mm based on two specimens in lactic acid before dissection. Caudal ramus (
Figure 6C) small, about 25.0 × 19.1 μm, as in female, bearing 5 setae. Body surface smooth.
The appendages including antennule, antenna, mandible, maxillule, and maxilla resembling that of female, but antennule with additional aesthetasc indicated by a dot on second segment (
Figure 3E). Maxilliped (
Figure 6D) 4-segmented; syncoxa (first segment) and first endopodal segment (third segment) unarmed; basis (second segment) with 2 medial setae; second endopodal segment (fourth segment) a claw with bifurcate tip, 2 proximal setae at base; concave margin with 3 serrations.
Legs 1–4 resembling those of female; leg 5 (
Figure 6B) minute, with 2 terminal setae, dorsal seta adjacent to the base. Leg 6 (
Figure 6B) as 2 small setae on genital operculum of genital somite.
Spermatophore not observed.
Etymology: The species is named after the type-locality Yanliao.
Material examined: Four specimens (two ♀♀ and two ♂♂) were obtained from washings of a
Psammocora columna colony collected at 5 m depth, at Longdon (25°06′49.5″ N, 121°55′11.8″ E), Gongliao District, New Taipei, Taiwan on 12 August 2010 (
Figure 1 and
Figure 2). Female holotype (NTUM-Inv-10021) and male allotype (NTUM-Inv-10022) were deposited in National Taiwan University Museum (NTUM), Taipei, Taiwan.
Female: Body (
Figure 7A,B) moderately stout. Length1.45 (1.33–1.58) mm and greatest width 0.27 (0.25–0.29) mm based on two specimens in lactic acid before dissection. Ration of body length to greatest width 5.4:1. External segmentation distinct, prosome having 4 lateral constrictions. Region dorsal to fifth legs with 3 equal long posteriorly processes (
Figure 7A–C). Genital somite and abdomen (
Figure 7C) 3-segmented, straight, together occupying ~20% of body length. Genital openings dorsolateral (
Figure 7A–C). Egg sacs not observed. Caudal ramus (
Figure 7D) elongate, 107.4 × 27.9 µm, bearing with 4 or 6 setae. Body surface smooth (
Figure 7A–C).
Antennule (
Figure 7E) 4-segmented, armature: 3, 18 + 1 aesthetasc, 2 + 1 aesthetasc, 4 + 1 aesthetasc; all setae naked. Antenna (
Figure 8A) 4-segmented, armature: 1, 1, 2, 2+claw; terminal claw small, about 1/2 length of terminal (fourth) segment. Mandible (
Figure 8B) with smooth blade. Maxillule (
Figure 8C) with 3 setae distally, including one small anterior spiniform process and two relatively long middle setae. Maxilla (
Figure 8D) 2-segmented; similar to that of
X. yanliaoensis sp. nov., but second segment (basis) slightly small and bearing with relatively longer outer seta. Maxilliped (
Figure 8E,F) 3-segmented; syncoxa (first segment) with a small, distal protuberance; basis (second segment) with 2 lateral lobes, 2 medial setae proximally; endopod (third segment) much small, bearing with 2 elements (the larger one equal to the length of third segment).
Legs 1–4 (
Figure 9A–D) each with 3-segmented exopod, 2-segmented endopod. The shape and armature of leg 2 as in leg 1. Exopods of legs 3 and 4 similar to legs 1 and 2, but their third segment armed with 2 setae and 1 seta, respectively. Endopod of legs 3 and 4 unarmed. Formula of spines (Roman numerals) and setae (Arabic numerals) as follows:
| | Coxa | Basis | Exopod | Endopod |
| Leg 1 | 0-0 | 1-0 | I-0; I-0; I, 3 | 0-0; 2 |
| Leg 2 | 0-0 | 1-0 | I-0; I-0; I, 3 | 0-0; 2 |
| Leg 3 | 0-0 | 1-0 | I-0; I-0; I, 2 | 0-0; 0 |
| Leg 4 | 0-0 | 1-0 | I-0; I-0; I, 1 | 0-0; 0 |
Leg 5 (
Figure 7A–C) as that in
X. yanliaoensis sp. nov., but slightly stout, about 275.5 μm long and 75.5 μm wide at base.
Male: Body (
Figure 10A,B) slender than female, about 8.2 times longer than wide. Length 1.55 (1.53–1.57) mm and greatest width 0.19 (0.16–0.21) mm based on two specimens in lactic acid before dissection. Abdomen (
Figure 10A) segmentation indistinct. Caudal ramus (
Figure 10C) small, about 55.8 × 27.9 μm, ornamented with several setules, bearing with 4 terminal setae (lateral seta not clearly evident). Body surface covered with several setules (
Figure 10A,B).
The appendages including antennule, antenna, mandible, maxillule, and maxilla as that of female, but antennule with additional aesthetasc indicated by a dot on second segment (
Figure 7E). Maxilliped (
Figure 10D) 4-segmented; syncoxa (first segment) and first endopodal segment (third segment) unarmed; basis (second segment) largest, with 2 medial setae (one normal and one modified with sclerotized base and hyaline attenuate distal part); second endopodal segment (fourth segment) a claw with bifurcate tip, bearing with 2 proximal setae equally; concave margin with 3 serrations.
Legs 1–4 as in female; leg 5 (
Figure 10B) consisting only of 3 setae. Leg 6 (
Figure 10A,B) represented by 2 small setae on the genital operculum of genital somite.
Spermatophore not observed.
Etymology: The specific name magnifica, Latin adjective for “big” or “majestic” refers to the relatively strong element on the third segment of maxilliped.
4. Discussion
Based on the findings from this study, the genus
Xarifia currently consists of 94 species and eight of them live in association with the
Psammocora coral. The armature of endopods of legs 1–4 is the main diagnostic feature of
Xarifia. The terminal segment of endopods of legs 1–4 of
X.
yanliaoensis sp. nov. is a unique character, armed with 1, 2, 0, 1 setae, respectively. After comparison with another valid species of
Xarifia, we recognized that only
Xarifia simplex Humes, 1985 [
4] parasitic in the
Scapophyllia cylindrica (=
Merulina cylindrica (Milne Edwards and Haime, 1849) [
17]) present the same armature of endopods of legs 1–4. However, the body surface of
X.
simplex covered with scattered small hairs (setules), while the body surface of
X.
yanliaoensis sp. nov. is smooth. The
X.
yanliaoensis sp. nov. differs from
X.
simplex by (1) the armature formula of the maxilla: three setae in
X.
yanliaoensis sp. nov. but two in
X.
simplex [
4] (p. 561: Figure 49l); (2) the armature of terminal segment of antenna: I + 2 in
X.
yanliaoensis sp. nov., while I + 1 in
X.
simplex [
4] (p. 561: Figure 49h); (3) the size of lobe or protuberance on the second segment of maxilliped: the protuberance in
X.
yanliaoensis sp. nov. is obviously bigger than that in
X.
simplex [
4] (p. 562: Figure 50a,b) and its length is similar to the second segment of maxilliped. In addition, the maxilliped of male between these two species also showed some differences: with trifurcate tip and triangular process on the concave edge of the fourth segment in
X.
simplex [
4] (p. 562: Figure 50j), but with bifurcate tip and 3 serrations on concave margin in
X.
yanliaoensis sp. nov.
The other new species described herein,
X.
magnifica sp. nov., is similar to
Xarifia anopla Humes and Dojiri, 1982 [
18],
Xarifia brevicauda Humes and Ho, 1968 [
19],
Xarifia filata Humes, 1985 [
4],
Xarifia hadra Humes and Dojiri, 1983 [
20],
Xarifia scutipes Humes and Dojiri, 1983 [
20],
Xarifia longa Cheng, Ho and Dai, 2007 [
21],
Xarifia capillata Cheng, Ho and Dai, 2011 [
22], and
Xarifia parva Cheng, Ho and Dai, 2011 [
22] in the armature formula of the terminal endopodal segments of legs 1–4 (2, 2, 0, 0).
Xarifia anopla,
X.
filata,
X.
hadra,
X.
longa, and
X.
parva can be excluded first because of their endopods of legs 1-4 exhibit only 1 segment.
Xarifia capillata can be distinguished from
X.
magnifica sp. nov. by the region dorsal to fifth legs with a single central process which is tipped with tuft of setules [
22] (p. 228: Figure 1A–C).
Xarifia scutipes may be distinguished easily from
X.
magnifica sp. nov. in the shield-like leg 5 in the female [
20] (p. 282: Figure 18a–c) and the claw of the maxilliped in the male with a large hyaline excrescence on the concave margin [
20] (p. 285: Figure 21d,e).
Xarifia brevicauda differs from
X.
magnifica sp. nov. by its abbreviated genital somite and abdomen [
19] (p. 448: Figures 114–116), and by the serrated excrescence on the claw of the male maxilliped [
19] (p. 449: Figure 130).
Eight species of
Xarifia have been known using
Psammocora corals as hosts (
Table 1). We selected certain external features of
Xarifia that are useful for the determination of species. Eleven features are shown in
Table 2 and a useful key is provided as follows:
Key to females of eight species of Xarifia parasitic in Psammocora corals (modified from [12]). 1a. Region dorsal to fifth legs with processes or knobs…2
1b. Region dorsal to fifth legs smooth… X. lata
2a. Sides of genital segment smooth…3
2b. Sides of genital segment with small sclerotized lobe…X. imitans
3a. Maxillule with 3 elements…4
3a. Maxillule with 2 elements…6
4a. Maxilliped with 2 lobes…5
4b. Maxilliped with 3 lobes…X. magnifica sp. nov.
5a. Terminal armature on endopods of legs 1 (4 2, 3, 0, 1)…X. conrepta
5b. Terminal armature on endopods of legs 1 (4 1, 2, 0, 1)…X. yanliaoensis sp. nov.
6a. Maxilliped with more than 2 lobes…7
6b. Maxilliped with 2 lobes…X. diminuta
7a. Terminal armature on endopods of legs 1 (4 2, 3, 0, 1)…X. formosa
7b. Terminal armature on endopods of legs 1 (4 3, 3, 2, 2)…X. gracilis
Table 2.
Differences between the eight species of Xarifia copepods isolated from Psammocora corals.
Table 2.
Differences between the eight species of Xarifia copepods isolated from Psammocora corals.
| (a) | |
|---|
| Species | Characters * |
|---|
| 1 | 2 | 3 | 4 | 5 | 6 |
|---|
| X. diminuta | 0.98
(0.78–118) | 5.4:1 | 3 | 4 | 2, 2, 0, 2 | 3, 3, 2, 2 |
| X. formosa | 1.15
(1.10–1.19) | 5.2:1 | 3 | 5 | 2, 3, 0, 1 | 3, 3, 3, 2 |
| X. imitans | 0.97
(0.94–1.01) | 6.6:1 | 3 | 5 | 2, 3, 0, 1 | 3, 3, 3, 2 |
| X. conrepta | 0.80
(0.74–0.82) | 4.7:1 | 3 | 5 | 2, 3, 0, 1 | 3, 3, 2, 2 |
| X. gracilis | 0.78
(0.75–0.80) | 6.5:1 | 3 | 5 or 6 | 3, 3, 2, 2 | 3, 3, 2, 2 |
| X. lata | 1.05
(1.02–1.09) | 5.8:1 | 0 | 4 | 1, 2, 1, 1 | 2, 2, 2, 1 |
| X. magnifica sp. nov. | 1.45
(1.33–1.58) | 5.4:1 | 3 | 4 or 6 | 2, 2, 0, 0 | 3, 3, 2, 1 |
| X.yanliaoensis sp. nov. | 1.17
(1.07–1.26) | 6.2:1 | 3 | 6 | 1, 2, 0, 1 | 3, 3, 2, 2 |
| (b) | |
| Species | Characters * |
| 7 | 8 | 9 | 10 | 11 |
| X. diminuta | 2 | 2 | 2 setae | 2 lobes | smooth |
| X. formosa | 2 | 2 | 2 setae | 4 lobes | smooth |
| X. imitans | 2 | 2 | 2 setae | 2 lobes | with lateral process |
| X. conrepta | 2 | 2 | 3 setae | 2 lobes | smooth |
| X. gracilis | 2 | 2 | 2 setae | 5 lobes | smooth |
| X. lata | 1 or 2 | 1 or 2 | 2 setae | 3 lobes | smooth |
| X.magnifica sp. nov. | 2 | 2 | 3 setae | 3 lobes | smooth |
| X.yanliaoensis sp. nov. | 2 | 2 | 3 setae | 2 lobes | smooth |
We believe that the abundance and species composition of coral-associated copepods in the host corals could be considered as bioindicators for predicting the health condition of host corals. Furthermore, this could further be an assessment index for coral conservation. However, this assessment could be feasible only if the interactions and ecological impacts between parasitic copepods and host corals are well recognized. In the present study, we have improved the understanding of species diversity of coral-associated copepods in the world. As accumulating knowledge of parasitic copepod and host coral relationships, we expect to reduce the stress of harms and diseases to host corals by applying biological control of parasitic copepods in the future.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









