
Organic Selenium (OH-MetSe) Effect on Whole Body Fatty Acids and Mx Gene Expression against Viral Infection in Gilthead Seabream (Sparus aurata) Juveniles

 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Feeding Trial
2.2. Experimental Diets
2.3. Selenium Analysis
2.4. Growth Performance
2.5. Biochemical Composition
2.5.1. Proximate Composition
2.5.2. Fatty Acid Profiles
2.5.3. Fatty Acid Retention Efficiency
- a: (weight (g) × body lipid (%) × FA (%) in whole body)/100
- b: (Initial weight (g) × initial body lipid (%) × FA (%) in initial whole body)/100
- c: Feed intake (g) × (dietary lipid (%) × FA (%) in diet/100) × 100
2.6. Crowding Stress Challenge
2.7. Anti-Viral Response Challenge
2.8. Statistical Analysis
3. Results
3.1. Selenium Analysis
3.2. Growth Performance
3.3. Biochemical Analysis
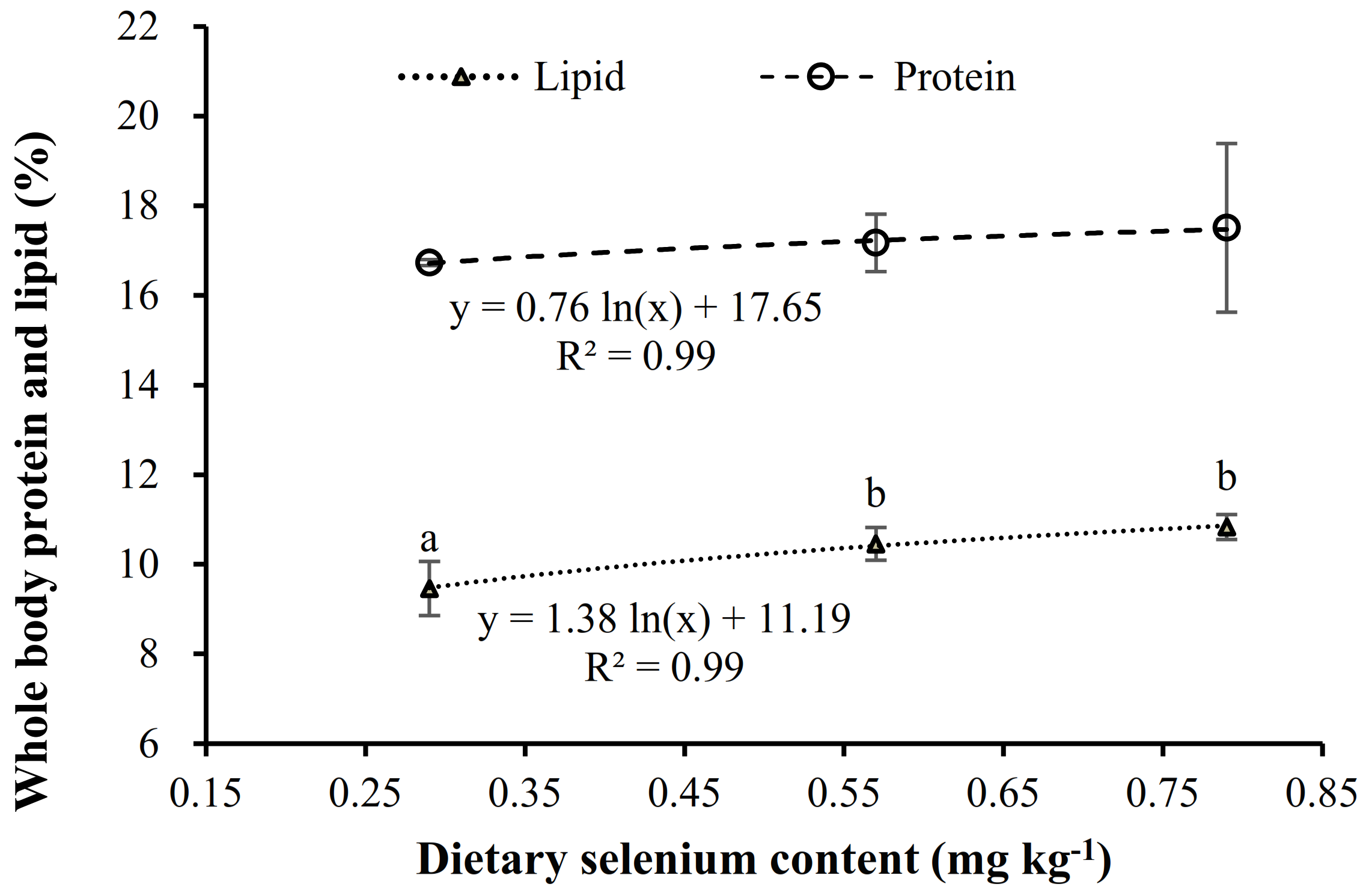
3.3.1. Proximate Composition
3.3.2. Fatty Acid Profiles
3.3.3. Fatty Acid Retention Efficiency
3.4. Crowding Stress Challenge
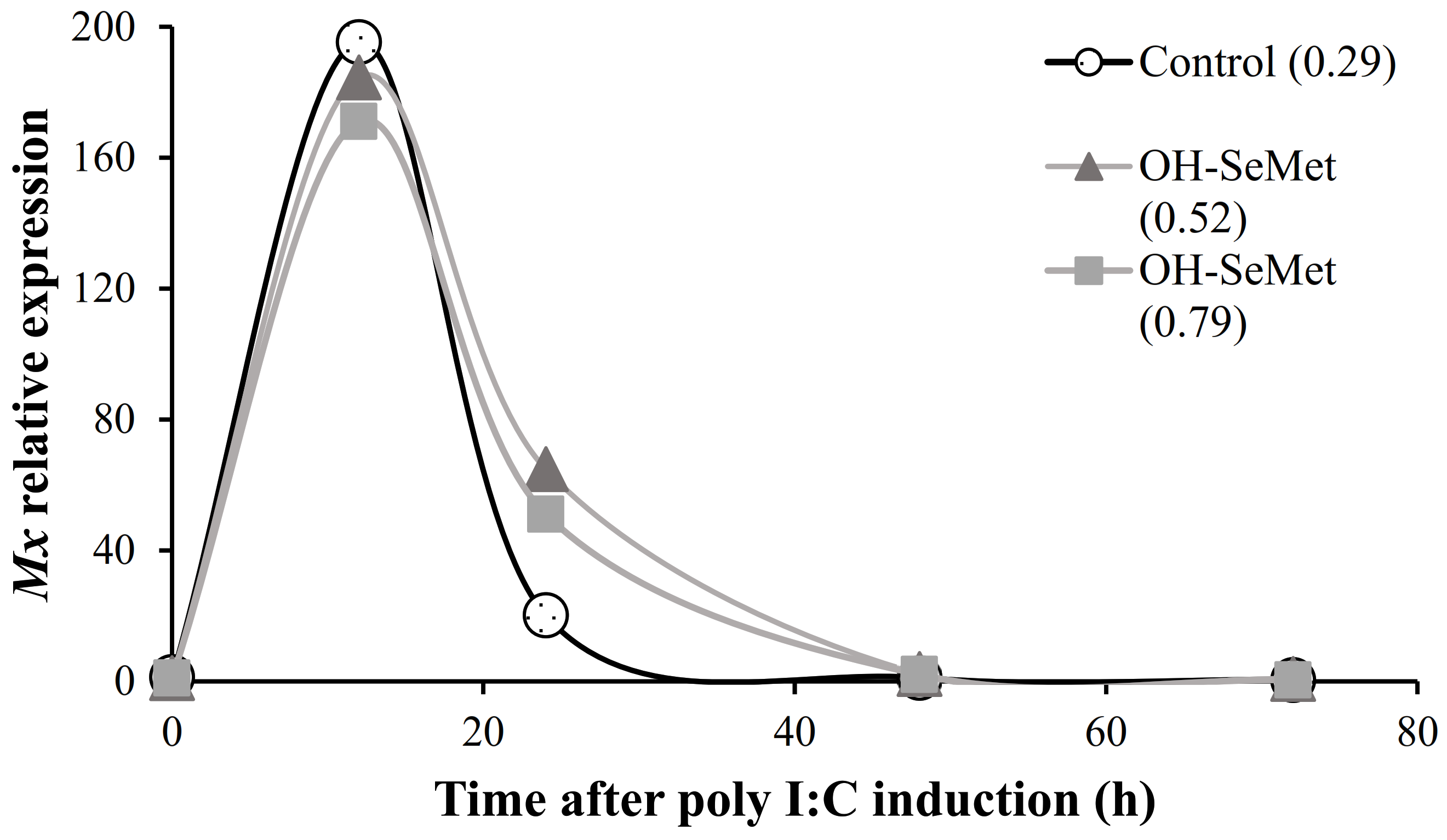
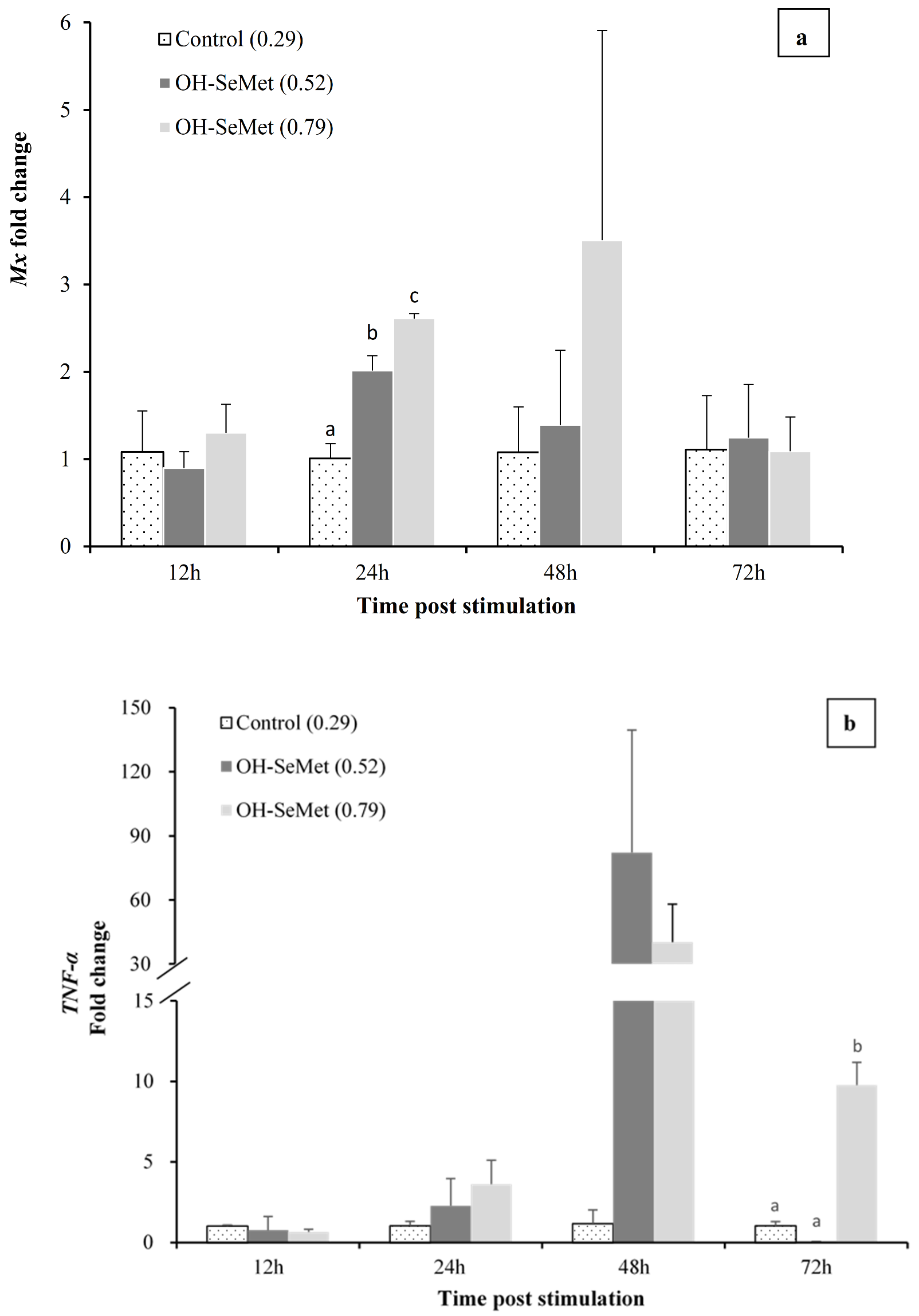
3.5. Anti-Viral Response Challenge
4. Discussion
4.1. Effect of Dietary Se in Growth Performance
4.2. Effect of Dietary Se in Whole-Body Composition and Se Content in Tissues
4.3. Effect of Dietary Se in Whole-Body FA’s Composition and Retention
4.4. Effect of Dietary Se in Plasma Cortisol
4.5. Effect of Dietary Se in Gene Expression of Mx Gene and Immune Related Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NRC. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011; ISBN 0309216192. [Google Scholar]
- Tripathy, A. Oxidative Stress, Reactive Oxygen Species (ROS) and Antioxidative Defense System, with Special Reference to Fish. Int. J. Curr. Res. Biosci. Plant Biol. 2016, 3, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-H.; Shiau, S.-Y. Dietary selenium requirements of juvenile grouper, Epinephelus malabaricus. Aquaculture 2005, 250, 356–363. [Google Scholar] [CrossRef]
- Atencio, L.; Moreno, I.; Jos, A.; Prieto, A.I.; Moyano, R.; Blanco, A.; Cameán, A.M. Effects of dietary selenium on the oxidative stress and pathological changes in tilapia (Oreochromis niloticus) exposed to a microcystin-producing cyanobacterial water bloom. Toxicon 2009, 53, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Wang, X.J.; Ai, Q.; Mai, K.; Zhang, W. Dietary selenium requirement for juvenile cobia, Rachycentron canadum L. Aquac. Res. 2010, 41, e594–e601. [Google Scholar] [CrossRef]
- Le, K.T.; Fotedar, R. Dietary selenium requirement of yellowtail kingfish (Seriola lalandi). Agric. Sci. 2013, 04, 68–75. [Google Scholar] [CrossRef]
- Domínguez, D.; Sehnine, Z.; Castro, P.; Robaina, L.; Fontanillas, R.; Prabhu, P.A.J.; Izquierdo, M. Optimum selenium levels in diets high in plant-based feedstuffs for gilthead sea bream (Sparus aurata) fingerlings. Aquac. Nutr. 2020, 26, 579–589. [Google Scholar] [CrossRef]
- Betancor, M.B.; Caballero, M.; Terova, G.; Saleh, R.; Atalah, E.; Benítez-Santana, T.; Bell, J.G.; Izquierdo, M. Selenium inclusion decreases oxidative stress indicators and muscle injuries in sea bass larvae fed high-DHA microdiets. Br. J. Nutr. 2012, 108, 2115–2128. [Google Scholar] [CrossRef] [Green Version]
- Saleh, R.; Betancor, M.B.; Roo, J.; Montero, D.; Zamorano, M.J.; Izquierdo, M. Selenium levels in early weaning diets for gilthead seabream larvae. Aquaculture 2014, 426, 256–263. [Google Scholar] [CrossRef]
- Plateau, P.; Saveanu, C.; Lestini, R.; Dauplais, M.; Decourty, L.; Jacquier, A.; Blanquet, S.; Lazard, M. Exposure to selenomethionine causes selenocysteine misincorporation and protein aggregation in Saccharomyces cerevisiae. Sci. Rep. 2017, 7, 44761. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, M.; Koven, W. Lipids. In Larval Fish Nutrition; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 47–81. [Google Scholar] [CrossRef]
- Tocher, D.R. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Carvalho, M.; Montero, D.; Gesto, M.; Lencina, A.; Lund, I.; Izquierdo, M. The effect of dietary n-3 LC-PUFA on the responses to acute and prolonged stress of meagre (Argyrosomus regius, Asso 1801) juveniles. Aquaculture 2019, 506, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Ganga, R.; Montero, D.; Bell, J.G.; Atalah, E.; Ganuza, E.; Vega-Orellana, O.; Tort, L.; Acerete, L.; Afonso, J.M.; Benitez-Sanatana, T.; et al. Stress response in sea bream (Sparus aurata) held under crowded conditions and fed diets containing linseed and/or soybean oil. Aquaculture 2011, 311, 215–223. [Google Scholar] [CrossRef]
- Montero, D.; Izquierdo, M. Welfare and Health of Fish Fed Vegetable Oil. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; CRC Press: Boca Raton, FL, USA, 2010; pp. 439–485. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research. Am. J. Clin. Nutr. 2010, 92, 1002. [Google Scholar] [CrossRef] [Green Version]
- Mansour, A.; Goda, A.; Omar, E.A.; Khalil, H.S.; Esteban, M. Dietary supplementation of organic selenium improves growth, survival, antioxidant and immune status of meagre, Argyrosomus regius, juveniles. Fish Shellfish Immunol. 2017, 68, 516–524. [Google Scholar] [CrossRef]
- Fontagné-Dicharry, S.; Véron, V.; Larroquet, L.; Godin, S.; Wischhusen, P.; Aguirre, P.; Terrier, F.; Richard, N.; Bueno, M.; Bouyssière, B.; et al. Effect of selenium sources in plant-based diets on antioxidant status and oxidative stress-related parameters in rainbow trout juveniles under chronic stress exposure. Aquaculture 2020, 529, 735684. [Google Scholar] [CrossRef]
- Domínguez, D.; Rimoldi, S.; Robaina, L.E.; Torrecillas, S.; Terova, G.; Zamorano, M.J.; Karalazos, V.; Hamre, K.; Izquierdo, M. Inorganic, organic, and encapsulated minerals in vegetable meal based diets forSparus aurata (Linnaeus, 1758). PeerJ 2017, 5, e3710. [Google Scholar] [CrossRef] [Green Version]
- Mechlaoui, M.; Dominguez, D.; Robaina, L.; Geraert, P.-A.; Kaushik, S.; Saleh, R.; Briens, M.; Montero, D.; Izquierdo, M. Effects of different dietary selenium sources on growth performance, liver and muscle composition, antioxidant status, stress response and expression of related genes in gilthead seabream (Sparus aurata). Aquaculture 2019, 507, 251–259. [Google Scholar] [CrossRef]
- Küçükbay, F.; Yazlak, H.; Karaca, I.; Sahin, N.; Tuzcu, M.; Cakmak, M.; Sahin, K. The effects of dietary organic or inorganic selenium in rainbow trout (Oncorhynchus mykiss) under crowding conditions. Aquac. Nutr. 2009, 15, 569–576. [Google Scholar] [CrossRef]
- Pérez-Valenzuela, J.; Mejías, M.; Ortiz, D.; Salgado, P.; Montt, L.; Chávez-Báez, I.; Vera-Tamargo, F.; Mandakovic, D.; Wacyk, J.; Pulgar, R. Increased dietary availability of selenium in rainbow trout (Oncorhynchus mykiss) improves its plasma antioxidant capacity and resistance to infection with Piscirickettsia salmonis. Vet. Res. 2021, 52, 1–14. [Google Scholar] [CrossRef]
- Neamat-Allah, A.N.; Mahmoud, E.; El Hakim, Y.A. Efficacy of dietary Nano-selenium on growth, immune response, antioxidant, transcriptomic profile and resistance of Nile tilapia, Oreochromis niloticus against Streptococcus iniae infection. Fish Shellfish Immunol. 2019, 94, 280–287. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. The Function of Fish Cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Secombes, C.; Hardie, L.; Daniels, G. Cytokines in fish: An update. Fish Shellfish Immunol. 1996, 6, 291–304. [Google Scholar] [CrossRef]
- Bravo, J.; Acosta, F.; Padilla, D.; Grasso, V.; Real, F. Mx expression in gilthead sea bream (Sparus aurata L.) in response to poly I:C, bacterial LPS and chromosomal DNA: Preliminary study. Fish Shellfish Immunol. 2011, 31, 170–172. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific Opinion on safety and efficacy of selenium in the form of organic compounds produced by the selenium?enriched yeast Saccharomyces cerevisiae NCYC R646 (Selemax 1000/2000) as feed additive for all species. EFSA J. 2012, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific Opinion on safety and efficacy of hydroxy-analogue of selenomethionine as feed additive for all species. EFSA J. 2013, 11, 1–30. [Google Scholar] [CrossRef] [Green Version]
- AOAC International; Cunniff, P. (Eds.) Official Methods of Analysis of AOAC INTERNATIONAL, 16th ed.; AOAC International: Arlington, VA, USA, 1995; ISBN 0935584544/9780935584547. Available online: https://ifremer.on.worldcat.org/oclc/300016697 (accessed on 21 September 2021).
- Floch, J. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Christie, W.W. Gas Chromatography and Lipids: A Practical Guide; The Oily Press Ltd.: Buckinghamshire, UK, 1989; 480p. [Google Scholar]
- Izquierdo, M.S. Optimal EFA levels in Artemia to meet the EFA requirements of red seabream (Pagrus major). In Proceedings of the 3th International Symposium on Feeding and Nutrition in Fish, Toba, Japan, 28 August–1 September 1989; pp. 221–232. [Google Scholar]
- Grasso, V.; Padilla, D.; Bravo, J.; Román, L.; Rosario, I.; Acosta, B.; Vega, B.; El Aamri, F.; Escuela, O.; Ramos-Vivas, J.; et al. Immunization of sea bream (Sparus aurata) juveniles against Photobacterium damselae subsp. piscicida by short bath: Effect on some pro-inflammatory molecules and the Mx gene expression. Fish Shellfish Immunol. 2015, 46, 292–296. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Morcillo, P.; Meseguer, J.; Esteban, M.; Cuesta, A. In vitro effects of metals on isolated head-kidney and blood leucocytes of the teleost fish Sparus aurata L. and Dicentrarchus labrax L. Fish Shellfish Immunol. 2016, 54, 77–85. [Google Scholar] [CrossRef]
- Castellana, B.; Iliev, D.B.; Sepulcre, M.P.; MacKenzie, S.; Goetz, F.W.; Mulero, V.; Planas, J.V. Molecular characterization of interleukin-6 in the gilthead seabream (Sparus aurata). Mol. Immunol. 2008, 45, 3363–3370. [Google Scholar] [CrossRef]
- Pellizzari, C.; Krasnov, A.; Afanasyev, S.; Vitulo, N.; Franch, R.; Pegolo, S.; Patarnello, T.; Bargelloni, L. High mortality of juvenile gilthead sea bream (Sparus aurata) from photobacteriosis is associated with alternative macrophage activation and anti-inflammatory response: Results of gene expression profiling of early responses in the head kidney. Fish Shellfish Immunol. 2013, 34, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Minganti, V.; Drava, G.; De Pellegrini, R.; Siccardi, C. Trace elements in farmed and wild gilthead seabream, Sparus aurata. Mar. Pollut. Bull. 2010, 60, 2022–2025. [Google Scholar] [CrossRef] [PubMed]
- Lall, S.P. The minerals. In Fish Nutrition; Elsevier: Amsterdam, The Netherlands, 2003; pp. 259–308. [Google Scholar]
- Wang, L.; Xiao, J.-X.; Hua, Y.; Xiang, X.-W.; Zhou, Y.-F.; Ye, L.; Shao, Q.-J. Effects of dietary selenium polysaccharide on growth performance, oxidative stress and tissue selenium accumulation of juvenile black sea bream, Acanthopagrus schlegelii. Aquaculture 2019, 503, 389–395. [Google Scholar] [CrossRef]
- Prabhu, P.A.J.; Holen, E.; Espe, M.; Silva, M.S.; Holme, M.-H.; Hamre, K.; Lock, E.-J.; Waagbø, R. Dietary selenium required to achieve body homeostasis and attenuate pro-inflammatory responses in Atlantic salmon post-smolt exceeds the present EU legal limit. Aquaculture 2020, 526, 735413. [Google Scholar] [CrossRef]
- Fontagné-Dicharry, S.; Godin, S.; Liu, H.; Prabhu, P.A.J.; Bouyssiere, B.; Bueno, M.; Tacon, P.; Médale, F.; Kaushik, S.J. Influence of the forms and levels of dietary selenium on antioxidant status and oxidative stress-related parameters in rainbow trout (Oncorhynchus mykiss) fry. Br. J. Nutr. 2015, 113, 1876–1887. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Wang, Y.; Gu, Q.; Li, W. Effects of different dietary selenium sources (selenium nanoparticle and selenomethionine) on growth performance, muscle composition and glutathione peroxidase enzyme activity of crucian carp (Carassius auratus gibelio). Aquaculture 2009, 291, 78–81. [Google Scholar] [CrossRef]
- Wang, Y.; Han, J.; Li, W.; Xu, Z. Effect of different selenium source on growth performances, glutathione peroxidase activities, muscle composition and selenium concentration of allogynogenetic crucian carp (Carassius auratus gibelio). Anim. Feed. Sci. Technol. 2007, 134, 243–251. [Google Scholar] [CrossRef]
- Kousoulaki, K.; Mørkøre, T.; Nengas, I.; Berge, R.; Sweetman, J. Microalgae and organic minerals enhance lipid retention efficiency and fillet quality in Atlantic salmon (Salmo salar L.). Aquaculture 2016, 451, 47–57. [Google Scholar] [CrossRef]
- Zhao, Z.; Barcus, M.; Kim, J.; Lum, K.L.; Mills, C.; Lei, X.G. High Dietary Selenium Intake Alters Lipid Metabolism and Protein Synthesis in Liver and Muscle of Pigs. J. Nutr. 2016, 146, 1625–1633. [Google Scholar] [CrossRef]
- Hu, X.; Chandler, J.D.; Orr, M.L.; Hao, L.; Liu, K.; Uppal, K.; Go, Y.-M.; Jones, D.P. Selenium Supplementation Alters Hepatic Energy and Fatty Acid Metabolism in Mice. J. Nutr. 2018, 148, 675–684. [Google Scholar] [CrossRef]
- Yu, L.; Wang, R.; Zhang, Y.; Kleemann, D.; Zhu, X.; Jia, Z. Effects of selenium supplementation on polyunsaturated fatty acid concentrations and antioxidant status in plasma and liver of lambs fed linseed oil or sunflower oil diets. Anim. Feed. Sci. Technol. 2008, 140, 39–51. [Google Scholar] [CrossRef]
- del Puerto, M.; Cabrera, M.C.; Saadoun, A. A Note on Fatty Acids Profile of Meat from Broiler Chickens Supplemented with Inorganic or Organic Selenium. Int. J. Food Sci. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, M.; Montero, D.; Robaina, L.; Caballero, M.J.; Rosenlund, G.; Ginés, R. Alterations in fillet fatty acid profile and flesh quality in gilthead seabream (Sparus aurata) fed vegetable oils for a long term period. Recovery of fatty acid profiles by fish oil feeding. Aquaculture 2005, 250, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, M.S. Essential fatty acid requirements of cultured marine fish larvae. Aquac. Nutr. 1996, 2, 183–191. [Google Scholar] [CrossRef]
- Carvalho, M.; Montero, D.; Rosenlund, G.; Fontanillas, R.; Ginés, R.; Izquierdo, M. Effective complete replacement of fish oil by combining poultry and microalgae oils in practical diets for gilthead sea bream (Sparus aurata) fingerlings. Aquaculture 2020, 529, 735696. [Google Scholar] [CrossRef]
- Senadheera, S.D.; Turchini, G.M.; Thanuthong, T.; Francis, D. Effects of dietary iron supplementation on growth performance, fatty acid composition and fatty acid metabolism in rainbow trout (Oncorhynchus mykiss) fed vegetable oil based diets. Aquaculture 2012, 342, 80–88. [Google Scholar] [CrossRef]
- Yu, H.; Chen, B.; Li, L.; Zhang, Q.; Liu, D.; Wang, Z.; Shan, L. Dietary iron (Fe) requirement of coho salmon (Oncorhynchus kisutch) alevins assessed using growth, whole body and hepatic Fe concentrations and hepatic antioxidant enzymes activities. Aquac. Res. 2021, 52, 4489–4497. [Google Scholar] [CrossRef]
- Luo, Z.; Tan, X.-Y.; Zheng, J.-L.; Chen, Q.-L.; Liu, C.-X. Quantitative dietary zinc requirement of juvenile yellow catfish Pelteobagrus fulvidraco, and effects on hepatic intermediary metabolism and antioxidant responses. Aquaculture 2011, 319, 150–155. [Google Scholar] [CrossRef]
- Musharraf, M.; Khan, M.A. Dietary zinc requirement of fingerling Indian major carp, Labeo rohita (Hamilton). Aquaculture 2019, 503, 489–498. [Google Scholar] [CrossRef]
- Mahfouz, M.M.; Kummerow, F.A. Effect of magnesium deficiency on Δ6 desaturase activity and fatty acid composition of rat liver microsomes. Lipids 1989, 24, 727–732. [Google Scholar] [CrossRef]
- Wei, C.-C.; Wu, K.; Gao, Y.; Zhang, L.-H.; Li, D.-D.; Luo, Z. Magnesium reduces hepatic lipid accumulation in yellow catfish (Pelteobagrus fulvidraco) and modulates lipogenesis and lipolysis via PPARA, JAK-STAT, and AMPK pathways in hepatocytes. J. Nutr. 2017, 147, 1070–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, F.; Li, M.; Tao, Z.; Yuan, L.; Song, M.; Ren, Q.; Xin, X.; Meng, Q.; Wang, R. Effect of high dietary copper on growth, antioxidant and lipid metabolism enzymes of juvenile larger yellow croaker Larimichthys croceus. Aquac. Rep. 2016, 3, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.-X.; Zhuo, M.-Q.; Li, D.-D.; Xu, Y.-H.; Wu, K.; Luo, Z. SREBP-1 and LXRα pathways mediated Cu-induced hepatic lipid metabolism in zebrafish Danio rerio. Chemosphere 2019, 215, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Naiel, M.A.; Negm, S.S.; El-Hameed, S.A.A.; Abdel-Latif, H.M. Dietary organic selenium improves growth, serum biochemical indices, immune responses, antioxidative capacity, and modulates transcription of stress-related genes in Nile tilapia reared under sub-optimal temperature. J. Therm. Biol. 2021, 99, 102999. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Shiau, S.-Y. The effects of dietary selenium on the oxidative stress of grouper, Epinephelus malabaricus, fed high copper. Aquaculture 2007, 267, 38–43. [Google Scholar] [CrossRef]
- Wiseman, S.; Thomas, J.K.; McPhee, L.; Hursky, O.; Raine, J.C.; Pietrock, M.; Giesy, J.P.; Hecker, M.; Janz, D.M. Attenuation of the cortisol response to stress in female rainbow trout chronically exposed to dietary selenomethionine. Aquat. Toxicol. 2011, 105, 643–651. [Google Scholar] [CrossRef]
- Hilton, J.W.; Hodson, P.V.; Slinger, S.J. The Requirement and Toxicity of Selenium in Rainbow Trout (Salmo Gairdneri). J. Nutr. 1980, 110, 2527–2535. [Google Scholar] [CrossRef]
- Ganga, R.; Bell, J.G.; Montero, D.; Atalah, E.; Vraskou, Y.; Tort, L.; Fernandez, A.; Izquierdo, M.S. Adrenocorticotrophic hormone-stimulated cortisol release by the head kidney inter-renal tissue from sea bream (Sparus aurata) fed with linseed oil and soyabean oil. Br. J. Nutr. 2011, 105, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Biller-Takahashi, J.D.; Takahashi, L.S.; Mingatto, F.E.; Urbinati, E.C. The immune system is limited by oxidative stress: Dietary selenium promotes optimal antioxidative status and greatest immune defense in pacu Piaractus mesopotamicus. Fish Shellfish Immunol. 2015, 47, 360–367. [Google Scholar] [CrossRef]
- Salinas, I.; Lockhart, K.; Bowden, T.; Collet, B.; Secombes, C.; Ellis, A. An assessment of immunostimulants as Mx inducers in Atlantic salmon (Salmo salar L.) parr and the effect of temperature on the kinetics of Mx responses. Fish Shellfish Immunol. 2004, 17, 159–170. [Google Scholar] [CrossRef]
- Wang, C.; Lovell, R.T.; Klesius, P.H. Response toEdwardsiella ictaluriChallenge by Channel Catfish Fed Organic and Inorganic Sources of Selenium. J. Aquat. Anim. Health 1997, 9, 172–179. [Google Scholar] [CrossRef]
- Kim, K.; Wang, X.; Choi, S.; Park, G.; Koo, J.; Bai, S.C. No synergistic effects by the dietary supplementation of ascorbic acid, α-tocopheryl acetate and selenium on the growth performance and challenge test of Edwardsiella tarda in fingerling Nile tilapia, Oreochromis niloticus L. Aquac. Res. 2003, 34, 1053–1058. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Mousa, M.A.; Abbass, F.E. Growth performance and physiological response of African catfish, Clarias gariepinus (B.) fed organic selenium prior to the exposure to environmental copper toxicity. Aquaculture 2007, 272, 335–345. [Google Scholar] [CrossRef]
- Cotter, P.A.; Craig, S.R.; McLean, E. Hyperaccumulation of selenium in hybrid striped bass: A functional food for aquaculture? Aquac. Nutr. 2008, 14, 215–222. [Google Scholar] [CrossRef]
- Haller, O.; Kochs, G. Interferon-Induced Mx Proteins: Dynamin-Like GTPases with Antiviral Activity. Traffic 2002, 3, 710–717. [Google Scholar] [CrossRef]
- Pacitti, D.; Lawan, M.M.; Feldmann, J.; Sweetman, J.; Wang, T.; Martin, S.A.M.; Secombes, C.J. Impact of selenium supplementation on fish antiviral responses: A whole transcriptomic analysis in rainbow trout (Oncorhynchus mykiss) fed supranutritional levels of Sel-Plex®. BMC Genom. 2016, 17, 116. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The Role of Selenium in Inflammation and Immunity: From Molecular Mechanisms to Therapeutic Opportunities. Antioxidants Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Petasis, N.A. Resolvins and Protectins in Inflammation Resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [Green Version]
- Weylandt, K.H.; Chiu, C.-Y.; Gomolka, B.; Waechter, S.F.; Wiedenmann, B. Omega-3 fatty acids and their lipid mediators: Towards an understanding of resolvin and protectin formation. Prostaglandins Other Lipid Mediat. 2012, 97, 73–82. [Google Scholar] [CrossRef]
- Magalhães, R.; Guardiola, F.; Guerreiro, I.; Fontinha, F.; Moutinho, S.; Olsen, R.; Peres, H.; Oliva-Teles, A. Effect of different dietary arachidonic, eicosapentaenoic, and docosahexaenoic acid content on selected immune parameters in gilthead sea bream (Sparus aurata). Fish Shellfish Immunol. Rep. 2021, 2, 100014. [Google Scholar] [CrossRef]
- Khair-El-Din, T.A.; Sicher, S.C.; Vazquez, M.A.; Wright, W.J.; Lu, C.Y. Docosahexaenoic acid, a major constituent of fetal serum and fish oil diets, inhibits IFN gamma-induced Ia-expression by murine macrophages in vitro. J. Immunol. 1995, 154, 1296–1306. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (%) | Diets | ||
|---|---|---|---|
| Control (0.29) | OH-SeMet (0.52) | OH-SeMet (0.79) | |
| Fish meal a | 10.00 | 10.00 | 10.00 |
| Blood meal b | 6.00 | 6.00 | 6.00 |
| Rapeseed meal a | 12.70 | 12.70 | 12.70 |
| Corn gluten meal a | 18.40 | 18.40 | 18.40 |
| Soy protein concentrate | 18.30 | 18.30 | 18.30 |
| Wheat meal | 4.72 | 4.72 | 4.72 |
| Wheat gluten | 6.00 | 6.00 | 6.00 |
| Fish oil | 7.00 | 7.00 | 7.00 |
| Vegetable oil mix c | 10.00 | 10.00 | 10.00 |
| Vitamin mix | 2.00 | 2.00 | 2.00 |
| Mineral mix | 2.00 | 2.00 | 2.00 |
| OH-SeMet d | - | 0.00125 | 0.0025 |
| Ca(H2PO4)2 | 1.62 | 1.62 | 1.62 |
| L-Lysine | 0.46 | 0.46 | 0.46 |
| DL-Methionine | 0.06 | 0.06 | 0.06 |
| L-Histidine | 0.14 | 0.14 | 0.14 |
| Cholesterol | 0.11 | 0.11 | 0.11 |
| CMC | 0.50 | 0.50 | 0.50 |
| Proximate composition (%) | |||
| Ash % (DM) | 6.98 | 6.96 | 7.05 |
| Total lipids% (DM) | 21.21 | 21.85 | 22.48 |
| Crude protein% (DM) | 48.20 | 48.05 | 48.23 |
| Analysed Se content | |||
| (mg kg−1) | 0.29 | 0.52 | 0.79 |
| Fatty Acids | Diets | ||
|---|---|---|---|
| Control (0.29) | OH-SeMet (0.52) | OH-SeMet (0.79) | |
| 11:0 | 0.16 | 0.13 | 0.13 |
| 12:0 | 0.46 | 0.45 | 0.46 |
| 13:0 | 2.66 | 2.82 | 2.92 |
| 14:0 | 2.17 | 2.21 | 2.21 |
| 14:1n-5 | 0.07 | 0.07 | 0.07 |
| 15:0 | 0.24 | 0.24 | 0.24 |
| 16:0 | 18.76 | 18.52 | 19.10 |
| 16:1n-7 | 2.28 | 2.34 | 2.32 |
| 16:1n-5 | 0.10 | 0.11 | 0.11 |
| 16:2n-4 | 0.17 | 0.18 | 0.17 |
| 17:0 | 0.14 | 0.14 | 0.13 |
| 16:3n-4 | 0.16 | 0.15 | 0.16 |
| 16:3n-3 | 0.07 | 0.07 | 0.07 |
| 16:4n-3 | 0.17 | 0.18 | 0.16 |
| 18:0 | 4.14 | 4.13 | 4.26 |
| 18:1n-9 | 34.46 | 33.87 | 34.47 |
| 18:1n-7 | 2.32 | 2.33 | 2.32 |
| 18:1n-5 | 0.07 | 0.07 | 0.07 |
| 18:2n-6 | 13.64 | 13.43 | 13.25 |
| 18:2n-4 | 0.06 | 0.06 | 0.06 |
| 18:3n-6 | 0.08 | 0.09 | 0.08 |
| 18:3n-4 | 0.05 | 0.06 | 0.06 |
| 18:3n-3 | 6.99 | 6.87 | 6.52 |
| 18:4n-3 | 0.37 | 0.41 | 0.36 |
| 18:4n-1 | 0.04 | 0.04 | 0.04 |
| 20:0 | 0.41 | 0.40 | 0.42 |
| 20:1n-9 | 0.15 | 0.16 | 0.16 |
| 20:1n-7 | 1.51 | 1.52 | 1.54 |
| 20:1n-5 | 0.11 | 0.11 | 0.12 |
| 20:2n-9 | 0.09 | 0.18 | 0.20 |
| 20:3n-9 | 0.05 | 0.05 | 0.05 |
| 20:4n-6 | 0.32 | 0.34 | 0.31 |
| 20:3n-3 | 0.07 | 0.07 | 0.07 |
| 20:4n-3 | 0.20 | 0.21 | 0.20 |
| 20:5n-3 | 2.24 | 2.45 | 2.17 |
| 22:1n-11 | 1.01 | 1.03 | 1.05 |
| 22:1n-9 | 0.23 | 0.23 | 0.24 |
| 22:4n-6 | 0.05 | 0.05 | 0.05 |
| 22:5n-6 | 0.02 | 0.02 | 0.02 |
| 22:5n-3 | 0.44 | 0.49 | 0.43 |
| 22:6n-3 | 3.13 | 3.57 | 3.08 |
| ∑SFA § | 29.17 | 29.08 | 29.90 |
| ∑MUFA § | 42.10 | 41.62 | 42.25 |
| ∑PUFA § | 28.50 | 29.07 | 27.61 |
| ∑n-3 | 13.67 | 14.32 | 13.07 |
| ∑n-6 | 14.17 | 14.00 | 13.79 |
| ∑n-9 | 34.76 | 34.28 | 34.90 |
| n-3/n-6 | 0.96 | 1.02 | 0.95 |
| DHA/EPA § | 1.40 | 1.46 | 1.42 |
| EPA/ARA § | 7.03 | 7.19 | 6.89 |
| Gene | Primers Sequence | Pmol/µL | Reference |
|---|---|---|---|
| β-actin | Forward 5′-TCT GTC TGG ATC GGA GGC TC-3′ | 10 | Grasso et al. [33] |
| Reverse 5′-AAG CAT TTG CGG TGG ACG-3′ | |||
| Mx | Forward 5′-GAC AGG GAG CGG CAT TGT TAC-3′ | 10 | Grasso et al. [33] |
| Reverse 5′-TCG TCC AGC TCT TCC TCG TG-3′ | |||
| TNF-a | Forward 5′-TCG TTC AGA GTC TCC TGC AG-3′ | 10 | Grasso et al. [33] |
| Reverse 5′-CAT GGA CTC TGA GTA GCG CGA-3′ | |||
| COX | Forward 5′-GAG TAC TGG AAG CCG AGC AC-3′ | 10 | Grasso et al. [33] |
| Reverse 5′-GAT ATC ACT GCC GCC TGA CT-3′ | |||
| IL-Ir2 | Forward 5′-AAG GAC TCC AGC TCC ACT GA-3′ | 10 | Grasso et al. [33] |
| Reverse 5′-ACG CCT TCT ACA TGG ACC AC-3′ | |||
| Casp-3 | Forward 5′-CTGATCTGGATGGAGGCATT-3′ | 10 | Morcillo et al. [35] |
| Reverse 5′-AGTAGTAGCCTGGGGCTGTG-3′ | |||
| IL-1β | Forward 5′-AGC GAC ATG GCA CGA TTT-3′ | 10 | Grasso et al. [33] |
| Reverse 5′-GCA CTC TCC TGG CAC ATA TCC-3′ | |||
| IL-6 | Forward 5′-GCT CTG CTG GGT GTG CTC C-3′ | 10 | Castellana et al. [36] |
| Reverse 5′-GTC TCC CAC TCC TCA CCT TG-3′ | |||
| IL-10 | Forward 5′-TGGAGGGCTTTCCTGTCAGA-3′ | 10 | Pellizzari et al. [37] |
| Reverse 5′-TGCTTCGTAGAAGTCTCGGATGT-3′ |
| Diets | |||
|---|---|---|---|
| Se Content in Water (µg L−1) | Control (0.29) | OH-SeMet (0.52) | OH-SeMet (0.79) |
| Before feeding (0 day) | 1.14 ± 0.06 | 0.78 ± 0.15 | 0.92 ± 0.54 |
| After feeding (4 day) | 1.72 ± 0.09 | 1.69 ± 0.28 | 1.67 ± 0.29 |
| Se content in body tissues (mg kg−1) | |||
| Liver | 0.79 ± 0.02 a | 1.04 ± 0.08 a,b | 1.35 ± 0.21 b |
| Muscle | 0.18 ± 0.01 | 0.45 ± 0.19 | 0.38 ± 0.04 |
| Diets | Control (0.29) | OH-SeMet (0.52) | OH-SeMet (0.79) |
|---|---|---|---|
| Survival rate (%) | 97.30 ± 3.82 | 98.65 ± 1.91 | 97.30 ± 3.82 |
| Total length (cm) | 15.22 ± 0.27 | 15.22 ± 0.18 | 15.28 ± 0.14 |
| Body weight (g) | 51.54 ± 4.19 | 49.68 ± 1.19 | 50.89 ± 2.57 |
| WG a (%) | 135.77 ±18.20 | 126.95 ± 3.44 | 135.50 ± 9.17 |
| SGR (%) | 0.93 ± 0.08 | 0.89 ± 0.02 | 0.93 ± 0.04 |
| FI (g) | 52.00 ± 4.68 | 51.01 ± 5.08 | 55.21 ± 2.39 |
| FE | 0.57 ± 0.03 | 0.55 ± 0.03 | 0.53 ± 0.02 |
| CF | 1.91 ± 0.07 | 1.86 ± 0.01 | 1.43 ± 0.03 |
| Diets | |||
|---|---|---|---|
| (% Fresh Weight) | Control (0.29) | OH-SeMet (0.52) | OH-SeMet (0.79) |
| Lipid | 9.46 ± 0.60 a | 10.45 ± 0.37 b | 10.83 ± 0.65 b |
| Moisture | 68.61 ± 0.85 b | 65.75 ± 2.64 a | 64.86 ± 2.77 a |
| Protein | 16.73 ± 0.07 | 17.17 ± 0.64 | 17.51 ± 1.88 |
| Ash | 3.80 ± 0.31 | 3.99 ± 0.43 | 4.02 ± 0.44 |
| Diets | |||
|---|---|---|---|
| Fatty Acids | Control (0.29) | OH-SeMet (0.52) | OH-SeMet (0.79) |
| 11:0 | 0.07 ± 0.03 | 0.05 ± 0.01 | 0.05 ± 0.03 |
| 12:0 | 0.13 ± 0.01 b | 0.11 ± 0.00 a | 0.13 ± 0.00 ab |
| 13:0 | 6.92 ± 0.73 | 4.58 ± 0.99 | 5.70 ± 1.45 |
| 14:0 | 2.83 ± 0.05 | 2.79 ± 0.01 | 2.65 ± 0.12 |
| 14:1n-7 | 0.05 ± 0.00 | 0.05 ± 0.00 | 0.05 ± 0.00 |
| 14:1n-5 | 0.08 ± 0.00 | 0.08 ± 0.00 | 0.07 ± 0.00 |
| 15:0 | 0.28 ± 0.01 b | 0.27 ±0.00 a | 0.26 ± 0.01 a |
| 16:0 | 15.53 ± 0.29 b | 14.53. ± 0.12 a | 14.47 ± 0.32 a |
| 16:1n-7 | 4.14 ± 0.06 | 4.27 ± 0.03 | 4.09 ± 0.20 |
| 16:1n-5 | 0.11 ± 0.00 | 0.11 ± 0.00 | 0.11 ± 0.00 |
| 16:2n-4 | 0.23 ± 0.01 a | 0.26 ± 0.00 b | 0.25 ± 0.01 b |
| 17:0 | 0.22 ± 0.08 | 0.22 ± 0.00 | 0.22 ± 0.01 |
| 16:3n-4 | 0.25 ± 0.06 | 0.22 ± 0.01 | 0.22 ± 0.00 |
| 16:3n-3 | 0.07 ± 0.00 | 0.07 ± 0.00 | 0.07 ± 0.00 |
| 16:3n-1 | 0.07 ± 0.00 | 0.08 ± 0.00 | 0.07 ± 0.00 |
| 16:4n-3 | 0.13 ± 0.02 a | 0.21 ± 0.01 b | 0.20 ± 0.01 b |
| 18:0 | 3.46 ± 0.09 b | 3.37 ± 0.03 b | 3.24 ± 0.05 a |
| 18:1n-9 | 32.90 ± 0.62 b | 31.29 ± 0.36 a | 31.16 ± 0.81 a |
| 18:1n-7 | 2.84 ± 0.04 b | 2.83 ± 0.03 b | 2.73 ± 0.02 a |
| 18:1n-5 | 0.11 ± 0.00 | 0.12 ± 0.00 | 0.11 ± 0.01 |
| 18:2n-9 | 0.37 ± 0.04 a | 0.40 ± 0.04 ab | 0.47 ± 0.03 b |
| 18:2n-6 | 12.11 ± 0.15 | 12.25 ± 0.14 | 12.19 ± 0.35 |
| 18:2n-4 | 0.03 ± 0.01 a | 0.02 ± 0.00 a | 0.04 ± 0.00 b |
| 18:3n-6 | 0.12 ± 0.01 | 0.13 ± 0.00 | 0.13 ± 0.00 |
| 18:3n-4 | 0.11 ± 0.00 a | 0.13 ± 0.01 c | 0.12 ± 0.00 b |
| 18:3n-3 | 4.11 ± 0.16 | 4.32 ± 0.13 | 4.48 ± 0.37 |
| 18:4n-3 | 0.49 ± 0.07 a | 0.70 ± 0.02 b | 0.71 ± 0.01 b |
| 18:4n-1 | 0.06 ± 0.01 a | 0.09 ± 0.00 b | 0.09 ± 0.00 b |
| 20:0 | 0.28 ± 0.01 b | 0.26 ± 0.00 a | 0.26 ± 0.01 a |
| 20:1n-9 | 0.29 ± 0.01 | 0.31 ± 0.00 | 0.29 ± 0.02 |
| 20:1n-7 | 1.82 ± 0.07 | 1.80 ± 0.04 | 1.73 ± 0.08 |
| 20:1n-5 | 0.14 ± 0.00 | 0.14 ± 0.00 | 0.14 ± 0.00 |
| 20:2n-9 | 0.23 ± 0.01 | 0.22 ± 0.03 | 0.25 ± 0.02 |
| 20:2n-6 | 0.36 ± 0.02 a | 0.42 ± 0.00 b | 0.40 ± 0.01 b |
| 20:3n-9 | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 |
| 20:3n-6 | 0.15 ± 0.00 a | 0.16 ± 0.01 b | 0.17 ± 0.01 b |
| 20:4n-6 | 0.35 ± 0.04 a | 0.44 ± 0.01 b | 0.43 ± 0.01 b |
| 20:3n-3 | 0.17 ± 0.01 a | 0.19 ± 0.00 b | 0.19 ± 0.00 b |
| 20:4n-3 | 0.35 ± 0.04 a | 0.40 ± 0.03 b | 0.40 ± 0.01 b |
| 20:5n-3 | 1.93 ± 0.32 a | 2.98 ± 0.11 b | 2.90 ± 0.10 b |
| 22:1n-11 | 1.13 ± 0.06 | 1.15 ± 0.02 | 1.09 ± 0.06 |
| 22:1n-9 | 0.43 ± 0.02 | 0.42 ± 0.00 | 0.42 ± 0.02 |
| 22:4n-6 | 0.10 ± 0.00 a | 0.11 ± 0.00 b | 0.11 ± 0.00 b |
| 22:5n-6 | 0.03 ± 0.00 | 0.03 ± 0.00 | 0.03 ± 0.01 |
| 22:5n-3 | 0.88 ± 0.13 a | 1.41 ± 0.06 b | 1.36 ± 0.03 b |
| 22:6n-3 | 3.41 ± 0.65 a | 5.84 ± 0.26 b | 5.73 ± 0.07 b |
| ∑SFA * | 29.77 ±1.02 b | 26.23 ± 0.88 a | 26.92 ± 1.23 a |
| ∑MUFA * | 44.07 ± 0.64 b | 42.58 ± 0.45 a | 42.00 ± 0.60 a |
| ∑PUFA * | 26.16 ±1.58 a | 31.18 ± 0.55 b | 31.09 ± 0.64 b |
| ∑n−3 | 11.56 ±1.40 a | 16.16 ± 0.48 b | 16.08 ± 0.33 b |
| ∑n−6 | 13.22 ± 0.21 | 13.55 ± 0.13 | 13.46 ± 0.32 |
| ∑n−9 | 34.23 ± 0.64 b | 32.67 ± 0.35 a | 32.60 ± 0.80 a |
| n−3/n−6 | 0.87 ± 0.09 a | 1.19 ± 0.04 b | 1.19 ± 0.01 b |
| DHA/EPA * | 1.76 ± 0.05 a | 1.96 ± 0.02 b | 1.98 ± 0.07 b |
| EPA/ARA * | 5.54 ± 0.40 a | 6.82 ± 0.08 b | 6.79 ± 0.10 b |
| 18:4n-3/18:3n-3 | 0.12 ± 0.01 a | 0.16 ± 0.01 b | 0.16 ± 0.02 b |
| Diets | R2 | p Value | |||
|---|---|---|---|---|---|
| Fatty Acid | Control (0.29) | OH-SeMet (0.52) | OH-SeMet (0.79) | ||
| 20:4 n-6 | 47.04 ± 8.61 a | 65.65 ± 1.08 b | 68.16 ± 9.79 b | 0.94 | 0.027 |
| 20:5 n-3 | 41.51 ± 8.98 a | 70.78 ± 1.01 b | 76.10 ± 8.20 b | 0.95 | 0.002 |
| 22:6 n-3 | 57.12 ± 13.65 a | 101.19 ± 1.45 b | 113.46 ± 15.82 b | 0.98 | 0.003 |
| n-3 LC-PUFA | 44.59 ± 6.88 a | 67.54 ± 1.68 ab | 72.91 ± 11.20 b | 0.97 | 0.009 |
| DHA/EPA | 1.37 ± 0.26 a | 1.43 ± 0.16 ab | 1.49 ± 0.29 b | 0.98 | 0.026 |
| EPA/ARA | 0.88 ± 0.19 a | 1.08 ± 0.06 b | 1.12 ± 0.08 b | 0.96 | 0.009 |
| 18:4 n-3/18:3 n-3 | 1.19 ± 0.04 a | 2.60 ± 0.01 b | 2.72 ± 0.05 b | 0.95 | 0.013 |
| n-3/n-6 | 1.07 ± 0.10 a | 1.45 ± 0.02 b | 1.58 ± 0.04 b | 0.98 | 0.006 |
| 18:3 n-6/18:2 n-6 | 0.95 ± 0.18 | 1.12 ± 0.07 | 1.20 ± 0.07 | 0.99 | 0.096 |
| Diets | One-Way ANOVA | Two-Way ANOVA | |||||
|---|---|---|---|---|---|---|---|
| Plasma Cortisol (ng mL−1) | Control (0.29) | OH-SeMet (0.52) | OH-SeMet (0.79) | p Value | Time | Se Level | Time × Se Level |
| 0 h | 8.39 ± 1.40 | 8.57 ± 1.83 | 10.75± 4.65 | 0.588 | 0.000 | 0.013 | 0.017 |
| 2 h | 70.75± 37.89 | 75.25 ± 1.77 | 185.67 ± 71.65 | 0.093 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseng, Y.; Dominguez, D.; Bravo, J.; Acosta, F.; Robaina, L.; Geraert, P.-A.; Kaushik, S.; Izquierdo, M. Organic Selenium (OH-MetSe) Effect on Whole Body Fatty Acids and Mx Gene Expression against Viral Infection in Gilthead Seabream (Sparus aurata) Juveniles. Animals 2021, 11, 2877. https://doi.org/10.3390/ani11102877
Tseng Y, Dominguez D, Bravo J, Acosta F, Robaina L, Geraert P-A, Kaushik S, Izquierdo M. Organic Selenium (OH-MetSe) Effect on Whole Body Fatty Acids and Mx Gene Expression against Viral Infection in Gilthead Seabream (Sparus aurata) Juveniles. Animals. 2021; 11(10):2877. https://doi.org/10.3390/ani11102877
Chicago/Turabian StyleTseng, Yiyen, David Dominguez, Jimena Bravo, Felix Acosta, Lidia Robaina, Pierre-André Geraert, Sadasivam Kaushik, and Marisol Izquierdo. 2021. "Organic Selenium (OH-MetSe) Effect on Whole Body Fatty Acids and Mx Gene Expression against Viral Infection in Gilthead Seabream (Sparus aurata) Juveniles" Animals 11, no. 10: 2877. https://doi.org/10.3390/ani11102877
APA StyleTseng, Y., Dominguez, D., Bravo, J., Acosta, F., Robaina, L., Geraert, P. -A., Kaushik, S., & Izquierdo, M. (2021). Organic Selenium (OH-MetSe) Effect on Whole Body Fatty Acids and Mx Gene Expression against Viral Infection in Gilthead Seabream (Sparus aurata) Juveniles. Animals, 11(10), 2877. https://doi.org/10.3390/ani11102877