
Analyses of Molecular Characteristics and Enzymatic Activities of Ovine HSD17B3

 ,
,  , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Collection
2.2. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) and Sequencing
2.3. Sequence Alignment and Phylogenetic Analysis
2.4. Cell Culture, Transfection, and Luciferase Assay
2.5. Plasmids
2.6. Western Blotting Analyses
2.7. Measurements by Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
2.8. Statistical Analyses
3. Results
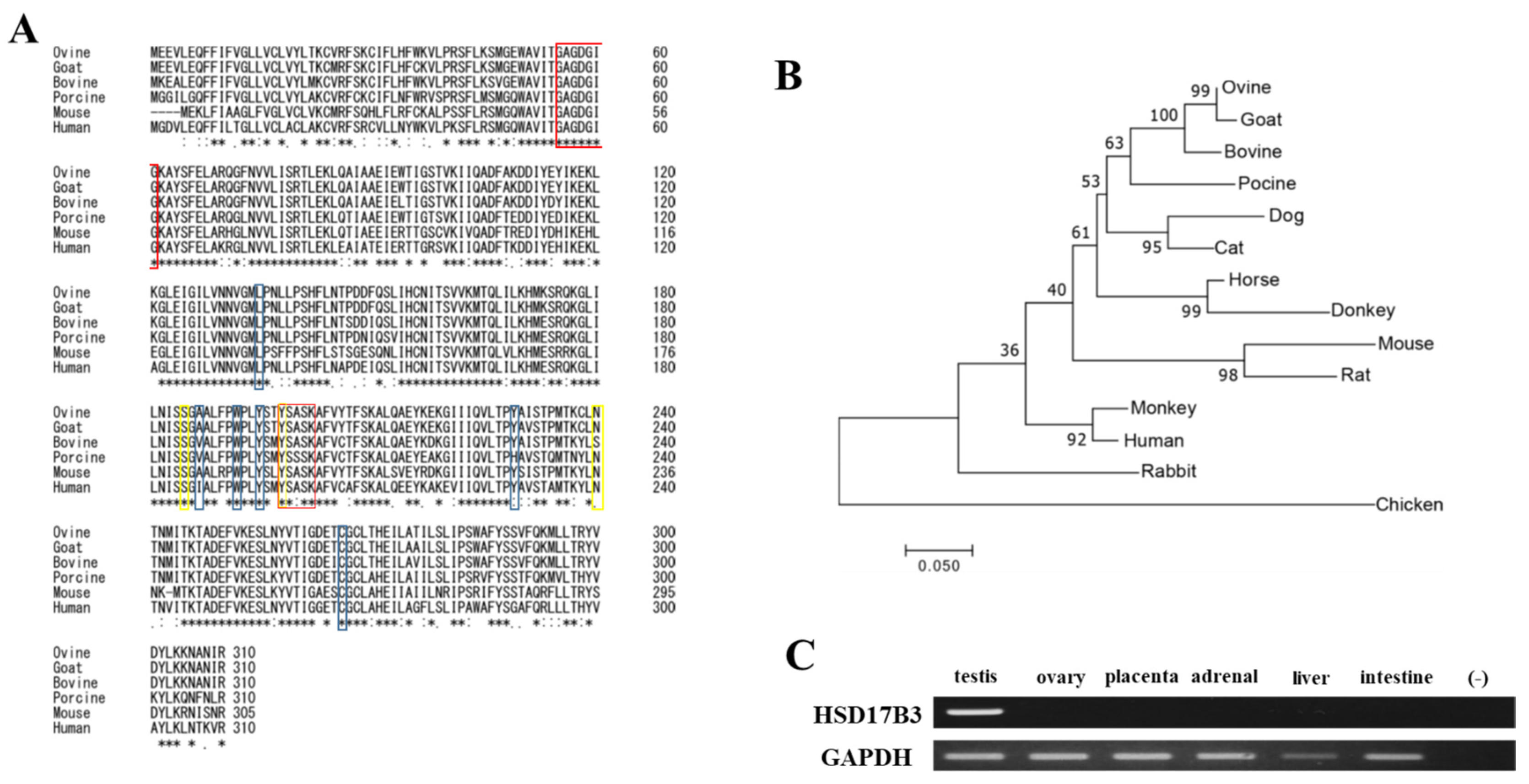
3.1. Conservation of Ovine HSD17B3 Gene
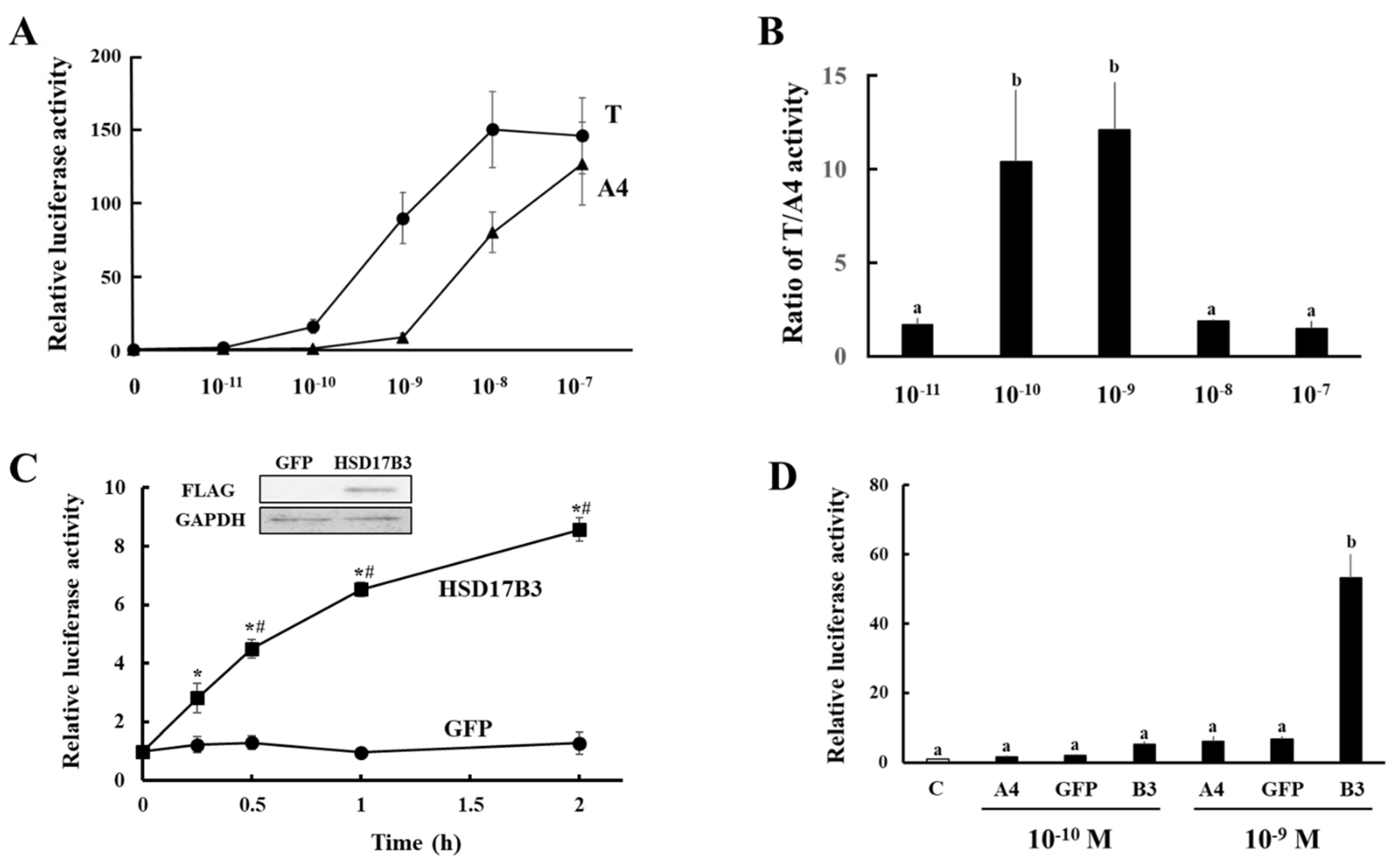
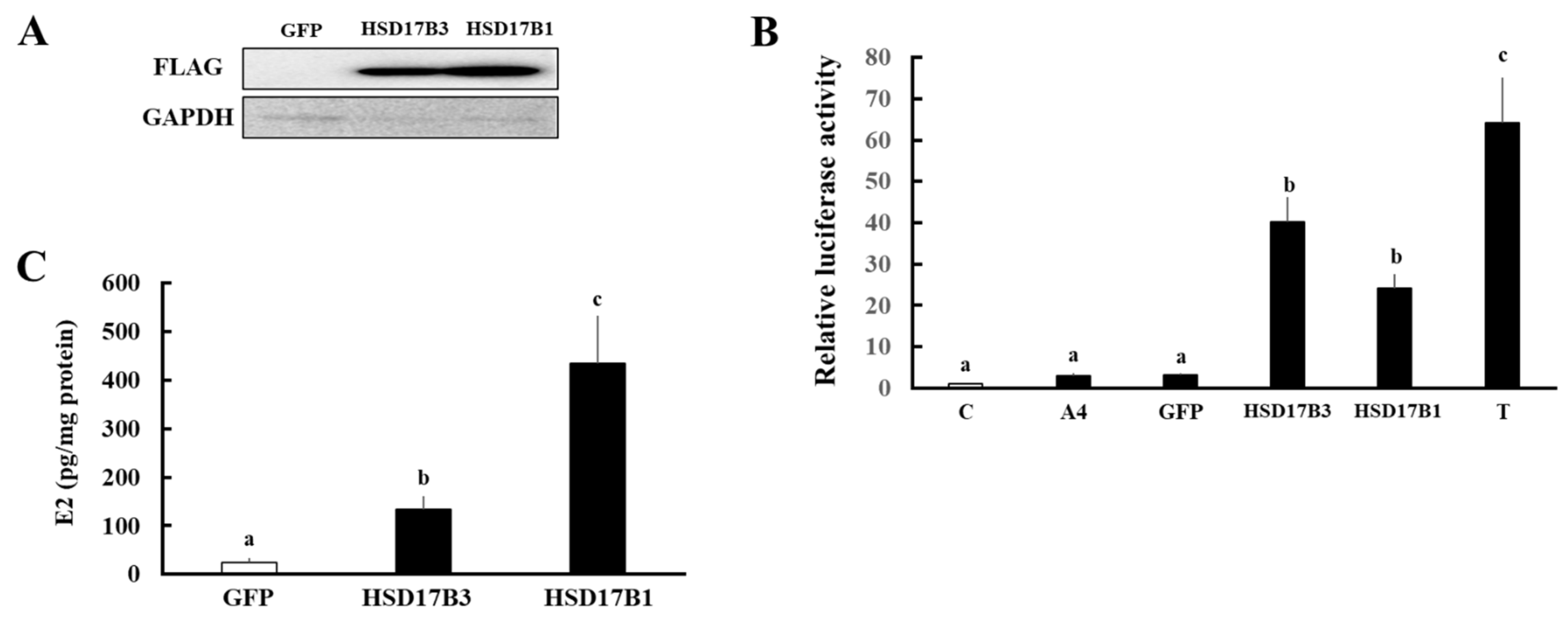
3.2. Evaluation of Enzymatic Activities of Ovine HSD17B3
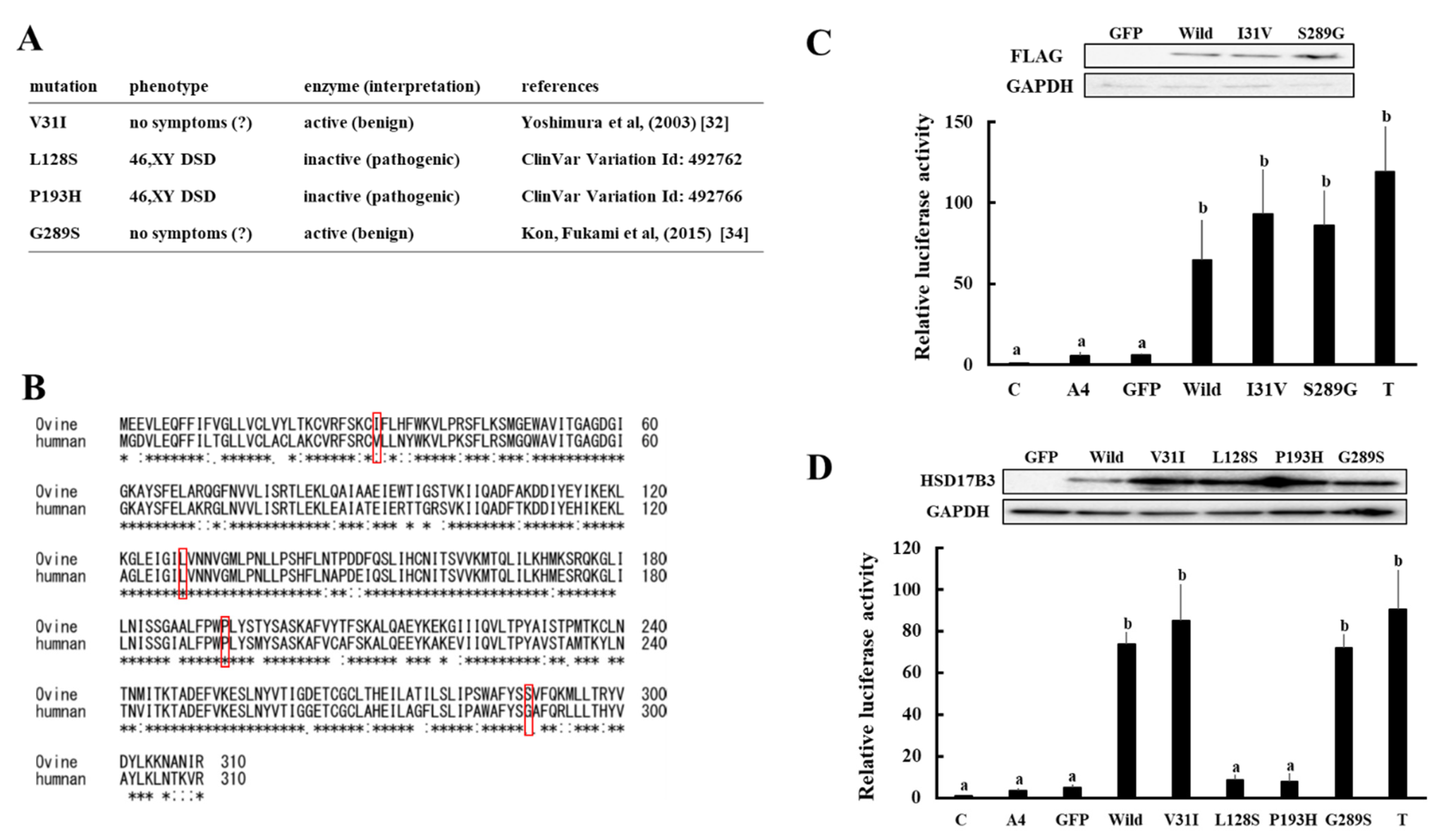
3.3. Enzymatic Activities Associated with Various Missense Mutations in Ovine and Human HSD17B3 Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.L. Steroidogenesis: Unanswered Questions. Trends Endocrinol. Metab. Tem 2017, 28, 771–793. [Google Scholar] [CrossRef]
- Tsachaki, M.; Odermatt, A. Subcellular localization and membrane topology of 17β-hydroxysteroid dehydrogenases. Mol. Cell. Endocrinol. 2019, 489, 98–106. [Google Scholar] [CrossRef]
- He, W.; Gauri, M.; Li, T.; Wang, R.; Lin, S.X. Current knowledge of the multifunctional 17β-hydroxysteroid dehydrogenase type 1 (HSD17B1). Gene 2016, 588, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Heinosalo, T.; Saarinen, N.; Poutanen, M. Role of hydroxysteroid (17beta) dehydrogenase type 1 in reproductive tissues and hormone-dependent diseases. Mol. Cell. Endocrinol. 2019, 489, 9–31. [Google Scholar] [CrossRef] [PubMed]
- George, M.M.; New, M.I.; Ten, S.; Sultan, C.; Bhangoo, A. The clinical and molecular heterogeneity of 17βHSD-3 enzyme deficiency. Horm. Res. Paediatr. 2010, 74, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, T.; Imamichi, Y.; Uwada, J.; Sekiguchi, T.; Mikami, D.; Kitano, T.; Ida, T.; Sato, T.; Nemoto, T.; Nagata, S.; et al. Evaluation of 17β-hydroxysteroid dehydrogenase activity using androgen receptor-mediated transactivation. J. Steroid Biochem. Mol. Biol. 2020, 196, 105493. [Google Scholar] [CrossRef]
- Geissler, W.M.; Davis, D.L.; Wu, L.; Bradshaw, K.D.; Patel, S.; Mendonca, B.B.; Elliston, K.O.; Wilson, J.D.; Russell, D.W.; Andersson, S. Male pseudohermaphroditism caused by mutations of testicular 17 beta-hydroxysteroid dehydrogenase 3. Nat. Genet. 1994, 7, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Boehmer, A.L.; Brinkmann, A.O.; Sandkuijl, L.A.; Halley, D.J.; Niermeijer, M.F.; Andersson, S.; de Jong, F.H.; Kayserili, H.; de Vroede, M.A.; Otten, B.J.; et al. 17Beta-hydroxysteroid dehydrogenase-3 deficiency: Diagnosis, phenotypic variability, population genetics, and worldwide distribution of ancient and de novo mutations. J. Clin. Endocrinol. Metab. 1999, 84, 4713–4721. [Google Scholar] [CrossRef] [PubMed]
- Krzeminska, P.; Nizanski, W.; Nowacka-Woszuk, J.; Switonski, M. Analysis of testosterone pathway genes in dogs (78,XY; SRY-positive) with ambiguous external genitalia revealed a homozygous animal for 2-bp deletion causing premature stop codon in HSD17B3. Anim. Genet. 2019, 50, 705–711. [Google Scholar] [CrossRef]
- Sipilä, P.; Junnila, A.; Hakkarainen, J.; Huhtaniemi, R.; Mairinoja, L.; Zhang, F.P.; Strauss, L.; Ohlsson, C.; Kotaja, N.; Huhtaniemi, I.; et al. The lack of HSD17B3 in male mice results in disturbed Leydig cell maturation and endocrine imbalance akin to humans with HSD17B3 deficiency. FASEB J. 2020, 34, 6111–6128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albarella, S.; D'Anza, E.; Galdiero, G.; Esposito, L.; De Biase, D.; Paciello, O.; Ciotola, F.; Peretti, V. Cytogenetic Analyses in Ewes with Congenital Abnormalities of the Genital Apparatus. Animal 2019, 9, 776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favetta, L.A.; Villagómez, D.A.; Iannuzzi, L.; Di Meo, G.; Webb, A.; Crain, S.; King, W.A. Disorders of sexual development and abnormal early development in domestic food-producing mammals: The role of chromosome abnormalities, environment and stress factors. Sex. Dev. 2012, 6, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Zhou, Y.; Zhao, H.; Wu, J.; Yu, H.; Yang, Y.; Yang, Y.; Zhao, H.; Li, H. Comprehensive transcriptome analysis of hypothalamus reveals genes associated with disorders of sex development in pigs. J. Steroid Biochem. Mol. Biol. 2021, 210, 105875. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzi, L.; Arrighi, S.; Rossi, E.; Grignani, P.; Previderè, C.; Bonacina, S.; Cremonesi, F.; Parma, P. XY (SRY-positive) Ovarian Disorder of Sex Development in Cattle. Sex. Dev. 2018, 12, 196–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bormann, C.L.; Smith, G.D.; Padmanabhan, V.; Lee, T.M. Prenatal testosterone and dihydrotestosterone exposure disrupts ovine testicular development. Reproduction 2011, 142, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, F.; Rae, M.T.; Bittner, L.; Hogg, K.; McNeilly, A.S.; Duncan, W.C. Excess androgens in utero alters fetal testis development. Endocrinology 2013, 154, 1921–1933. [Google Scholar] [CrossRef]
- Scully, C.M.; Estill, C.T.; Amodei, R.; McKune, A.; Gribbin, K.P.; Meaker, M.; Stormshak, F.; Roselli, C.E. Early prenatal androgen exposure reduces testes size and sperm concentration in sheep without altering neuroendocrine differentiation and masculine sexual behavior. Domest. Anim. Endocrinol. 2018, 62, 1–9. [Google Scholar] [CrossRef]
- Recabarren, S.E.; Recabarren, M.; Sandoval, D.; Carrasco, A.; Padmanabhan, V.; Rey, R.; Richter, H.G.; Perez-Marin, C.C.; Sir-Petermann, T.; Rojas-Garcia, P.P. Puberty arises with testicular alterations and defective AMH expression in rams prenatally exposed to testosterone. Domest. Anim. Endocrinol. 2017, 61, 100–107. [Google Scholar] [CrossRef]
- Robinson, J.E.; Birch, R.A.; Taylor, J.A.; Foster, D.L.; Padmanabhan, V. In utero programming of sexually differentiated gonadotrophin releasing hormone (GnRH) secretion. Domest. Anim. Endocrinol. 2002, 23, 43–52. [Google Scholar] [CrossRef]
- Cheng, G.; Coolen, L.M.; Padmanabhan, V.; Goodman, R.L.; Lehman, M.N. The kisspeptin/neurokinin B/dynorphin (KNDy) cell population of the arcuate nucleus: Sex differences and effects of prenatal testosterone in sheep. Endocrinology 2010, 151, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Roselli, C.E.; Reddy, R.C.; Estill, C.T.; Scheldrup, M.; Meaker, M.; Stormshak, F.; Montilla, H.J. Prenatal influence of an androgen agonist and antagonist on the differentiation of the ovine sexually dimorphic nucleus in male and female lamb fetuses. Endocrinology 2014, 155, 5000–5010. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.C.; Steadman, C.J.; Lee, T.M.; Padmanabhan, V.; Lehman, M.N.; Coolen, L.M. Sex differences and effects of prenatal exposure to excess testosterone on ventral tegmental area dopamine neurons in adult sheep. Eur. J. Neurosci. 2015, 41, 1157–1166. [Google Scholar] [CrossRef] [Green Version]
- Mizutani, T.; Yamada, K.; Yazawa, T.; Okada, T.; Minegishi, T.; Miyamoto, K. Cloning and characterization of gonadotropin-inducible ovarian transcription factors (GIOT1 and -2) that are novel members of the (Cys)(2)-(His)(2)-type zinc finger protein family. Mol. Endocrinol. 2001, 15, 1693–1705. [Google Scholar] [CrossRef]
- Yazawa, T.; Mizutani, T.; Yamada, K.; Kawata, H.; Sekiguchi, T.; Yoshino, M.; Kajitani, T.; Shou, Z.; Umezawa, A.; Miyamoto, K. Differentiation of adult stem cells derived from bone marrow stroma into Leydig or adrenocortical cells. Endocrinology 2006, 147, 4104–4111. [Google Scholar] [CrossRef] [Green Version]
- Yazawa, T.; Inaoka, Y.; Okada, R.; Mizutani, T.; Yamazaki, Y.; Usami, Y.; Kuribayashi, M.; Orisaka, M.; Umezawa, A.; Miyamoto, K. PPAR-gamma coactivator-1alpha regulates progesterone production in ovarian granulosa cells with SF-1 and LRH-1. Mol. Endocrinol. 2010, 24, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Yazawa, T.; Inaba, H.; Imamichi, Y.; Sekiguchi, T.; Uwada, J.; Islam, M.S.; Orisaka, M.; Mikami, D.; Ida, T.; Sato, T.; et al. Profiles of 5α-Reduced Androgens in Humans and Eels: 5α-Dihydrotestosterone and 11-Ketodihydrotestosterone Are Active Androgens Produced in Eel Gonads. Front. Endocrinol. 2021, 12, 657360. [Google Scholar] [CrossRef]
- Imamichi, Y.; Yuhki, K.I.; Orisaka, M.; Kitano, T.; Mukai, K.; Ushikubi, F.; Taniguchi, T.; Umezawa, A.; Miyamoto, K.; Yazawa, T. 11-Ketotestosterone Is a Major Androgen Produced in Human Gonads. J. Clin. Endocrinol. Metab. 2016, 101, 3582–3591. [Google Scholar] [CrossRef]
- Yazawa, T.; Mizutani, T.; Yamada, K.; Kawata, H.; Sekiguchi, T.; Yoshino, M.; Kajitani, T.; Shou, Z.; Miyamoto, K. Involvement of cyclic adenosine 5'-monophosphate response element-binding protein, steroidogenic factor 1, and Dax-1 in the regulation of gonadotropin-inducible ovarian transcription factor 1 gene expression by follicle-stimulating hormone in ovarian granulosa cells. Endocrinology 2003, 144, 1920–1930. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, T.; Sato, T.; Nemoto, T.; Nagata, S.; Imamichi, Y.; Kitano, T.; Sekiguchi, T.; Uwada, J.; Islam, M.S.; Mikami, D.; et al. 11-Ketotestosterone is a major androgen produced in porcine adrenal glands and testes. J. Steroid Biochem. Mol. Biol. 2021, 210, 105847. [Google Scholar] [CrossRef]
- Yoshimura, K.; Hanaoka, T.; Ohnami, S.; Ohnami, S.; Kohno, T.; Liu, Y.; Yoshida, T.; Sakamoto, H.; Tsugane, S. Allele frequencies of single nucleotide polymorphisms (SNPs) in 40 candidate genes for gene-environment studies on cancer: Data from population-based Japanese random samples. J. Hum. Genet. 2003, 48, 654–658. [Google Scholar] [CrossRef] [Green Version]
- Sata, F.; Kurahashi, N.; Ban, S.; Moriya, K.; Tanaka, K.D.; Ishizuka, M.; Nakao, H.; Yahata, Y.; Imai, H.; Kakizaki, H.; et al. Genetic polymorphisms of 17 β-hydroxysteroid dehydrogenase 3 and the risk of hypospadias. J. Sex. Med. 2010, 7, 2729–2738. [Google Scholar] [CrossRef]
- Kon, M.; Suzuki, E.; Dung, V.C.; Hasegawa, Y.; Mitsui, T.; Muroya, K.; Ueoka, K.; Igarashi, N.; Nagasaki, K.; Oto, Y.; et al. Molecular basis of non-syndromic hypospadias: Systematic mutation screening and genome-wide copy-number analysis of 62 patients. Hum. Reprod. 2015, 30, 499–506. [Google Scholar] [CrossRef] [Green Version]
- Hughes, I.A.; Werner, R.; Bunch, T.; Hiort, O. Androgen insensitivity syndrome. Semin. Reprod. Med. 2012, 30, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Luu-The, V.; Zhang, Y.; Poirier, D.; Labrie, F. Characteristics of human types 1, 2 and 3 17 beta-hydroxysteroid dehydrogenase activities: Oxidation/reduction and inhibition. J. Steroid Biochem. Mol. Biol. 1995, 55, 581–587. [Google Scholar] [CrossRef]
- Simpson, E.R. Genetic mutations resulting in estrogen insufficiency in the male. Mol. Cell. Endocrinol. 1998, 145, 55–59. [Google Scholar] [CrossRef]
- Lombardi, G.; Zarrilli, S.; Colao, A.; Paesano, L.; Di Somma, C.; Rossi, F.; De Rosa, M. Estrogens and health in males. Mol. Cell. Endocrinol. 2001, 178, 51–55. [Google Scholar] [CrossRef]
- Akingbemi, B.T. Estrogen regulation of testicular function. Reprod. Biol. Endocrinol. 2005, 3, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, K.M.; O’Donnell, L.; Jones, M.E.; Meachem, S.J.; Boon, W.C.; Fisher, C.R.; Graves, K.H.; McLachlan, R.I.; Simpson, E.R. Impairment of spermatogenesis in mice lacking a functional aromatase (cyp 19) gene. Proc. Natl. Acad. Sci. USA 1999, 96, 7986–7991. [Google Scholar] [CrossRef] [Green Version]
- Eddy, E.M.; Washburn, T.F.; Bunch, D.O.; Goulding, E.H.; Gladen, B.C.; Lubahn, D.B.; Korach, K.S. Targeted disruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis and infertility. Endocrinology 1996, 137, 4796–4805. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Tang, D.; Li, F.; Tian, H.; Yue, X.; Li, F.; Weng, X.; Sun, W.; Wang, W.; Mo, F. Supplementation with dietary linseed oil during peri-puberty stimulates steroidogenesis and testis development in rams. Theriogenology 2017, 102, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Hedia, M.G.; El-Belely, M.S.; Ismail, S.T.; Abo El-Maaty, A.M. Monthly changes in testicular blood flow dynamics and their association with testicular volume, plasma steroid hormones profile and semen characteristics in rams. Theriogenology 2019, 123, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Puranen, T.; Poutanen, M.; Ghosh, D.; Vihko, R.; Vihko, P. Origin of substrate specificity of human and rat 17beta-hydroxysteroid dehydrogenase type 1, using chimeric enzymes and site-directed substitutions. Endocrinology 1997, 138, 3532–3539. [Google Scholar] [CrossRef] [PubMed]
- Margiotti, K.; Kim, E.; Pearce, C.L.; Spera, E.; Novelli, G.; Reichardt, J.K. Association of the G289S single nucleotide polymorphism in the HSD17B3 gene with prostate cancer in Italian men. Prostate 2002, 53, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Moghrabi, N.; Hughes, I.A.; Dunaif, A.; Andersson, S. Deleterious missense mutations and silent polymorphism in the human 17beta-hydroxysteroid dehydrogenase 3 gene (HSD17B3). J. Clin. Endocrinol. Metab. 1998, 83, 2855–2860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engeli, R.T.; Tsachaki, M.; Hassan, H.A.; Sager, C.P.; Essawi, M.L.; Gad, Y.Z.; Kamel, A.K.; Mazen, I.; Odermatt, A. Biochemical Analysis of Four Missense Mutations in the HSD17B3 Gene Associated With 46,XY Disorders of Sex Development in Egyptian Patients. J. Sex. Med. 2017, 14, 1165–1174. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.S.; Uwada, J.; Hayashi, J.; Kikuya, K.-i.; Muranishi, Y.; Watanabe, H.; Yaegashi, K.; Hasegawa, K.; Ida, T.; Sato, T.; et al. Analyses of Molecular Characteristics and Enzymatic Activities of Ovine HSD17B3. Animals 2021, 11, 2876. https://doi.org/10.3390/ani11102876
Islam MS, Uwada J, Hayashi J, Kikuya K-i, Muranishi Y, Watanabe H, Yaegashi K, Hasegawa K, Ida T, Sato T, et al. Analyses of Molecular Characteristics and Enzymatic Activities of Ovine HSD17B3. Animals. 2021; 11(10):2876. https://doi.org/10.3390/ani11102876
Chicago/Turabian StyleIslam, Mohammad Sayful, Junsuke Uwada, Junki Hayashi, Kei-ichiro Kikuya, Yuki Muranishi, Hiroyuki Watanabe, Kazuhide Yaegashi, Kazuya Hasegawa, Takanori Ida, Takahiro Sato, and et al. 2021. "Analyses of Molecular Characteristics and Enzymatic Activities of Ovine HSD17B3" Animals 11, no. 10: 2876. https://doi.org/10.3390/ani11102876
APA StyleIslam, M. S., Uwada, J., Hayashi, J., Kikuya, K.-i., Muranishi, Y., Watanabe, H., Yaegashi, K., Hasegawa, K., Ida, T., Sato, T., Imamichi, Y., Kitano, T., Miyashiro, Y., Khan, R. I., Takahashi, S., Umezawa, A., Suzuki, N., Sekiguchi, T., & Yazawa, T. (2021). Analyses of Molecular Characteristics and Enzymatic Activities of Ovine HSD17B3. Animals, 11(10), 2876. https://doi.org/10.3390/ani11102876