
Goji Berry (Lycium barbarum) Supplementation during Pregnancy Influences Insulin Sensitivity in Rabbit Does but Not in Their Offspring



 , , ,
, , , 
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
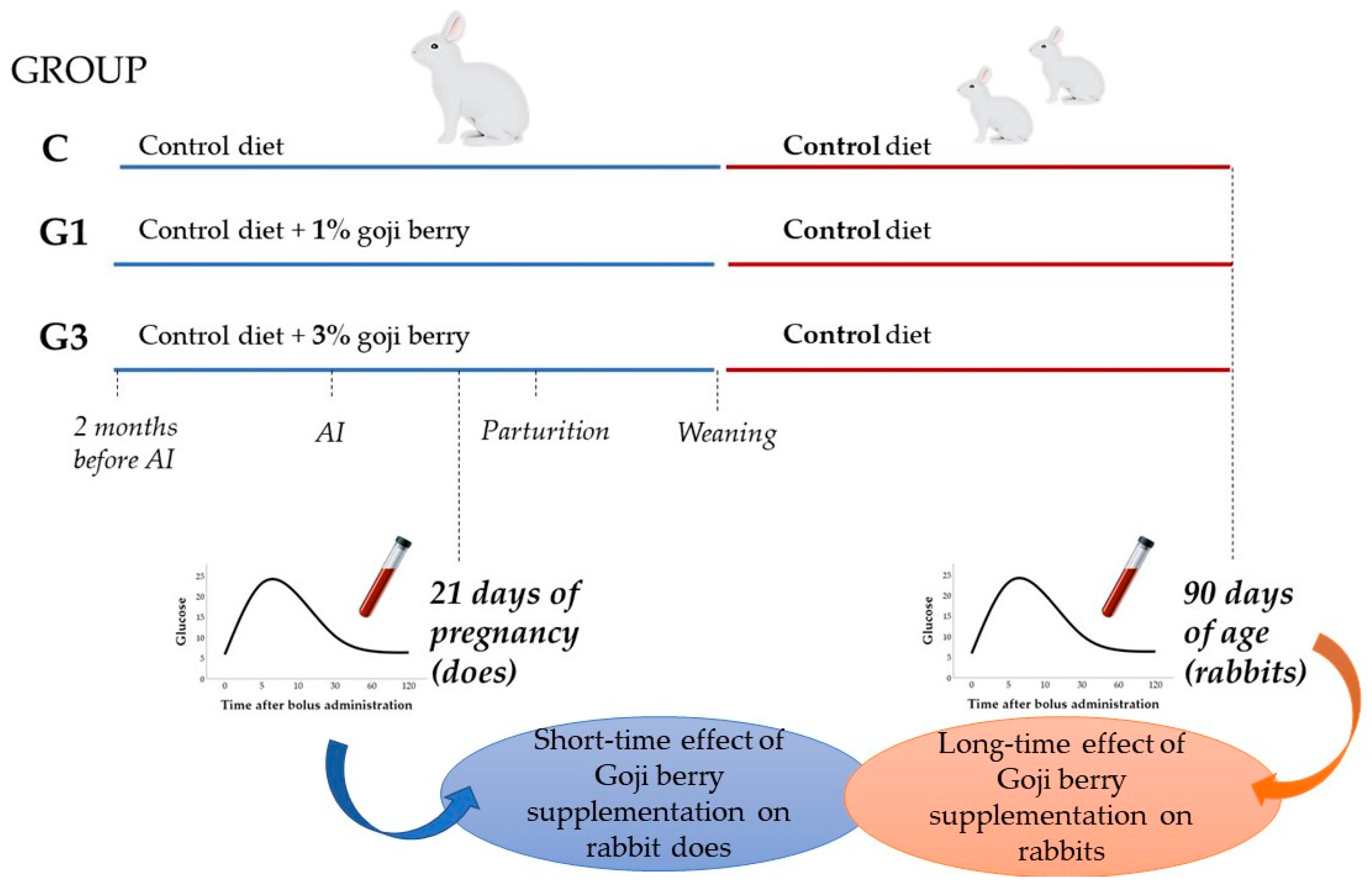
2.1. Animals and Experimental Design
2.2. Indices of Insulin Resistance
2.2.1. Simple Insulin Resistance Indices with Fasting Sampling
2.2.2. Intravenous Glucose Tolerance Test and Determination of Kinetic Parameters of Glucose
2.3. Statistical Analysis
3. Results
3.1. Body Weight
3.2. Fasting Blood Sample-Based Indices
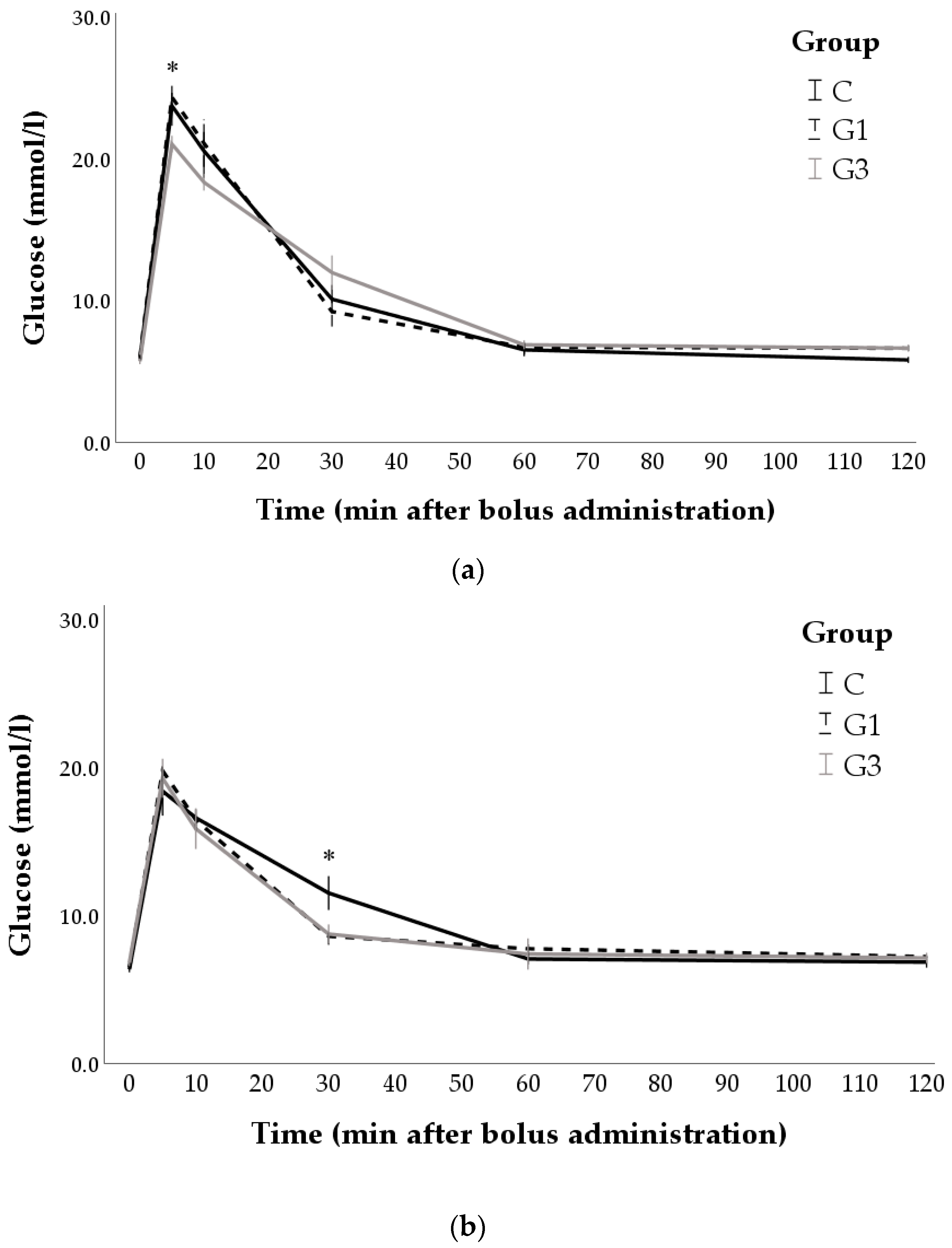
3.3. Changes after Bolus Administration and IVGTT Derived Indices
3.4. Correlations between Body Weight and Indices of Insulin Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American College of Physicians. In the clinic. Type 2 Diabetes. Ann. Intern. Med. 2010, 152, ITC31-15. [Google Scholar] [CrossRef]
- Franzago, M.; Fraticelli, F.; Stuppia, L.; Vitacolonna, E. Nutrigenetics, epigenetics and gestational diabetes: Consequences in mother and child. Epigenetics 2019, 14, 215–235. [Google Scholar] [CrossRef] [Green Version]
- Kampmann, U.; Knorr, S.; Fuglsang, J.; Ovesen, P. Determinants of Maternal Insulin Resistance during Pregnancy: An Updated Overview. J. Diabetes Res. 2019, 2019, 5320156. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M. Metabolic programming of long-term outcomes due to fatty acid nutrition in early life. Matern. Child Nutr. 2011, 7, 112–123. [Google Scholar] [CrossRef]
- Meruvu, S.; Schutz, L.F.; Choudhury, M. Nutritional influence on miRNA epigenetic regulation. In Molecular Nutrition: Mother and Infant; Academic Press: Cambridge, MA, USA, 2021; pp. 401–420. [Google Scholar]
- Barker, D.J.P.; Eriksson, J.G.; Forsén, T.; Osmond, C. Fetal origins of adult disease: Strength of effects and biological basis. Int. J. Epidemiol. 2002, 31, 1235–1239. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, K.; Rana, M.B.M.; Sultan, S. Oral Hypoglycemic Medications. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Potterat, O. Goji (Lycium barbarum and L. chinense): Phytochemistry, pharmacology and safety in the perspective of traditional uses and recent popularity. Planta Med. 2010, 76, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwok, S.S.; Bu, Y.; Lo, A.C.Y.; Chan, T.C.Y.; So, K.F.; Lai, J.S.M.; Shih, K.C. A Systematic Review of Potential Therapeutic Use of Lycium barbarum Polysaccharides in Disease. BioMed Res. Int. 2019, 2019, 4615745. [Google Scholar] [CrossRef]
- Masci, A.; Carradori, S.; Casadei, M.A.; Paolicelli, P.; Petralito, S.; Ragno, R.; Cesa, S. Lycium barbarum polysaccharides: Extraction, purification, structural characterisation and evidence about hypoglycaemic and hypolipidaemic effects. A review. Food Chem. 2018, 254, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wei, Y.; Wang, Y.; Gao, F.; Chen, Z. Lycium barbarum: A traditional Chinese herb and a promising anti-aging agent. Aging Dis. 2017, 8, 778–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pop, C.; Berce, C.; Ghibu, S.; Scurtu, I.; Soritău, O.; Login, C.; Kiss, B.; Stefan, M.G.; Fizesan, I.; Silaghi, H.; et al. Effects of Lycium barbarum L. Polysaccharides on inflammation and oxidative stress markers in a pressure overload-induced heart failure rat model. Molecules 2020, 25, 466. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Xiao, J.; Peng, B.; Xing, F.; So, K.-F.; Tipoe, G.L.; Lin, B. Retinal structure and function preservation by polysaccharides of wolfberry in a mouse model of retinal degeneration. Sci. Rep. 2014, 4, 7601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.; Qiu, B.; Li, Q.; Zhang, T.; Zhao, H.; Chen, Z.; Cai, Y.; Ruan, H.; Ge, W.; Zheng, X. LBP-4a improves insulin resistance via translocation and activation of GLUT4 in OLETF rats. Food Funct. 2014, 5, 811–820. [Google Scholar] [CrossRef]
- Ceccarini, M.R.; Vannini, S.; Cataldi, S.; Moretti, M.; Villarini, M.; Fioretti, B.; Albi, E.; Beccari, T.; Codini, M. Effect of Lycium barbarum berries cultivated in Umbria (Italy) on human hepatocellular carcinoma cells. J. Biotechnol. 2016, 231, S26–S27. [Google Scholar] [CrossRef]
- Nardi, G.M.; Januário, A.G.F.; Freire, C.G.; Megiolaro, F.; Schneider, K.; Perazzoli, M.R.A.; Do Nascimento, S.R.; Gon, A.C.; Mariano, L.N.B.; Wagner, G.; et al. Anti-inflammatory activity of berry fruits in mice model of inflammation is based on oxidative stress modulation. Pharmacogn. Res. 2016, 8, 42–49. [Google Scholar]
- Lee, S.R.; Hwang, H.J.; Yoon, J.G.; Bae, E.Y.; Goo, K.S.; Cho, S.J.; Cho, J.A. Anti-inflammatory effect of Lycium barbarum on polarized human intestinal epithelial cells. Nutr. Res. Pract. 2019, 13, 95–104. [Google Scholar] [CrossRef]
- You, J.; Chang, Y.; Zhao, D.; Zhuang, J.; Zhuang, W. A mixture of functional complex extracts from Lycium barbarum and grape seed enhances immunity synergistically in vitro and in vivo. J. Food Sci. 2019, 84, 1577–1585. [Google Scholar] [CrossRef]
- Kang, Y.; Xue, Y.; Du, M.; Zhu, M.J. Preventive effects of Goji berry on dextran-sulfate-sodium-induced colitis in mice. J. Nutr. Biochem. 2017, 40, 70–76. [Google Scholar] [CrossRef]
- Ruíz-Salinas, A.K.; Vázquez-Roque, R.A.; Díaz, A.; Pulido, G.; Treviño, S.; Floran, B.; Flores, G. The treatment of Goji berry (Lycium barbarum) improves the neuroplasticity of the prefrontal cortex and hippocampus in aged rats. J. Nutr. Biochem. 2020, 83, 108416. [Google Scholar] [CrossRef] [PubMed]
- Menchetti, L.; Vecchione, L.; Filipescu, I.; Petrescu, V.F.; Fioretti, B.; Beccari, T.; Ceccarini, M.R.; Codini, M.; Quattrone, A.; Trabalza-Marinucci, M.; et al. Effects of Goji berries supplementation on the productive performance of rabbit. Livest. Sci. 2019, 220, 123–128. [Google Scholar] [CrossRef]
- Lei, X.; Huo, P.; Wang, Y.; Xie, Y.; Shi, Q.; Tu, H.; Yao, J.; Mo, Z.; Zhang, S. Lycium barbarum Polysaccharides Improve Testicular Spermatogenic Function in Streptozotocin-Induced Diabetic Rats. Front. Endocrinol. 2020, 11, 164. [Google Scholar] [CrossRef]
- Cai, H.; Liu, F.; Zuo, P.; Huang, G.; Song, Z.; Wang, T.; Lu, H.; Guo, F.; Han, C.; Sun, G. Practical application of antidiabetic efficacy of Lycium barbarum polysaccharide in patients with type 2 diabetes. Med. Chem. 2015, 11, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Du, M.; Hu, X.; Kou, L.; Zhang, B.; Zhang, C. Lycium barbarum Polysaccharide Mediated the Antidiabetic and Antinephritic Effects in Diet-Streptozotocin-Induced Diabetic Sprague Dawley Rats via Regulation of NF-κB. BioMed Res. Int. 2016, 2016, 3140290. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Cai, Y.; Yan, J.; Sun, M.; Corke, H. Hypoglycemic and hypolipidemic effects and antioxidant activity of fruit extracts from Lycium barbarum. Life Sci. 2004, 76, 137–149. [Google Scholar] [CrossRef]
- Jing, L.; Cui, G.; Feng, Q.; Xiao, Y. Evaluation of hypoglycemic activity of the polysaccharides extracted from Lycium barbarum. Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; He, H.; Ji, H.; Willis, J.; Willard, L.; Jiang, Y.; Medeiros, D.M.; Wark, L.; Han, J.; Liu, Y.; et al. Wolfberries potentiate mitophagy and enhance mitochondrial biogenesis leading to prevention of hepatic steatosis in obese mice: The role of AMP-activated protein kinase α2 subunit. Mol. Nutr. Food Res. 2014, 58, 1005–1015. [Google Scholar] [CrossRef]
- Ming, M.; Guanhua, L.; Zhanhai, Y.; Guang, C.; Xuan, Z. Effect of the Lycium barbarum polysaccharides administration on blood lipid metabolism and oxidative stress of mice fed high-fat diet in vivo. Food Chem. 2009, 113, 872–877. [Google Scholar] [CrossRef]
- Menchetti, L.; Brecchia, G.; Branciari, R.; Barbato, O.; Fioretti, B.; Codini, M.; Bellezza, E.; Trabalza-Marinucci, M.; Miraglia, D. The effect of Goji berries (Lycium barbarum) dietary supplementation on rabbit meat quality. Meat Sci. 2020, 161, 108018. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, J.; Yan, J.; Liu, S.; Guo, Y.; Chen, D.; Luo, Q. Lycium barbarum polysaccharides ameliorates renal injury and inflammatory reaction in alloxan-induced diabetic nephropathy rabbits. Life Sci. 2016, 157, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, S.; Morimoto, M.; Liu, E.; Koike, T.; Higaki, Y.; Taura, Y.; Mamba, K.; Itamoto, K.; Watanabe, T.; Tsutsumi, K.; et al. Overexpression of lipoprotein lipases improves insulin resistance induced by a high-fat diet in transgenic rabbits. Diabetologia 2004, 47, 1202–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, B.; Wang, X.; Yu, Y.; Waqar, A.B.; Yu, Q.; Koike, T.; Shiomi, M.; Liu, E.; Wang, Y.; Fan, J. High-fructose and high-fat diet-induced insulin resistance enhances atherosclerosis in Watanabe heritable hyperlipidemic rabbits. Nutr. Metab. 2015, 12, 30. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Ito, T.; Ohwada, K.; Mera, Y.; Matsushita, M.; Tomoike, H. Hereditary postprandial hypertriglyceridemic rabbit exhibits insulin resistance and central obesity: A novel model of metabolic syndrome. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2752–2757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.B.; Nasir, A.; Prabhu, K.M.; Murthy, P.S.; Dev, G. Hypoglycaemic and hypolipidemic effect of ethanolic extract of seeds of Eugenia jambolana in alloxan-induced diabetic rabbits. J. Ethnopharmacol. 2003, 85, 201–206. [Google Scholar] [CrossRef]
- Dastmalchi, S.; Garjani, A.; Maleki, N.; Sheikhee, G.; Baghchevan, V.; Jafari-Azad, P.; Valizadeh, H.; Barzegar-Jalali, M. Enhancing dissolution, serum concentrations and hypoglycemic effect of glibenclamide using solvent deposition technique. J. Pharm. Pharm. Sci. 2005, 8, 175–181. [Google Scholar]
- Martínez-Paredes, E.; Ródenas, L.; Martínez-Vallespín, B.; Cervera, C.; Blas, E.; Brecchia, G.; Boiti, C.; Pascual, J.J. Effects of feeding programme on the performance and energy balance of nulliparous rabbit does. Animal 2012, 6, 1086–1095. [Google Scholar] [CrossRef] [Green Version]
- Menchetti, L.; Brecchia, G.; Cardinali, R.; Polisca, A.; Boiti, C. Feed restriction during pregnancy: Effects on body condition and productive performance of primiparous rabbit does. World Rabbit Sci. 2015, 23, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Boiti, C.; Guelfi, G.; Zerani, M.; Zampini, D.; Brecchia, G.; Gobbetti, A. Expression patterns of cytokines, p53 and nitric oxide synthase isoenzymes in corpora lutea of pseudopregnant rabbits during spontaneous luteolysis. Reproduction 2004, 127, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Menchetti, L.; Barbato, O.; Filipescu, I.E.; Traina, G.; Leonardi, L.; Polisca, A.; Troisi, A.; Guelfi, G.; Piro, F.; Brecchia, G. Effects of local lipopolysaccharide administration on the expression of Toll-like receptor 4 and pro-inflammatory cytokines in uterus and oviduct of rabbit does. Theriogenology 2018, 107, 162–174. [Google Scholar] [CrossRef]
- Brecchia, G.; Menchetti, L.; Cardinali, R.; Castellini, C.; Polisca, A.; Zerani, M.; Maranesi, M.; Boiti, C. Effects of a bacterial lipopolysaccharide on the reproductive functions of rabbit does. Anim. Reprod. Sci. 2014, 147, 128–134. [Google Scholar] [CrossRef]
- Denton, K.M.; Flower, R.L.; Stevenson, K.M.; Anderson, W.P. Adult rabbit offspring of mothers with secondary hypertension have increased blood pressure. Hypertension 2003, 41, 634–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menchetti, L.; Brecchia, G.; Canali, C.; Cardinali, R.; Polisca, A.; Zerani, M.; Boiti, C. Food restriction during pregnancy in rabbits: Effects on hormones and metabolites involved in energy homeostasis and metabolic programming. Res. Vet. Sci. 2015, 98, 7–12. [Google Scholar] [CrossRef]
- Menchetti, L.; Andoni, E.; Barbato, O.; Canali, C.; Quattrone, A.; Vigo, D.; Codini, M.; Curone, G.; Brecchia, G. Energy homeostasis in rabbit does during pregnancy and pseudopregnancy. Anim. Reprod. Sci. 2020, 218, 106505. [Google Scholar] [CrossRef] [PubMed]
- Menchetti, L.; Canali, C.; Castellini, C.; Boiti, C.; Brecchia, G. The different effects of linseed and fish oil supplemented diets on insulin sensitivity of rabbit does during pregnancy. Res. Vet. Sci. 2018, 118, 126–133. [Google Scholar] [CrossRef]
- Menchetti, L.; Curone, G.; Andoni, E.; Barbato, O.; Troisi, A.; Fioretti, B.; Polisca, A.; Codini, M.; Canali, C.; Vigo, D.; et al. Impact of goji berries (Lycium barbarum) supplementation on the energy homeostasis of rabbit does: Uni- and multivariate approach. Animals 2020, 10, 2000. [Google Scholar] [CrossRef] [PubMed]
- De Blas, C.; Mateos, G. Nutrition of the Rabbit; De Blas, C., Wiseman, J., Eds.; CABI: Oxon, UK, 1998; ISBN 9781845936693. [Google Scholar]
- Andoni, E.; Curone, G.; Agradi, S.; Barbato, O.; Menchetti, L.; Vigo, D.; Zelli, R.; Cotozzolo, E.; Ceccarini, M.R.; Faustini, M.; et al. Effect of Goji Berry (Lycium barbarum) Supplementation on Reproductive Performance of Rabbit Does. Animals 2021, 11, 1672. [Google Scholar] [CrossRef]
- Castrica, M.; Menchetti, L.; Balzaretti, C.C.M.; Branciari, R.; Ranucci, D.; Cotozzolo, E.; Vigo, D.; Curone, G.; Brecchia, G.; Miraglia, D. Impact of dietary supplementation with goji berries (Lycium barbarum) on microbiological quality, physico-chemical, and sensory characteristics of rabbit meat. Foods 2020, 9, 148. [Google Scholar] [CrossRef]
- Maertens, L.; Moermans, R.; De Groote, G. Prediction of the apparent digestible energy content of commercial pelleted feeds for rabbits. J. Appl. Rabbit Res. 1988, 11, 60–67. [Google Scholar]
- Brecchia, G.; Bonanno, A.; Galeati, G.; Federici, C.; Maranesi, M.; Gobbetti, A.; Zerani, M.; Boiti, C. Hormonal and metabolic adaptation to fasting: Effects on the hypothalamic-pituitary-ovarian axis and reproductive performance of rabbit does. Domest. Anim. Endocrinol. 2006, 31, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Borai, A.; Livingstone, C.; Kaddam, I.; Ferns, G. Selection of the appropriate method for the assessment of insulin resistance. BMC Med. Res. Methodol. 2011, 11, 158. [Google Scholar] [CrossRef] [Green Version]
- Rommers, J.M.; Boiti, C.; Brecchia, G.; Meijerhof, R.; Noordhuizen, J.P.T.M.; Decuypere, E.; Kemp, B. Metabolic adaptation and hormonal regulation in young rabbit does during long-term caloric restriction and subsequent compensatory growth. Anim. Sci. 2004, 79, 255–264. [Google Scholar] [CrossRef] [Green Version]
- García-García, R.M.; Rebollar, P.G.; Arias-Álvarez, M.; Sakr, O.G.; Bermejo-Álvarez, P.; Brecchia, G.; Gutierrez-Adan, A.; Zerani, M.; Boiti, C.; Lorenzo, P.L. Acute fasting before conception affects metabolic and endocrine status without impacting follicle and oocyte development and embryo gene expression in the rabbit. Reprod. Fertil. Dev. 2011, 23, 759–768. [Google Scholar] [CrossRef]
- Field, A. Discovering Statistics Using SPSS, 3rd ed.; SAGE Publications Ltd.: London, UK, 2009; Volume 81, ISBN 9781847879066. [Google Scholar]
- Yang, Y.; Li, W.; Li, Y.; Wang, Q.; Gao, L.; Zhao, J. Dietary Lycium barbarum polysaccharide induces Nrf2/ARE pathway and ameliorates insulin resistance induced by high-fat via activation of PI3K/AKT signaling. Oxid. Med. Cell. Longev. 2014, 2014, 145641. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Tang, H.; Wang, F.; Yang, X.; Wang, Z.; Liu, H.; Pan, D.; Yang, C.; Wang, S.; Sun, G. An untargeted metabolomics approach reveals further insights of Lycium barbarum polysaccharides in high fat diet and streptozotocin-induced diabetic rats. Food Res. Int. 2019, 116, 20–29. [Google Scholar] [CrossRef]
- Barker, J.M.; McFann, K.; Harrison, L.C.; Fourlanos, S.; Krischer, J.; Cuthbertson, D.; Chase, H.P.; Eisenbarth, G.S. Pre-Type 1 Diabetes Dysmetabolism: Maximal Sensitivity Achieved with Both Oral and Intravenous Glucose Tolerance Testing. J. Pediatr. 2007, 150, 31–36.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meshkani, R.; Adeli, K. Hepatic insulin resistance, metabolic syndrome and cardiovascular disease. Clin. Biochem. 2009, 42, 1331–1346. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Montgomery, B.; Howell, W.; Ligueros-Saylan, M.; Hsu, C.; Devineni, D.; McLeod, J.; Horowitz, A.; Foley, J. Importance of Early Phase Insulin Secretion to Intravenous Glucose Tolerance in Subjects with Type 2 Diabetes Mellitus. J. Clin. Endocrinol. Metab. 2001, 86, 5824–5829. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.J. Carbohydrate Metabolism and Its Diseases. In Clinical Biochemistry of Domestic Animals; Academic Press: Cambridge, MA, USA, 1997; pp. 45–81. [Google Scholar]
- Lacroix, M.; Kina, E.; Hivert, M.F. Maternal/fetal determinants of insulin resistance in women during pregnancy and in offspring over life. Curr. Diabetes Rep. 2013, 13, 238–244. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Diet | |||
|---|---|---|---|
| C | G1 | G3 | |
| Ingredients 1 | |||
| Wheat bran | 30.0 | 29.5 | 29.0 |
| Dehydrated alfalfa meal | 42.0 | 41.5 | 41.0 |
| Barley | 9.5 | 9.5 | 9.0 |
| Sunflower meal | 4.5 | 4.5 | 4.2 |
| Rice bran | 4.0 | 4.0 | 3.9 |
| Soybean meal | 4.0 | 4.0 | 3.9 |
| Calcium carbonate | 2.2 | 2.2 | 2.2 |
| Cane molasses | 2.0 | 2.0 | 2.0 |
| Dicalcium phosphate | 0.7 | 0.7 | 0.7 |
| Vitamin–mineral premix 2 | 0.4 | 0.4 | 0.4 |
| Soybean oil | 0.4 | 0.4 | 0.4 |
| Salt | 0.3 | 0.3 | 0.3 |
| Goji berries | - | 1.0 | 3.0 |
| Analytical data 1 | |||
| Crude protein | 15.74 | 15.64 | 15.66 |
| Ether extract | 2.25 | 2.23 | 2.47 |
| Ash | 9.28 | 9.36 | 9.25 |
| Starch | 16.86 | 17.07 | 16.99 |
| NDF | 38.05 | 38.55 | 37.49 |
| ADF | 19.54 | 19.60 | 19.01 |
| ADL | 4.01 | 4.31 | 3.98 |
| Digestible Energy 3 | 2464 | 2463 | 2459 |
| Animal Category | Parameter | Group | RMSE | p-Value | ||
|---|---|---|---|---|---|---|
| C | G1 | G3 | ||||
| Pregnant does | Fasting glucose (mmol/L) | 5.88 | 6.01 | 5.73 | 0.48 | 0.457 |
| Fasting insulin (μU/mL) | 7.32 | 6.50 | 5.86 | 3.30 | 0.648 | |
| Glucose-to-insulin ratio * | 0.96 | 1.25 | 1.30 | 0.67 | 0.632 | |
| HOMA-IR * | 0.11 | 0.09 | 0.09 | 0.01 | 0.567 | |
| 90-day-old rabbits | Fasting glucose (mmol/L) | 6.36 | 6.50 | 6.80 | 0.61 | 0.271 |
| Fasting insulin (μU/mL) | 7.47 | 8.03 | 9.32 | 1.86 | 0.094 | |
| Glucose-to-insulin ratio * | 0.92 | 0.86 | 0.73 | 0.21 | 0.594 | |
| HOMA-IR * | 0.12 b | 0.13 a,b | 0.16 a | 0.01 | 0.037 | |
| Animal Category | Parameter | Group | RMSE | p-Value | ||
|---|---|---|---|---|---|---|
| C | G1 | G3 | ||||
| Pregnant does | Cmax (mmol/L) | 23.80 a | 24.91 a | 20.84 b | 2.21 | 0.019 |
| Glucose 60 min (mmol/L) | 6.53 | 6.72 | 6.89 | 0.96 | 0.862 | |
| AUC ((mmol/L) × min) | 1111.93 | 1133.50 | 1158.60 | 115.84 | 0.802 | |
| Kel (%/min) | 1.03 a | 1.05 a | 0.86 b | 0.10 | 0.018 | |
| t1/2 (min) | 68.52 B | 66.85 B | 79.11 A | 7.75 | 0.047 | |
| Vd (dl/kg) | 1.46 a,b | 1.37 b | 1.59 a | 0.14 | 0.023 | |
| CL (dl/min/kg) | 1.45 | 1.43 | 1.39 | 0.18 | 0.837 | |
| 90-day-old rabbits | Cmax (mmol/L) | 18.81 | 18.51 | 19.26 | 2.92 | 0.904 |
| Glucose 60 min (mmol/L) | 7.06 | 7.76 | 7.40 | 1.59 | 0.794 | |
| AUC ((mmol/L) × min) | 1126.28 | 1101.67 | 1076.77 | 110.70 | 0.745 | |
| Kel (%/min) | 0.75 | 0.74 | 0.76 | 0.15 | 0.959 | |
| t1/2 (min) | 93.73 | 97.13 | 85.10 | 13.00 | 0.326 | |
| Vd (dl/kg) | 1.91 | 1.71 | 1.71 | 0.31 | 0.681 | |
| CL (dl/min/kg) | 1.41 | 1.25 | 1.31 | 0.32 | 0.732 | |
| Parameter | Animal Category | |
|---|---|---|
| Pregnant Does | 90-Day-Old Rabbits | |
| Fasting glucose | 0.022 | 0.281 |
| Fasting insulin | −0.137 | −0.054 |
| Glucose–insulin ratio # | 0.109 | 0.166 |
| HOMA | −0.105 | 0.024 |
| Cmax | 0.153 | 0.060 |
| Glucose 60 min | 0.158 | 0.165 |
| AUC | 0.539 * | 0.248 |
| Kel | −0.116 | 0.071 |
| t1/2 | −0.033 | 0.254 |
| Vd # | −0.266 | −0.290 |
| CL | −0.202 | −0.119 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brecchia, G.; Sulce, M.; Curone, G.; Barbato, O.; Canali, C.; Troisi, A.; Munga, A.; Polisca, A.; Agradi, S.; Ceccarini, M.R.; et al. Goji Berry (Lycium barbarum) Supplementation during Pregnancy Influences Insulin Sensitivity in Rabbit Does but Not in Their Offspring. Animals 2022, 12, 39. https://doi.org/10.3390/ani12010039
Brecchia G, Sulce M, Curone G, Barbato O, Canali C, Troisi A, Munga A, Polisca A, Agradi S, Ceccarini MR, et al. Goji Berry (Lycium barbarum) Supplementation during Pregnancy Influences Insulin Sensitivity in Rabbit Does but Not in Their Offspring. Animals. 2022; 12(1):39. https://doi.org/10.3390/ani12010039
Chicago/Turabian StyleBrecchia, Gabriele, Majlind Sulce, Giulio Curone, Olimpia Barbato, Claudio Canali, Alessandro Troisi, Albana Munga, Angela Polisca, Stella Agradi, Maria Rachele Ceccarini, and et al. 2022. "Goji Berry (Lycium barbarum) Supplementation during Pregnancy Influences Insulin Sensitivity in Rabbit Does but Not in Their Offspring" Animals 12, no. 1: 39. https://doi.org/10.3390/ani12010039
APA StyleBrecchia, G., Sulce, M., Curone, G., Barbato, O., Canali, C., Troisi, A., Munga, A., Polisca, A., Agradi, S., Ceccarini, M. R., Vigo, D., Quattrone, A., Draghi, S., & Menchetti, L. (2022). Goji Berry (Lycium barbarum) Supplementation during Pregnancy Influences Insulin Sensitivity in Rabbit Does but Not in Their Offspring. Animals, 12(1), 39. https://doi.org/10.3390/ani12010039