
Evaluation of the Respiratory Microbiome and the Use of Tracheal Lavage as a Diagnostic Tool in Kemp’s Ridley Sea Turtles (Lepidochelys kempii)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Culture Methods
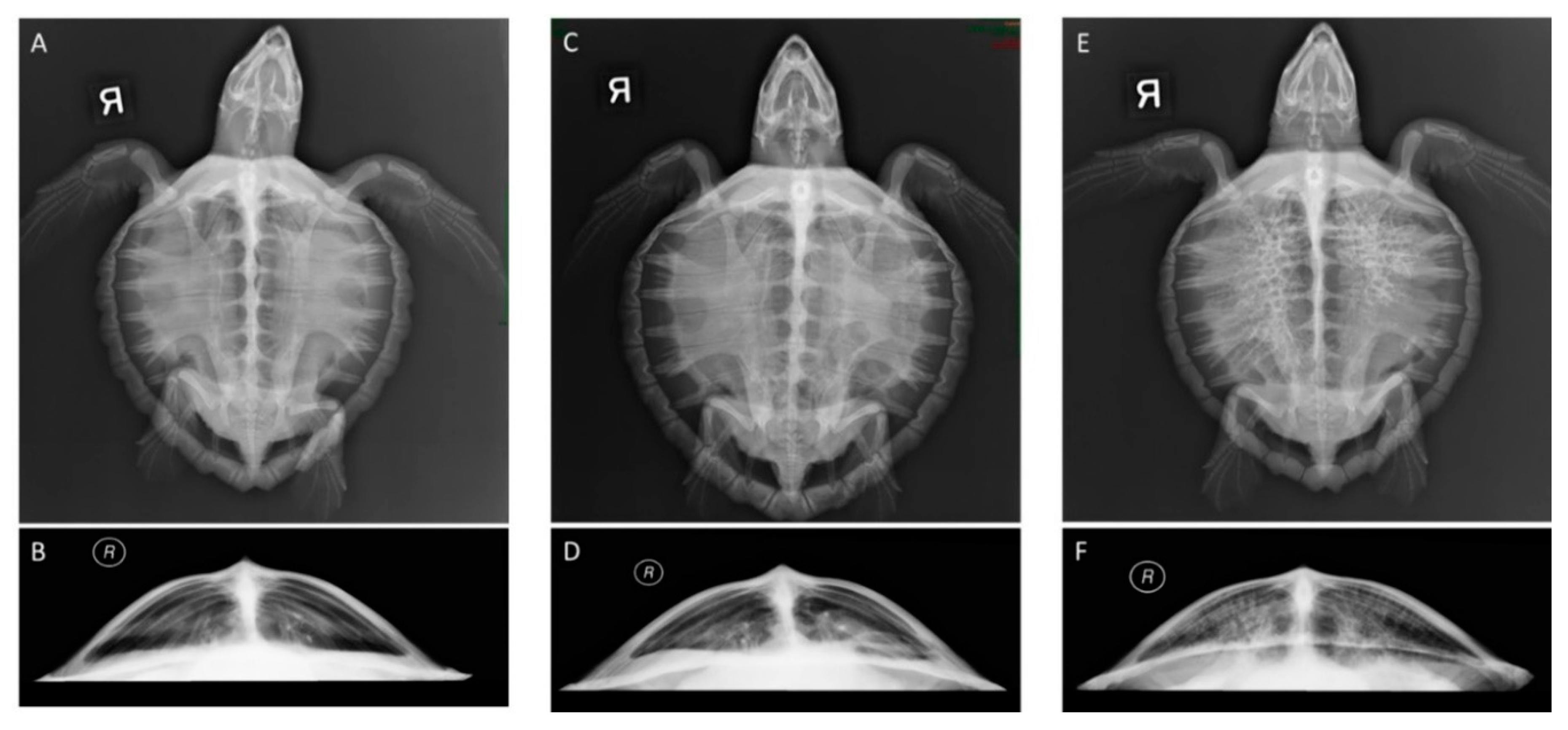
2.3. Retrospective Radiographic Review and Scoring System
2.4. DNA Extraction and Sequencing
2.5. Data Analysis
3. Results
3.1. Sample Data
3.2. The Tracheal Lavage Microbiome
3.3. Comparison of Culture-Independent and Culture-Dependent Methods
3.4. Previously Reported Sea Turtle Pathogens in Sequence Data
3.5. Necropsy Samples
4. Discussion
4.1. The Respiratory Microbiome and Pneumonia
4.2. The Tracheal Lavage Microbiome over Time
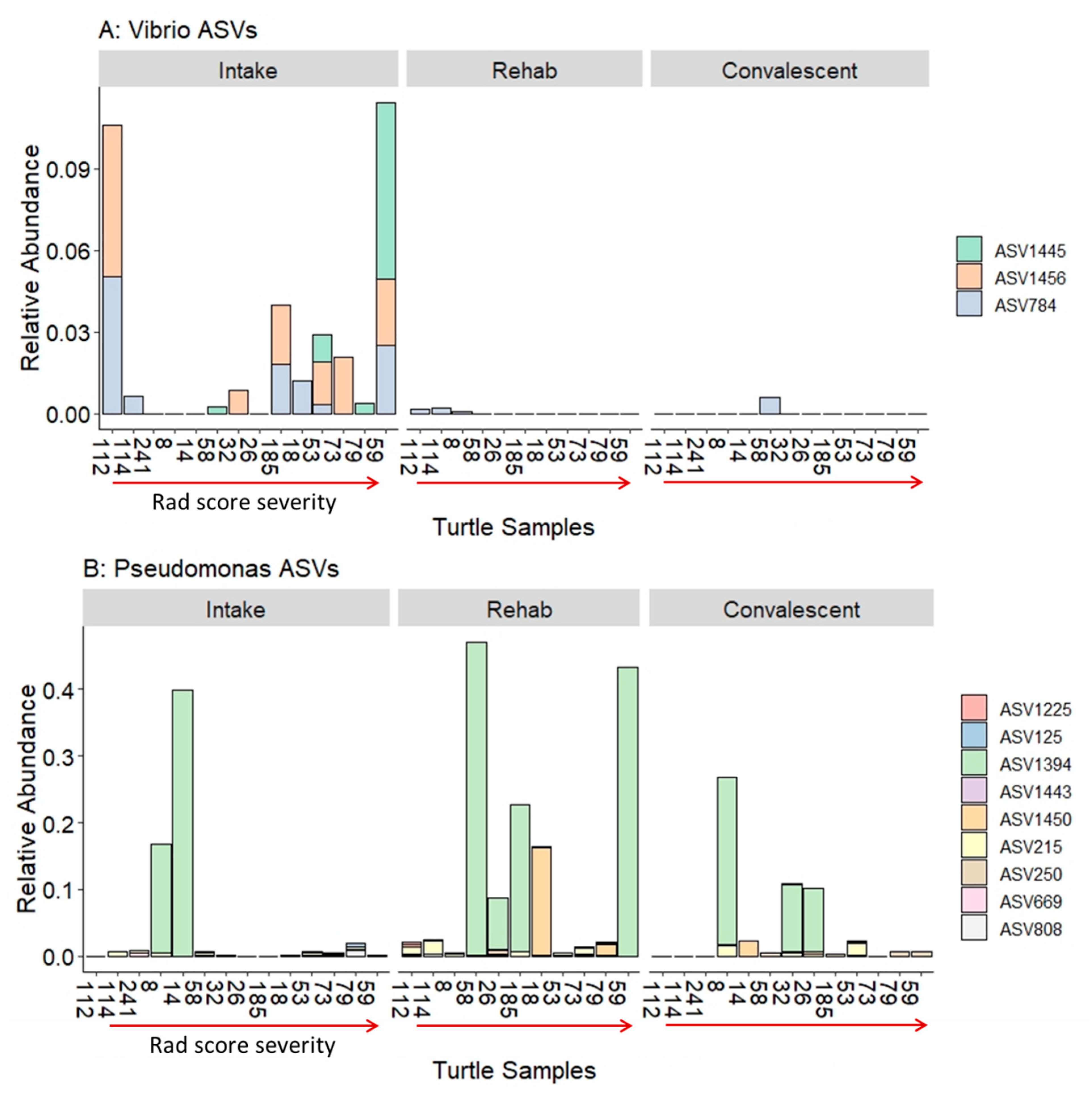
4.3. The Tracheal Lavage Microbiome and Degree of Radiographic Lung Abnormalities
4.4. Culture-Dependent vs. Culture-Independent Methods
4.5. Screening of Previously Reported Pathogens in the Microbiome
4.6. Regionality of the Respiratory Tract Microbiome
4.7. Tracheal Lavage as a Diagnostic Tool
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boylan, S.M.; Valente, A.L.S.; Innis, C.J.; Stacy, B.A.; Wyneken, J. Respiratory System. In Sea Turtle Health and Rehabilitation; Manire, C.A., Norton, T.M., Stacy, B.A., Innis, C.J., Harms, C.A., Eds.; J. Ross Publishing: Plantation, FL, USA, 2017; pp. 325–337. [Google Scholar]
- Stockman, J.; Innis, C.J.; Solano, M.; Brisson, J.O.; Kass, P.H.; Tlusty, M.F.; Weber, E.S. Prevalence, distribution, and progression of radiographic abnormalities in the lungs of cold-stunned Kemp’s ridley sea turtles (Lepidochelys kempii): 89 cases (2002–2005). J. Am. Vet. Med. Assoc. 2013, 242, 675–681. [Google Scholar] [CrossRef]
- Murray, M.J. Medical care of sea turtles. In Reptile Medicine and Surgery, 2nd ed.; Mader, D.M., Ed.; Elsevier: St. Louis, MO, USA, 2006; pp. 865–877. [Google Scholar]
- Dickson, R.P.; Erb-Downward, J.R.; Huffnagle, G.B. The role of the bacterial microbiome in lung disease. Expert Rev. Respir. Med. 2013, 7, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Dickson, R.P.; Erb-Downward, J.R.; Huffnagle, G.B. Towards an ecology of the lung: New conceptual models of pulmonary microbiology and pneumonia pathogenesis. Lancet Respir. Med. 2014, 2, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Dickson, R.P.; Erb-Downward, J.R.; Martinez, F.J.; Huffnagle, G.B. The Microbiome and the Respiratory Tract. Annu. Rev. Physiol. 2016, 78, 481–504. [Google Scholar] [CrossRef] [Green Version]
- Dickson, R.P.; Huffnagle, G.B. The Lung Microbiome: New Principles for Respiratory Bacteriology in Health and Disease. PLoS Pathog. 2015, 11, e1004923. [Google Scholar] [CrossRef]
- Martin, C.; Burgel, P.-R.; Lepage, P.; Andréjak, C.; De Blic, J.; Bourdin, A.; Brouard, J.; Chanez, P.; Dalphin, J.-C.; Deslee, G.; et al. Host–microbe interactions in distal airways: Relevance to chronic airway diseases. Eur. Respir. Rev. 2015, 24, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Ho Man, W.; de Steenhuijsen Piters, W.A.A.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Zaneveld, J.R.; McMinds, R.; Thurber, R.V. Stress and stability: Applying the Anna Karenina principle to animal microbiomes. Nat. Microbiol. 2017, 2, 17121. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.P.; Erb-Downward, J.R.; Prescott, H.; Martinez, F.J.; Curtis, J.; Lama, V.N.; Huffnagle, G.B. Analysis of Culture-Dependent versus Culture-Independent Techniques for Identification of Bacteria in Clinically Obtained Bronchoalveolar Lavage Fluid. J. Clin. Microbiol. 2014, 52, 3605–3613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritz, K. The Plate Debate: Cultivable communities have no utility in contemporary environmental microbial ecology. FEMS Microbiol. Ecol. 2007, 60, 358–362. [Google Scholar] [CrossRef] [Green Version]
- Stewart, E.J. Growing Unculturable Bacteria. J. Bacteriol. 2012, 194, 4151–4160. [Google Scholar] [CrossRef] [Green Version]
- Vientós-Plotts, A.I.; Ericsson, A.C.; Rindt, H.; Reinero, C.R. Respiratory Dysbiosis in Canine Bacterial Pneumonia: Standard Culture vs. Microbiome Sequencing. Front. Vet. Sci. 2019, 6, 354. [Google Scholar] [CrossRef] [PubMed]
- Abutarbush, S.M.; Al-Rukibat, R.K.; Qura’N, W.; Hananeh, W.M. Laboratory findings of tracheal wash and bronchoalveolar lavage in normal adult dairy cattle. J. Appl. Anim. Res. 2019, 47, 46–53. [Google Scholar] [CrossRef]
- Sumner, C.M.; Rozanski, E.A.; Sharp, C.R.; Shaw, S.P. The use of deep oral swabs as a surrogate for transoral tracheal wash to obtain bacterial cultures in dogs with pneumonia. J. Vet. Emerg. Crit. Care 2011, 21, 515–520. [Google Scholar] [CrossRef]
- Glendinning, L.; Wright, S.; Pollock, J.; Tennant, P.; Collie, D.; McLachlan, G. Variability of the Sheep Lung Microbiota. Appl. Environ. Microbiol. 2016, 82, 3225–3238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morreale, S.J.; Standora, E.A. Western North Atlantic waters: Crucial developmental habitat for Kemp’s ridley and loggerhead sea turtles. Chelonian Conserv. Biol. 2005, 4, 872–882. [Google Scholar]
- Still, B.M.; Griffin, C.R.; Prescott, R. Climatic and oceanographic factors affecting daily patterns of juvenile sea turtle cold-stunning in Cape Cod Bay, Massachusetts. Chelonian Conserv. Biol. 2005, 4, 883–890. [Google Scholar]
- Griffin, L.P.; Griffin, C.R.; Finn, J.T.; Prescott, R.L.; Faherty, M.; Still, B.M.; Danylchuk, A.J. Warming seas increase cold-stunning events for Kemp’s ridley sea turtles in the northwest Atlantic. PLoS ONE 2019, 14, e0211503. [Google Scholar] [CrossRef] [Green Version]
- Wibbels, T.; Bevan, E. Lepidochelys Kempii. The IUCN Red List of Threatened Species. 2019. e.T11533A142050590. Available online: http://dx.doi.org/10.2305/IUCN.UK.2019-2.RLTS.T11533A142050590.en (accessed on 10 September 2021).
- Innis, C.J.; Tlusty, M.; Merigo, C.; Weber, E.S. Metabolic and respiratory status of cold-stunned Kemp’s ridley sea turtles (Lepidochelys kempii). J. Comp. Physiol. B 2007, 177, 623–630. [Google Scholar] [CrossRef]
- Innis, C.; Nyaoke, A.C.; Williams, C.R.; Dunnigan, B.; Merigo, C.; Woodward, D.L.; Weber, E.S.; Frasca Jr, S. Pathologic and parasitologic findings of cold-stunned Kemp’s ridley sea turtles (Lepidochelys kempii) stranded on Cape Cod, Massachusetts, 2001–2006. J. Wildl. Dis. 2009, 45, 594–610. [Google Scholar] [CrossRef] [Green Version]
- Innis, C.J.; Ravich, J.B.; Tlusty, M.F.; Hoge, M.S.; Wunn, D.S.; Boerner-Neville, L.B.; Merigo, C.; Weber, E.S. Hematologic and plasma biochemical findings in cold-stunned Kemp’s ridley turtles: 176 cases (2001–2005). J. Am. Vet. Med. Assoc. 2009, 235, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Innis, C.J.; Braverman, H.; Cavin, J.M.; Ceresia, M.L.; Baden, L.R.; Kuhn, D.M.; Frasca, S.; McGowan, J.P.; Hirokawa, K.; Weber, E.S.; et al. Diagnosis and management of Enterococcus spp. infections during rehabilitation of cold-stunned Kemp’s ridley turtles (Lepidochelys kempii): 50 cases (2006–2012). J. Am. Vet. Med. Assoc. 2014, 245, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Innis, C.J.; Kennedy, A.; McGowan, J.P.; Buchweitz, J.P.; McNally, K. Glomerular Filtration Rates of Naturally Cold-Stunned Kemp’s Ridley Turtles (Lepidochelys kempii): Comparison of Initial vs. Convalescent Values. J. Herpetol. Med. Surg. 2016, 26, 100–103. [Google Scholar] [CrossRef]
- Keller, K.A.; Innis, C.J.; Tlusty, M.F.; Kennedy, A.E.; Bean, S.B.; Cavin, J.M.; Merigo, C. Metabolic and respiratory derangements associated with death in cold-stunned Kemp’s ridley turtles (Lepidochelys kempii): 32 cases (2005–2009). J. Am. Vet. Med. Assoc. 2012, 240, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, A.; Innis, C.J.; Rumbeiha, W. Determination of Glomerular Filtration Rate in Juvenile Kemp’s Ridley Turtles (Lepidochelys kempii) Using Iohexol Clearance, with Preliminary Comparison of Clinically Healthy Turtles vs. Those with Renal Disease. J. Herpetol. Med. Surg. 2012, 22, 25. [Google Scholar] [CrossRef]
- Innis, C.; Merigo, C.; Dodge, K.; Tlusty, M.; Dodge, M.; Sharp, B.; Myers, A.; McIntosh, A.; Wunn, D.; Perkins, C.; et al. Health Evaluation of Leatherback Turtles (Dermochelys coriacea) in the Northwestern Atlantic During Direct Capture and Fisheries Gear Disentanglement. Chelonian Conserv. Biol. 2010, 9, 205–222. [Google Scholar] [CrossRef]
- Wayne, P.A. Performance Standards for Antimicrobial Susceptibility Testing; Twentieth Informa-Tional Supplement M100–S20; Wayne, P., Ed.; Clinical and Laboratory Standards Institute: Annapolis Junction, MD, USA, 2010. [Google Scholar]
- Mettel, C.; Kim, Y.; Shrestha, P.M.; Liesack, W. Extraction of mRNA from Soil. Appl. Environ. Microbiol. 2010, 76, 5995–6000. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Eren, A.M.; Vineis, J.H.; Morrison, H.; Sogin, M.L. A Filtering Method to Generate High Quality Short Reads Using Illumina Paired-End Technology. PLoS ONE 2013, 8, e66643. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 10 September 2021).
- Murali, A.; Bhargava, A.; Wright, E.S. IDTAXA: A novel approach for accurate taxonomic classification of microbiome sequences. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Innis, C.J.; Frasca, S., Jr. Bacterial and Fungal Diseases. In Sea Turtle Health and Rehabilitation; Manire, C.A., Norton, T.M., Stacy, B.A., Innis, C.J., Harms, C.A., Eds.; J. Ross Publishing: Plantation, FL, USA, 2017; pp. 779–790. [Google Scholar]
- Faner, R.; Sibila, O.; Agustí, A.; Bernasconi, E.; Chalmers, J.D.; Huffnagle, G.B.; Manichanh, C.; Molyneaux, P.L.; Paredes, R.; Brocal, V.P.; et al. The microbiome in respiratory medicine: Current challenges and future perspectives. Eur. Respir. J. 2017, 49, 1602086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlson, E.S.; Bittinger, K.; Haas, A.R.; Fitzgerald, A.S.; Frank, I.; Yadav, A.; Bushman, F.; Collman, R.G. Topographical Continuity of Bacterial Populations in the Healthy Human Respiratory Tract. Am. J. Respir. Crit. Care Med. 2011, 184, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Dickson, R.P.; Erb-Downward, J.R.; Freeman, C.M.; McCloskey, L.; Falkowski, N.R.; Huffnagle, G.B.; Curtis, J.L. Bacterial Topography of the Healthy Human Lower Respiratory Tract. mBio 2017, 8, e02287-16. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, C.L.; Sandmeier, F.C.; Tracy, C.R. Host species, pathogens and disease associated with divergent nasal microbial communities in tortoises. R. Soc. Open Sci. 2018, 5, 181068. [Google Scholar] [CrossRef] [Green Version]
- Lima, N.; Rogers, T.; Acevedo-Whitehouse, K.; Brown, M.V. Temporal stability and species specificity in bacteria associated with the bottlenose dolphins respiratory system. Environ. Microbiol. Rep. 2011, 4, 89–96. [Google Scholar] [CrossRef]
- Wellehan, J.F.X.; Divers, S.J. Bacteriology. In Reptile Medicine and Surgery, 3rd ed.; Divers, S.J., Stahl, S.J., Eds.; Elsevier: St. Louis, MO, USA, 2019; pp. 235–247. [Google Scholar]
- Frans, I.; Michiels, C.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef]
- Gross, H.; Loper, J.E. Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep. 2009, 26, 1408–1446. [Google Scholar] [CrossRef]
- Tseng, S.-Y.; Liu, P.-Y.; Lee, Y.-H.; Wu, Z.-Y.; Huang, C.-C.; Cheng, C.-C.; Tung, K.-C. The Pathogenicity of Shewanella algae and Ability to Tolerate a Wide Range of Temperatures and Salinities. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jooste, P.; Hugo, C.J. The taxonomy, ecology and cultivation of bacterial genera belonging to the family Flavobacteriaceae. Int. J. Food Microbiol. 1999, 53, 81–94. [Google Scholar] [CrossRef]
- West, J.B. Regional Differences in the Lung. Chest 1978, 74, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Zeltner, T.B.; Bertacchini, M.; Messerli, A.; Burri, P.H. Morphometric Estimation of Regional Differences in the Rat Lung. Exp. Lung Res. 1990, 16, 145–158. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Description | Radiographic Features |
|---|---|---|
| 0 | Normal | No radiographic abnormalities of the pulmonary parenchyma |
| 1 | Minimal | Interstitial, increased opacity with ill-defined pulmonary vasculature |
| 2 | Mild | Diffuse interstitial |
| 3 | Mild-Moderate | Mixed interstitial and alveolar pattern, increased opacity with obscured pulmonary vasculature |
| 4 | Moderate | Alveolar focal or multifocal, mild reticular |
| 5 | Marked | Diffuse bilateral alveolar or marked reticular pattern |
| Intake | Rehab | Conv | |
|---|---|---|---|
| n | n | n | |
| Vibrio alginolyticus | 1 | 1 | 0 |
| Pseudomonas sp. | 0 | 2 | 0 |
| Raoultella planticola | 1 | 0 | 0 |
| Vibrio sp. | 2 | 3 | 3 |
| Shewanella putrefaciens | 1 | 0 | 0 |
| Enterococcus sp. | 0 | 1 | 0 |
| Non-enteric gram neg rod | 3 | 2 | 1 |
| No growth | 2 | 2 | 4 |
| ASV | Genus | BLAST Matches |
|---|---|---|
| ASV784 | Vibrio | chagasii, cuclitrophicus, splendidus |
| ASV1445 | Vibrio | anguillarum, cortegadensis |
| ASV1456 | Vibrio | tapetis |
| ASV1225 | Pseudomonas | graminis, viridiflava, donghuensis |
| ASV125 | Pseudomonas | marginalis, grimontii, rhodesiae |
| ASV1394 | Pseudomonas | flourescens, veronii |
| ASV1443 | Pseudomonas | koreensis |
| ASV1450 | Pseudomonas | pachastrellae, aestusnigri |
| ASV215 | Pseudomonas | oleovorans, stutzeri, taeanensis |
| ASV250 | Pseudomonas | arsenicoxydans, yamanorum, proteolytica |
| ASV669 | Pseudomonas | paralactis, tolaasii, gessardii |
| ASV808 | Pseudomonas | entomophila, mosselii, guariconensis |
| ASV1096 | Shewanella | marisflavi, fidelis, schlegeliana |
| ASV1267 | Shewanella | algae |
| ASV53 | Shewanella | woodyi |
| Bacteria | Genus Prevalence % | Genus SD % | Number of ASVs in Genus | Family | Family Prevalence % | Family SD % | Number of ASVs in Family |
|---|---|---|---|---|---|---|---|
| Aeromonas hydrophila | 0 | 0 | 0 | Aeromonadaceae | 0 | 0 | 0 |
| Bacillus sp. | 0 | 0 | 0 | Bacillaceae | 1.3 | 3.5 | 7 |
| Corynebacterium sp. | 1.7 | 2.1 | 8 | Corynebacteriaceae | 2.1 | 2.4 | 13 |
| Hafnia alvei | 0 | 0 | 0 | Enterobacteriaceae | 0.35 | 0.43 | 2 |
| Citrobacter braakii | 0 | 0 | 0 | Enterobacteriaceae | |||
| Citrobacter freundii | 0 | 0 | 0 | Enterobacteriaceae | |||
| Escherichia coli | 0 | 0 | 0 | Enterobacteriaceae | |||
| Enterococcus spp. | 0 | 0 | 0 | Enterococcaceae | 0.09 | 0.16 | 1 |
| Flavobacterium sp. | 0.07 | 0.12 | 1 | Flavobacteriaceae | 8.82 | 6.45 | 48 |
| Acinetobacter calcoaceticus | 0.66 | 0.7 | 5 | Moraxellaceae | 1 | 0.9 | 10 |
| Chromobacterium violaceum | 0 | 0 | 0 | Neisseriaceae | 0.17 | 0.33 | 4 |
| Pseudomonas sp. | 5.10 | 12.3 | 7 | Pseudomonadaceae | 5.15 | 12.2 | 8 |
| Shewanella algae | 3.60 | 7.2 | 3 | Shewanellaceae | 3.66 | 7.14 | 4 |
| Vibrio alginolyticus | 2.10 | 3.0 | 4 | Vibrionaceae | 21.14 | 20.13 | 12 |
| Vibrio fluvialis | Vibrionaceae | ||||||
| Vibrio cholerae | Vibrionaceae | ||||||
| Mycobacterium spp. | 0.03 | 0.1 | 1 | Mycobacteriaceae | 0.03 | 0.1 | 1 |
| Turtle ID | Date of Admit | Date of Necropsy | Histologic Diagnoses (Lung) | Culture Isolates from Tracheal Wash at Admit | Culture Isolates at Necropsy |
|---|---|---|---|---|---|
| 25 | 23 November 2015 | 14 December 2015 | Severe necrotizing and granulomatous pneumonia with intralesional fungal hyphae (with vascular invasion) | Serratia liquefaciens Shewanella sp. Vibrio sp. Non-enteric gram neg rod | Hafnia (Enterobacter) alvei Mucor sp. Paecilomyces sp. |
| 27 | 23 November 2015 | 2 December 2015 | Heterophilic bronchopneumonia with intralesional bacteria | NE | NE |
| 36 | 24 November 2015 | 26 Dececmber 2015 | Severe necrotizing, heterophilic, and granulomatous pneumonia with numerous intralesional fungi and bacteria | Vibrio sp. | Serratia marcesens Citrobacter braakii Providencia rettgeri |
| 247 | 20 December 2015 | 14 February 2016 | Severe, diffuse, chronic granulomatous pneumonia with intralesional fungal hyphae | NE | gram neg rods (rare) * Paecilomyces sp. * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McNally, K.L.; Bowen, J.L.; Brisson, J.O.; Kennedy, A.; Innis, C.J. Evaluation of the Respiratory Microbiome and the Use of Tracheal Lavage as a Diagnostic Tool in Kemp’s Ridley Sea Turtles (Lepidochelys kempii). Animals 2021, 11, 2927. https://doi.org/10.3390/ani11102927
McNally KL, Bowen JL, Brisson JO, Kennedy A, Innis CJ. Evaluation of the Respiratory Microbiome and the Use of Tracheal Lavage as a Diagnostic Tool in Kemp’s Ridley Sea Turtles (Lepidochelys kempii). Animals. 2021; 11(10):2927. https://doi.org/10.3390/ani11102927
Chicago/Turabian StyleMcNally, Kerry L., Jennifer L. Bowen, Jennifer O. Brisson, Adam Kennedy, and Charles J. Innis. 2021. "Evaluation of the Respiratory Microbiome and the Use of Tracheal Lavage as a Diagnostic Tool in Kemp’s Ridley Sea Turtles (Lepidochelys kempii)" Animals 11, no. 10: 2927. https://doi.org/10.3390/ani11102927
APA StyleMcNally, K. L., Bowen, J. L., Brisson, J. O., Kennedy, A., & Innis, C. J. (2021). Evaluation of the Respiratory Microbiome and the Use of Tracheal Lavage as a Diagnostic Tool in Kemp’s Ridley Sea Turtles (Lepidochelys kempii). Animals, 11(10), 2927. https://doi.org/10.3390/ani11102927