
Characterization, Stress Response and Functional Analyses of Giant River Prawn (Macrobrachium rosenbergii) Glucose-Regulated Protein 78 (Mr-grp78) under Temperature Stress and during Aeromonas hydrophila Infection



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. PCR with 5′ Rapid Amplification cDNA Ends (5′RACE)
2.3. Cloning and Characterization of the Full-Length cDNA Encoding the grp78 Gene of Giant River Prawn
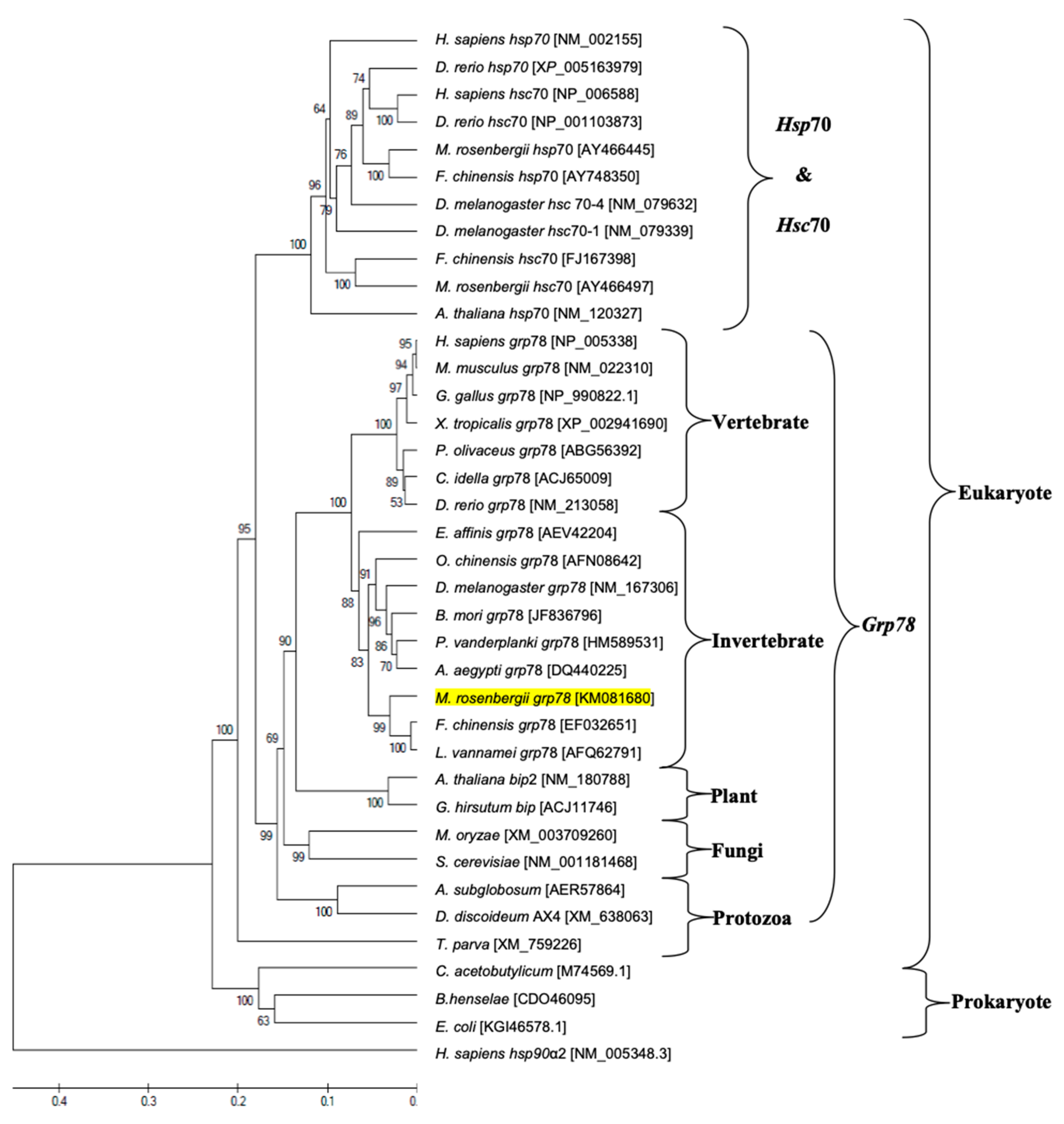
2.4. Phylogenetic Analysis
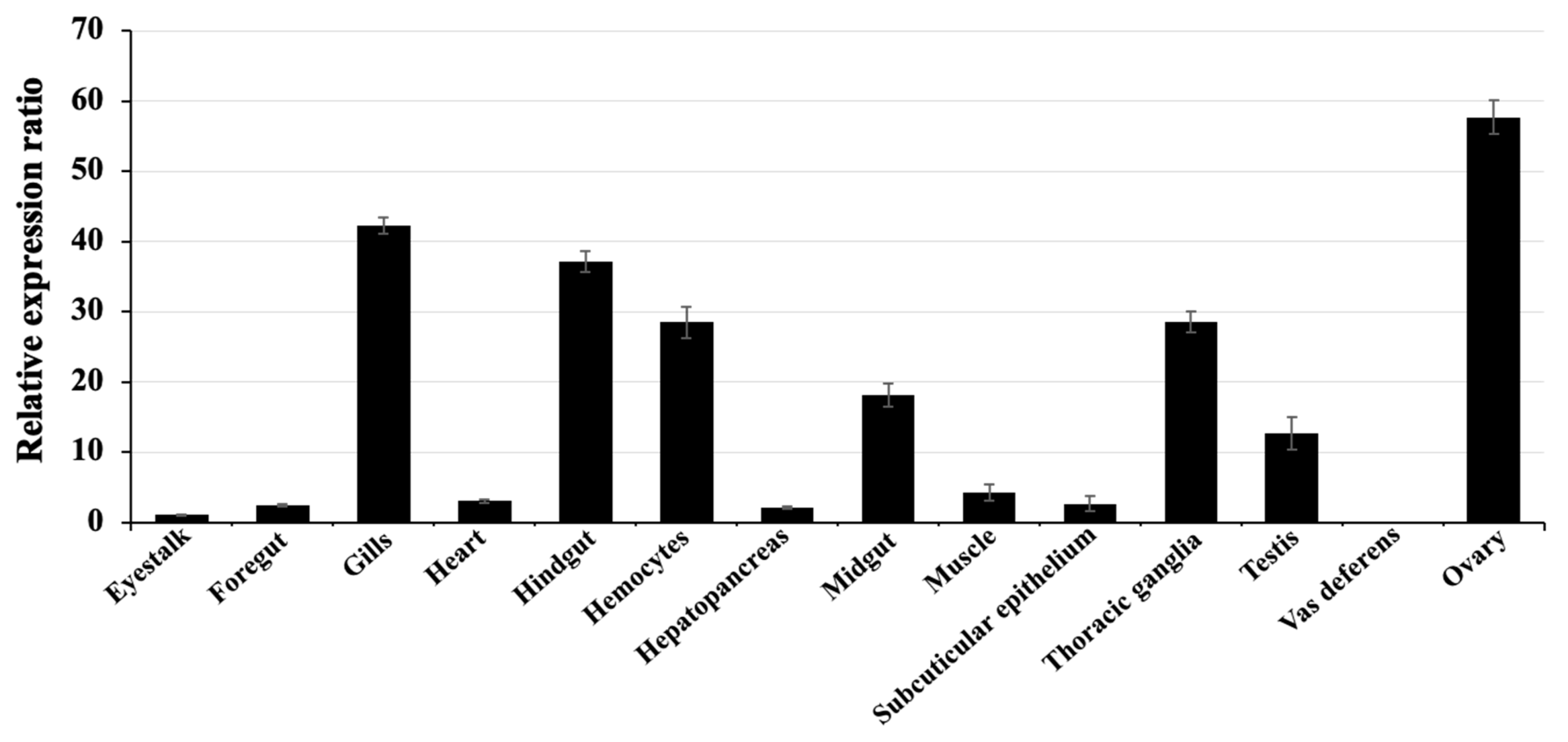
2.5. Determination of the Distribution of grp Genes in Various Tissues of Healthy Giant River Prawns Based on Quantitative Real-Time RT-PCR (qRT-PCR)
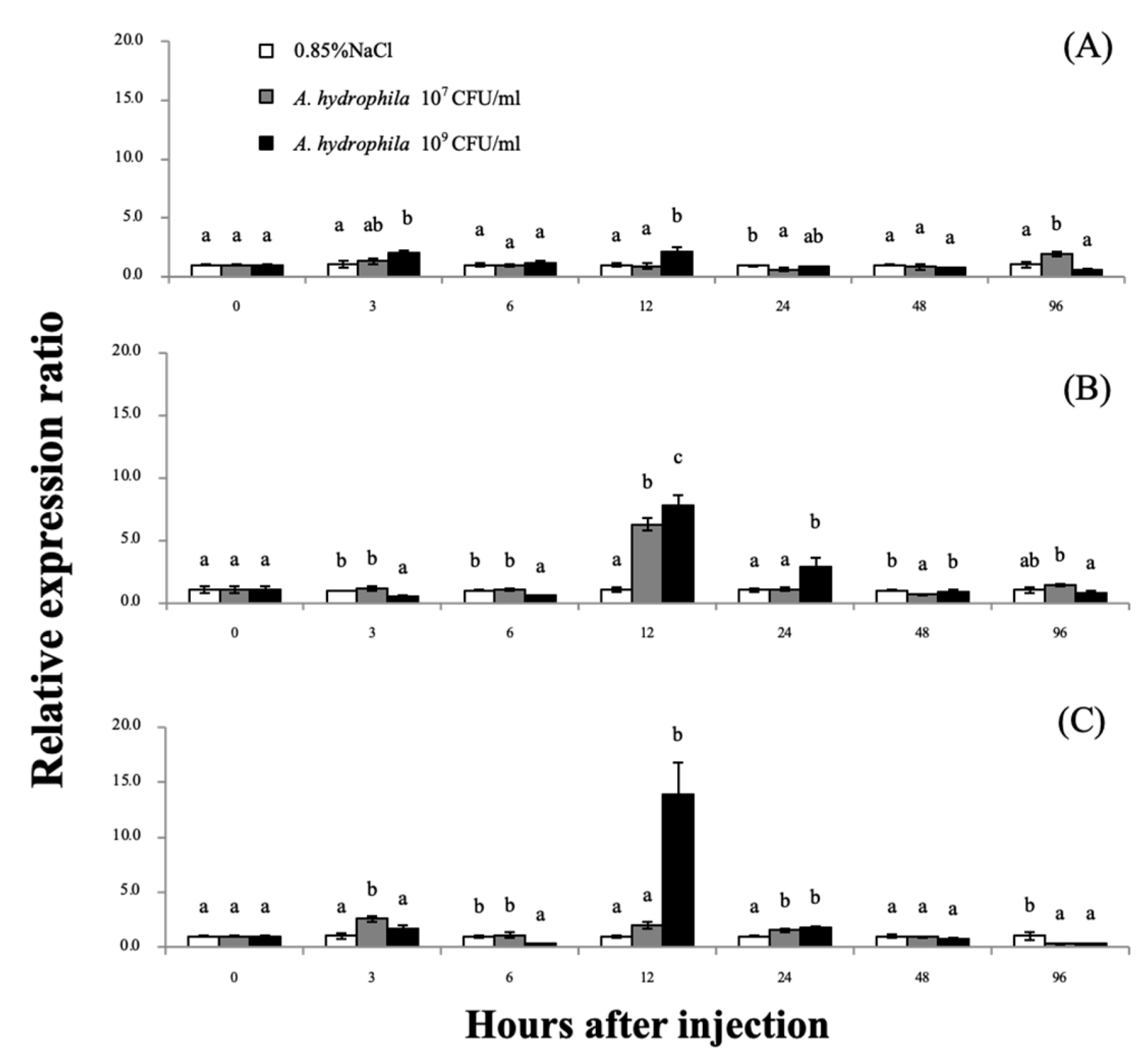
2.6. Analysis of the grp78 mRNA Expression Response in Giant River Prawns under Aeromonas hydrophila Stimulation
2.7. Response Analyses of grp78 mRNA in Giant River Prawns to Heat- and Cold-Shock Induction
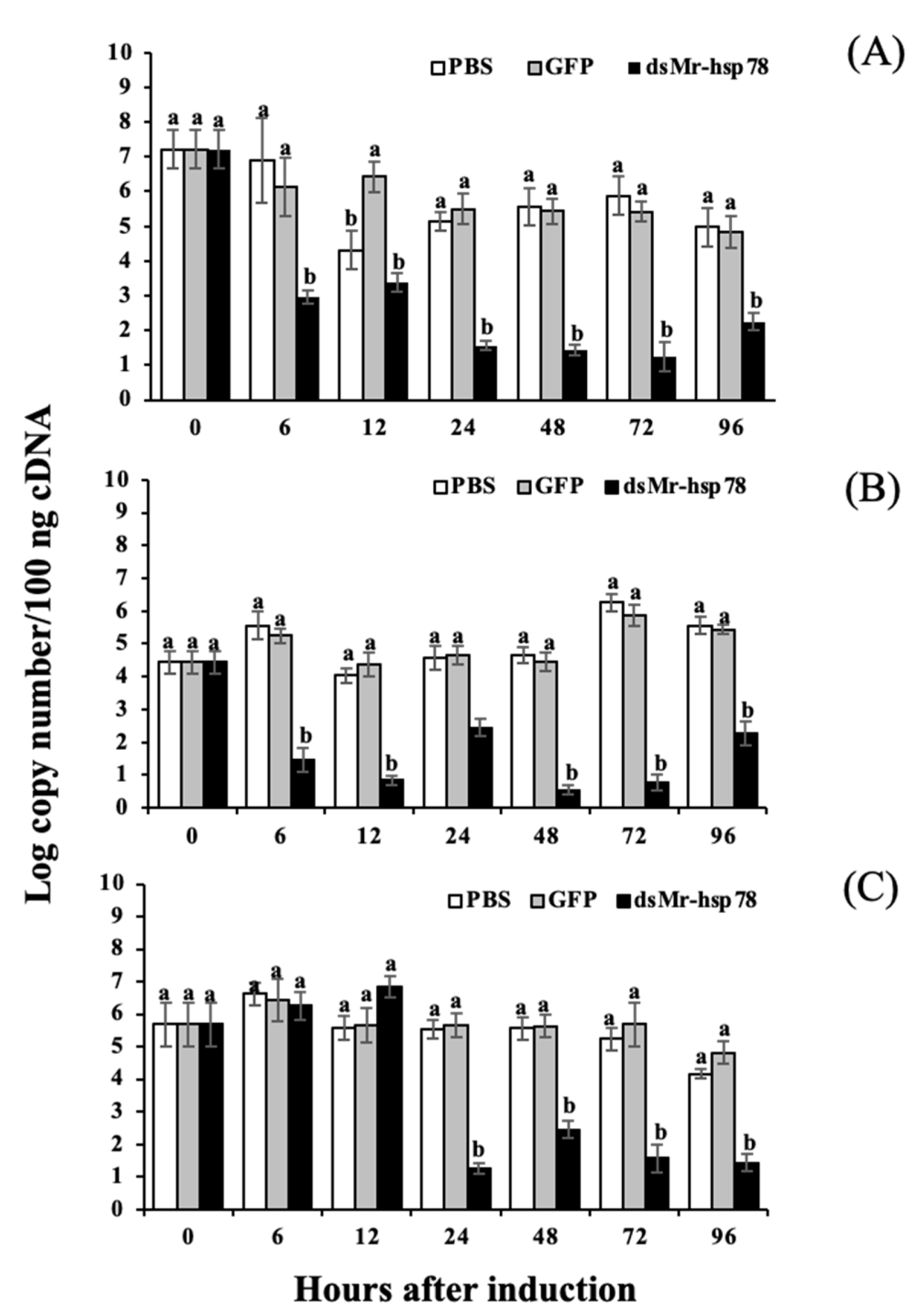
2.8. Gene Knockdown Analysis of the Mr-grp78 Gene
2.9. Effects of Mr-grp78 Gene Silencing under High Temperature Stress and during A. hydrophila Infection
2.10. Effects of Mr-grp78 Gene Silencing on A. hydrophila (AH) Infection
3. Results
3.1. Analysis of the Mr-grp78 cDNA of Giant River Prawn
3.2. Nucleotide and Amino Acid Sequence Analyses and Phylogenetic Tree Construction of Mr-grp78
3.3. mRNA Level Expression of the grp78 Gene in Various Tissues of Giant River Prawn by qRT-PCR Analysis
3.4. Expression Analyses of the Mr-grp78 mRNAs after A. hydrophila Induction
3.5. Expression Analyses of Mr-grp78 mRNA in Response to Heat and Cold Shocks
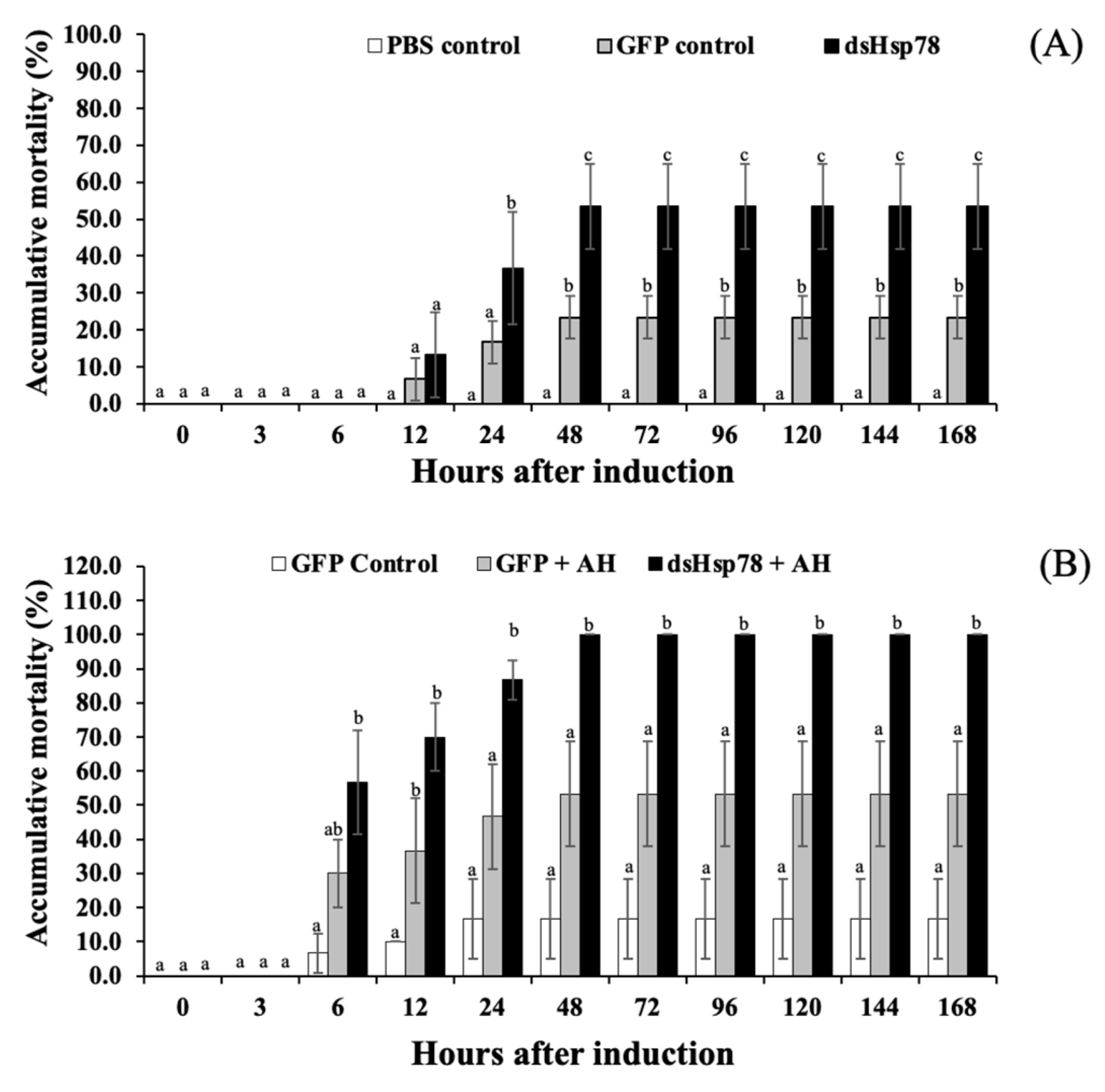
3.6. Silencing Analysis of Mr-grp78 mRNA under Heat Stress and A. hydrophila Infection
4. Discussion
4.1. Characterization of Mr-grp78
4.2. Expression Analysis of Mr-grp78
4.3. Expression Analysis of Mr-grp78 under A. hydrophila Induction
4.4. Analysis of Mr-grp78 Expression under Heat and Cold Temperature Conditions
4.5. Functional Analysis of Mr-grp78 under Heat and Cold Temperature Conditions by Gene Knockdown
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Rupik, W.; Jasik, K.; Bembenek, J.; Widlak, W. The expression patterns of heat shock genes and proteins and their role during vertebrate's development. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 159, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S.; Craig, E.A. The heat shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef] [PubMed]
- Moseley, P. Stress proteins and the immune response. Immunopharmacology 2000, 48, 299–302. [Google Scholar] [CrossRef]
- Schröder, M. Endoplasmic reticulum stress responses. Cell. Mol. Life Sci. 2008, 65, 862–894. [Google Scholar] [CrossRef]
- Giorgi, C.; de Stefani, D.; Bononi, A.; Rizzuto, R.; Pinton, P. Structural and functional link between the mitochondrial network and the endoplasmic reticulum. Int. J. Biochem. Cell Biol. 2009, 41, 1817–1827. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Hendershot, L.M. The unfolding tale of the unfolded protein response. Cell 2001, 107, 827–830. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, R.J.; Scheuner, D.; Schroder, M.; Shen, X.; Lee, K.; Liu, C.Y.; Arnold, S.M. The unfolded protein response in nutrient sensing and differentiation. Nat. Rev. Mol. Cell Biol. 2002, 3, 411–421. [Google Scholar] [CrossRef]
- Adams, C.J.; Kopp, M.C.; Larburu, N.; Nowak, P.R.; Ali, M.M.U. Structure and molecular mechanism of ER stress signaling by the unfolded protein response signal activator IRE1. Front. Mol. Biosci. 2019, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.C.; Wei, S.; Li, C.W.; Chang, Y.S.; Chao, C.C.; Lai, Y.K. Localization of GRP78 to mitochondria under the unfolded protein response. Biochem. J. 2006, 396, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Falahatpisheh, H.; Nanez, A.; Montoya-Durango, D.; Qian, Y.; Tiffany-Castiglioni, E.; Ramos, K.S. Activation profiles of HSPA5 during the glomerular mesangial cell stress response to chemical injury. Cell Stress Chaperones 2007, 12, 209–218. [Google Scholar] [CrossRef]
- Yu, Z.F.; Luo, H.; Fu, W.M.; Mattson, M.P. The endoplasmic reticulum stress-responsive protein GRP78 protects neurons against excitotoxicity and apoptosis: Suppression of oxidative stress and stabilization of calcium homeostasis. Exp. Neurol. 1999, 155, 302–314. [Google Scholar] [CrossRef]
- Misra, U.K.; Mario, G.G.; Gawdi, G.; Wang, F.; Pizzo, S.V. A novel receptor function for the heat shock protein Grp78: Silencing of Grp78 gene expression attenuates α2M-induced signaling. Cell. Signal. 2004, 16, 929–938. [Google Scholar] [CrossRef]
- Shields, A.M.; Panayi, G.S.; Corrigall, V.M. Resolution-associated molecular patterns (RAMP): RAM Parts defending immunological homeostasis. Clin. Exp. Immunol. 2011, 165, 292–300. [Google Scholar] [CrossRef]
- Morito, D.; Nagata, K. ER stress proteins in autoimmune and inflammatory diseases. Front. Immunol. 2012, 3, 48. [Google Scholar] [CrossRef] [Green Version]
- Moffitt, C.M.; Cajas-Cano, L. Blue growth: The 2014 FAO state of world fisheries and aquaculture. Fisheries 2014, 39, 552–553. [Google Scholar] [CrossRef]
- New, M.B. Freshwater prawn culture: A review. Aquaculture 1990, 88, 99–143. [Google Scholar] [CrossRef]
- Cheng, W.; Chen, J.C. Enterococcus-like infections in Macrobrachium rosenbergii are exacerbated by high pH and temperature but reduced by low salinity. Dis. Aquat. Org. 1998, 34, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habashy, M.; Hassan, M.M.S. Effects of temperature and salinity on growth and reproduction of the river prawn, Macrobrachium rosenbergii (Crustacea- Decapoda) in Egypt. Madlen, IJESE 2011, 1, 83–90. [Google Scholar]
- Kumaresan, V.; Palanisamy, R.; Pasupuleti, M.; Arockiaraj, J. Impacts of environmental and biological stressors on immune system of Macrobrachium rosenbergii. Rev. Aquac. 2016, 9, 1–25. [Google Scholar]
- Sung, H.H.; Hwang, S.F.; Tasi, F.M. Responses of giant river prawn (Macrobrachium rosenbergii) to challenge by two strains of Aeromonas spp. J. Invertebr. Pathol. 2000, 76, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Abdolnabi, S.; Ina-Salwany, M.Y.; Daud, H.M.; Mariana, N.S.; Abdelhadi, Y.M. Pathogenicity of Aeromonas hydrophila in giant river prawn; Macrobrachium rosenbergii, cultured in East Malaysia. Iran. J. Fish. Sci. 2014, 14, 232–245. [Google Scholar]
- Luan, W.; Li, F.; Zhang, J.; Wang, B.; Xiang, J. Cloning and expression of glucose regulated protein 78 (GRP78) in Fenneropenaeus chinensis. Mol. Biol. Rep. 2009, 36, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, P.; Zhao, C.; Qiu, L. The anti-stresses capability of GRP78 in Penaeus monodon: Evidence from in vitro and in vivo studies. Fish Shellfish Immunol. 2018, 72, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Zhao, L.; Pang, L.R.; Li, X.Y.; Weng, S.P.; He, J.G. Identification and characterization of Inositol-requiring enzyme-1 and X-box binding protein 1, two proteins involved in the unfolded protein response of Litopenaeus vannamei. Dev. Comp. Immunol. 2012, 38, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Toe, A.; Areechon, N.; Srisapoome, P. Molecular characterization and immunological response analysis of a novel transferrin-like, pacifastin heavy chain protein in giant freshwater prawn, Macrobrachium rosenbergii (De Man, 1879). Fish Shellfish Immunol. 2012, 33, 801–812. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Itoh, H.; Komatsuda, A.; Ohtani, H.; Wakui, H.; Imai, H.; Sawada, K.I.; Otaka, M.; Ogura, M.; Suzuki, A.; Hamada, F. Mammalian HSP60 is quickly sorted into the mitochondria under conditions of dehydration. Eur. J. Biochem. 2002, 269, 5931–5938. [Google Scholar] [CrossRef]
- Krenek, S.; Schlegel, M.; Berendonk, T.U. Convergent evolution of heat-inducibility during subfunctionalization of the Hsp70 gene family. BMC Evol. Biol. 2013, 13, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukau, B.; Horwich, A.L. The Hsp70 and Hsp60 chaperone machines. Cell 1998, 92, 351–366. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, W.J.; Zhu, X.J.; Karouna-Renier, N.K.; Rao, R.K. Molecular cloning and expression of two HSP70 genes in the prawn, Macrobrachium rosenbergii. Cell Stress Chaperones 2004, 9, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Munro, S.; Pelham, H.R. An hsp70-like protein in the ER: Identity with the 78 kD glucose-regulated protein and immunoglobulin heavy chain binding protein. Cell 1986, 46, 291–300. [Google Scholar] [CrossRef]
- Shim, J.K.; Kim, H.; Lee, K.Y. Molecular identification of glucose-regulated protein 78 (grp78) gene from the Indian meal moth, Plodia interpunctella, and its regulation by nutrient uptake. J. Asia Pac. Entomol. 2014, 17, 303–309. [Google Scholar] [CrossRef]
- Chaurasia, M.K.; Nizam, F.; Ravichandran, G.; Arasu, M.V.; Al-Dhabi, N.A.; Arshad, A.; Elumalai, P.; Arockiaraj, J. Molecular importance of prawn large heat shock proteins 60, 70 and 90. Fish Shellfish Immunol. 2016, 48, 228–238. [Google Scholar] [CrossRef]
- Dhahbi, J.M.; Mote, P.L.; Tillman, J.B.; Walford, R.L.; Spindler, S.R. Dietary energy tissue-specifically regulates endoplasmic reticulum chaperone gene expression in the liver of mice. J. Nutr. 1997, 127, 1758–1764. [Google Scholar] [CrossRef]
- Otero, J.H.; Beáta, L.; Linda, M. Hendershot life and death of a BiP substrate. Semin. Cell Dev. Biol. 2010, 21, 472–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, Y.Y.; MacRae, T.H. Heat shock proteins and disease control in aquatic organisms. J. Aquac. Res. Dev. 2012, S2, 006. [Google Scholar]
- Tsutsumi, S.; Neckers, L. Extracellular heat shock protein 90: A role for a molecular chaperone in cell motility and cancer metastasis. Cancer Sci. 2007, 98, 1536–1539. [Google Scholar] [CrossRef] [PubMed]
- Vega, V.L.; Rodriguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; de Maio, A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. J. Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, J.; Untergasser, G.; Zenzmaier, C.; Sarg, B.; Gastl, B.; Gunsilius, E.; Steurer, M. GRP-78 secreted by tumor cells blocks the antiangiogenic activity of bortezomib. Blood 2009, 114, 3960–3967. [Google Scholar] [CrossRef] [Green Version]
- Chalmin, F.; Ladoire, S.; Mignot, G.; Vincent, J.; Bruchard, M.; Remy-Martin, J.P.; Boireau, W.; Rouleau, A.; Simon, B.; Lanneau, D.; et al. Membrane-associated Hsp72 from tumor-derived exosomes mediates STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. J. Clin. Investig. 2010, 120, 457–471. [Google Scholar] [CrossRef]
- Langer, T.; Lu, C.; Echols, H.; Flanagan, J.; Hayer, M.K.; Hartl, F.U. Successive action of DnaK, DnaJ and GroEL along the pathway of chaperone mediated protein folding. Nature 1992, 356, 683–689. [Google Scholar] [CrossRef]
- Hsu, J.P.; Huang, C.; Liao, C.M.; Hsuan, S.L.; Hung, H.H.; Chien, M.S. Engulfed pathogen-induced apoptosis in haemocytes of giant river prawn, Macrobrachium rosenbergii. J. Fish Dis. 2005, 28, 729–735. [Google Scholar] [CrossRef]
- Quinones, Q.J.; de Ridder, G.G.; Pizzo, S.V. GRP78: A chaperone with diverse roles beyond the endoplasmic reticulum. Histol. Histopathol. 2008, 23, 1409–1416. [Google Scholar]
- Kim, Y.S.; Seo, H.W.; Jung, G. Reactive oxygen species promote heat shock protein 90-mediated HBV capsid assembly. Biochem. Biophys. Res. Commun. 2015, 457, 328–333. [Google Scholar] [CrossRef]
- Breckenridge, D.G.; Germain, M.; Mathai, J.P.; Nguyen, M.; Shore, G.C. Regulation of apoptosis by endoplasmic reticulum pathways. Oncogene 2003, 22, 8608–8618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brett, J.R. Implications and assessments of environmental stress. In Investigations of Fish-Power Problems; Larkin, P.A., MacMillan, H.R., Eds.; Lectures in Fisheries; University of British Columbia: Vancouver, BC, Canada, 1958; pp. 69–83. [Google Scholar]
- Hennig, O.L.; Andreatta, E.R. Effect of temperature in an intensive nursery system for Penaeus paulensis (Pérez Farfante, 1967). Aquaculture 1998, 164, 167–172. [Google Scholar] [CrossRef]
- Manush, S.M.; Pal, A.K.; Chatterjee, N.; Das, T.; Mukherjee, S.C. Thermal tolerance and oxygen consumption of Macrobrachium rosenbergii acclimated to three temperatures. J. Therm. Biol. 2004, 29, 15–19. [Google Scholar] [CrossRef]
- Snyder, G. Water Temperature Effects on Fish and Aquatic Life. 2016. Available online: http://sciencefairwater.com/physical-water-quality-parameters/water-temperature/water-temperature-effects-on-fish-and-aquatic-life/,2016 (accessed on 16 January 2021).
- Hall, D.M.; Xu, L.; Drake, V.J.; Oberley, L.W.; Oberley, T.D.; Moseley, P.L.; Kregel, K.C. Aging reduces adaptive capacity and stress protein expression in the liver after heat stress. J. Appl. Physiol. 2000, 89, 749–759. [Google Scholar] [CrossRef]
- Deane, E.E.; Woo, N.Y.S. Growth hormone increases hsc70/hsp70 expression and protects against apoptosis in whole blood preparations from silver sea bream. Ann. N.Y. Acad. Sci. 2005, 1040, 288–292. [Google Scholar] [CrossRef]
- Ojima, N.; Yamashita, M.; Watabe, S. Comparative expression analysis of two paralogous Hsp70s in rainbow trout cells exposed to heat stress. Biochim. Biophys. Acta 2005, 1681, 99–106. [Google Scholar] [CrossRef]
- Xuereb, B.; Forget-Leray, J.; Souissi, S.; Glippa, O.; Devreker, D.; Lesueur, T.; Marie, S.; Danger, J.M.; Boulangé-Lecomte, C. Molecular characterization and mRNA expression of grp78 and hsp90A in the estuarine copepod Eurytemora affinis. Cell Stress Chaperones 2012, 17, 457–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spees, J.L.; Chang, S.A.; Snyder, M.J.; Chang, E.S. Thermal acclimation and stress in the American lobster, Homarus americanus: Equivalent temperature shifts elicit unique gene expression patterns for molecular chaperones and polyubiquitin. Cell Stress Chaperones 2002, 7, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.S. Stressed-out lobsters: Crustacean hyperglycemic hormone and stress proteins. Integr. Comp. Biol. 2005, 45, 43–50. [Google Scholar] [CrossRef]
- Mote, P.L.; Tillman, J.B.; Spindler, S.R. Glucose regulation of GRP78 gene expression. Mech. Ageing Dev. 1998, 104, 149–158. [Google Scholar] [CrossRef]
- Kim, B.M.; Jeong, C.B.; Han, J.; Kim, I.C.; Rhee, J.S.; Lee, J.S. Role of crustacean hyperglycemic hormone (CHH) in the environmental stressor-exposed intertidal copepod Tigriopus japonicas. Comp. Biochem. Physiol. C 2013, 158, 131–141. [Google Scholar]
- Paschen, W.; Mengesdorf, T.; Althausen, S.; Hotop, S. Peroxidative stress selectively down-regulates the neuronal stress response activated under conditions of endoplasmic reticulum dysfunction. J. Neurochem. 2001, 76, 1916–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prangnell, D.I.; Fotedar, R. The growth and survival of western king prawns, Penaeus latisulcatus Kishinouye, in potassium-fortified inland saline water. Aquaculture 2006, 259, 234–242. [Google Scholar] [CrossRef]
- Wang, W.; Wang, A.; Liu, Y.; Xiu, J.; Liu, Z.; Sun, R. Effects of temperature on growth, adenosine phosphates, ATPase and cellular defense response of juvenile shrimp Macrobrachium nipponense. Aquaculture 2006, 256, 624–630. [Google Scholar] [CrossRef]
- Malev, O.; Rut, M.; Maguire, I.; Tambuk, A.; Ferrero, E.A.; Lorenzon, S.; Klobučar, G.I.V. Genotoxic physiological and immunological effects caused by temperature increase, air exposure or food deprivation in river crayfish Astacus leptodactylus. Comp. Biochem. Physiol. C 2010, 152, 433–443. [Google Scholar]
- Zhu, G.; Lee, A.S. Role of the unfolded protein response, GRP78 and GRP94 in organ homeostasis. J. Cell. Physiol. 2015, 230, 1413–1420. [Google Scholar] [CrossRef] [Green Version]
- Whangchai, N.; Ungsethaphand, T.; Chitmanat, C.; Mengumphan, K.; Uraiwan, S. Performance of giant river prawn (Macrobrachium rosenbergii de Man) reared in earthen ponds beneath plastic film shelters. Chiang Mai J. Sci. 2007, 34, 89–96. [Google Scholar]
- Qiu, J.; Wang, W.N.; Wang, L.; Liu, Y.F.; Wang, A.L. Oxidative stress, DNA damage and osmolality in the Pacific white shrimp, Litopenaeus vannamei exposed to acute low temperature stress. Comp. Biochem. Physiol. C 2011, 154, 36–41. [Google Scholar] [CrossRef]
- Weber, T.E.; Bosworth, B.G. Effects of 28-day exposure to cold temperature or feed restriction on growth, body composition, and expression of genes related to muscle growth and metabolism in channel catfish. Aquaculture 2005, 246, 483–492. [Google Scholar] [CrossRef]
- Wu, Y.P.; Chang, C.M.; Hung, C.Y.; Tsai, M.C.; Schuyler, S.C.; Wang, R.Y. Japanese encephalitis virus co-opts the ER-stress response protein GRP78 for viral infectivity. Virol. J. 2011, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, D.S.-C.; Lee, S.-B.; Chu, C.-S.; Chang, L.-H.; Chung, B.-C.; Juan., L.-J. Transcriptional activation of endoplasmic reticulum chaperone GRP78 by HCMV IE1-72 protein. Cell Res. 2011, 21, 642–653. [Google Scholar]
- Wei, D.; Li, N.L.; Zeng, Y.; Liu, B.; Kumthip, K.; Wang, T.T.; Huo, D.; Ingels, J.F.; Lu, L.; Shang, J.; et al. The molecular chaperone GRP78 contributes to Toll-like receptor 3-mediated innate immune response to hepatitis C virus in hepatocytes. J. Biol. Chem. 2016, 291, 12294–12309. [Google Scholar] [CrossRef] [Green Version]
- Nunes, F.M.F.; Aleixo, A.C.; Barchuk, A.R.; Bomtorin, A.D.; Grozinger, C.M.; Simões, Z.L.P. Non-target effects of green fluorescent protein (GFP)-derived double-stranded RNA (dsRNA-GFP) used in honey bee RNA interference (RNAi) assays. Insects 2013, 4, 90–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Primer Sequence (5’-3’) | Size | Purposes |
|---|---|---|---|---|
| β-actin | RTB-actin_F | TTCACCATCGGCATTGAGAGGTTC | 119 bp | Real-Time PCR |
| RTB-actin_R | CACGTCGCACTTCATGATGGAGTT | Real-Time PCR | ||
| Mr-grp78 | F_GFP_grp78 | TCTGCTGAAGATAAGGGCACTGGT | 524 bp | RT-RCR |
| R_GFP_grp78 | ACCTGCACCCTGGTATAACTTAGC | RT-RCR, Touchdown PCR in RACE | ||
| RTF_GFP_grp78 | TCTGCTGAAGATAAGGGCACTGGT | 110 bp | Real-Time PCR | |
| RTR_GFP_grp78 | TCAGCATCCTTGATCATCCTCTCG | Real-Time PCR | ||
| Mr-HSP78T7_F | GGATCCTAATACGACTCACTATAGGGGCGACAAGTCTGTCCAGCA | 272 bp | Gene silencing | |
| Mr-HSP78T7_R | GGATCCTAATACGACTCACTATAGGACAGTCATGCCCGCAATGGT | Gene silencing | ||
| Green fluorescence protein (GFP) | GFP_F | TAATACGACTCACTAAGGGAGACACATGAAGCAGCACGACCT | Gene silencing | |
| GFP_R | TAATACGACTCACTATAGGGAGAAGTTCACCTTGATGCCGTTC | Gene silencing | ||
| UPM | LongUPM primer | CTAATACGACTCACTATAGGGCAAGCAGTGGTAACAACGCAGAGT | Touchdown PCR in RACE | |
| ShortUPM primer | CTAATACGACTCACTATAGGGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srisapoome, P.; Ju-Ngam, T.; Wongpanya, R. Characterization, Stress Response and Functional Analyses of Giant River Prawn (Macrobrachium rosenbergii) Glucose-Regulated Protein 78 (Mr-grp78) under Temperature Stress and during Aeromonas hydrophila Infection. Animals 2021, 11, 3004. https://doi.org/10.3390/ani11103004
Srisapoome P, Ju-Ngam T, Wongpanya R. Characterization, Stress Response and Functional Analyses of Giant River Prawn (Macrobrachium rosenbergii) Glucose-Regulated Protein 78 (Mr-grp78) under Temperature Stress and during Aeromonas hydrophila Infection. Animals. 2021; 11(10):3004. https://doi.org/10.3390/ani11103004
Chicago/Turabian StyleSrisapoome, Prapansak, Tanya Ju-Ngam, and Ratree Wongpanya. 2021. "Characterization, Stress Response and Functional Analyses of Giant River Prawn (Macrobrachium rosenbergii) Glucose-Regulated Protein 78 (Mr-grp78) under Temperature Stress and during Aeromonas hydrophila Infection" Animals 11, no. 10: 3004. https://doi.org/10.3390/ani11103004
APA StyleSrisapoome, P., Ju-Ngam, T., & Wongpanya, R. (2021). Characterization, Stress Response and Functional Analyses of Giant River Prawn (Macrobrachium rosenbergii) Glucose-Regulated Protein 78 (Mr-grp78) under Temperature Stress and during Aeromonas hydrophila Infection. Animals, 11(10), 3004. https://doi.org/10.3390/ani11103004