
Modified Nano-Montmorillonite and Monensin Modulate In Vitro Ruminal Fermentation, Nutrient Degradability, and Methanogenesis Differently

 ,
,  ,
,  ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of MNM Products
2.2. Physicochemical Properties of NM, MNMsds, and MNMCETAB
2.3. In Vitro Gas Production (GP)
2.3.1. Basal Feed Substrate and Experimental Design
2.3.2. Procedure of GP
2.3.3. In Vitro Ruminal Batch Culture Fermentation, Protozoal Count, and Nutrient Degradability
2.4. Statistical Analysis
3. Results
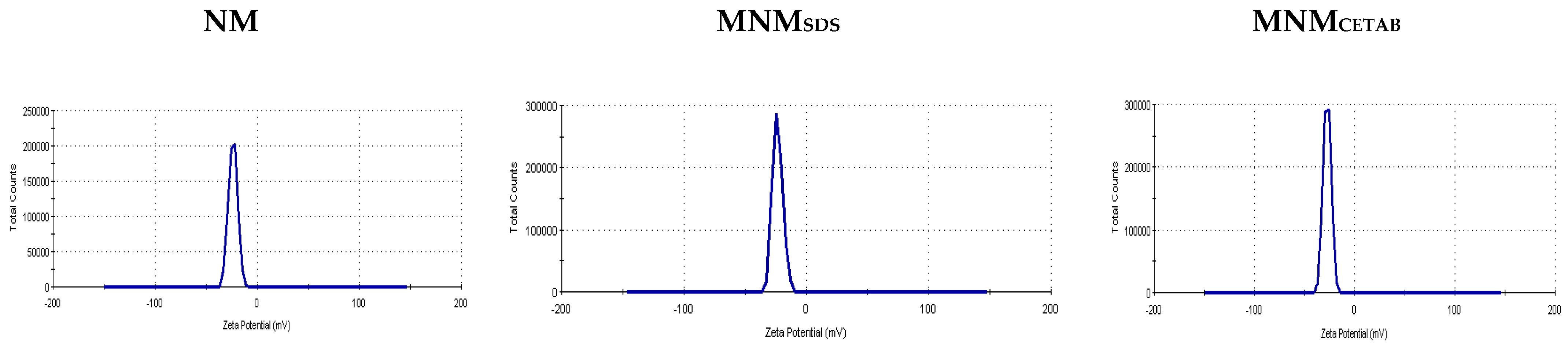
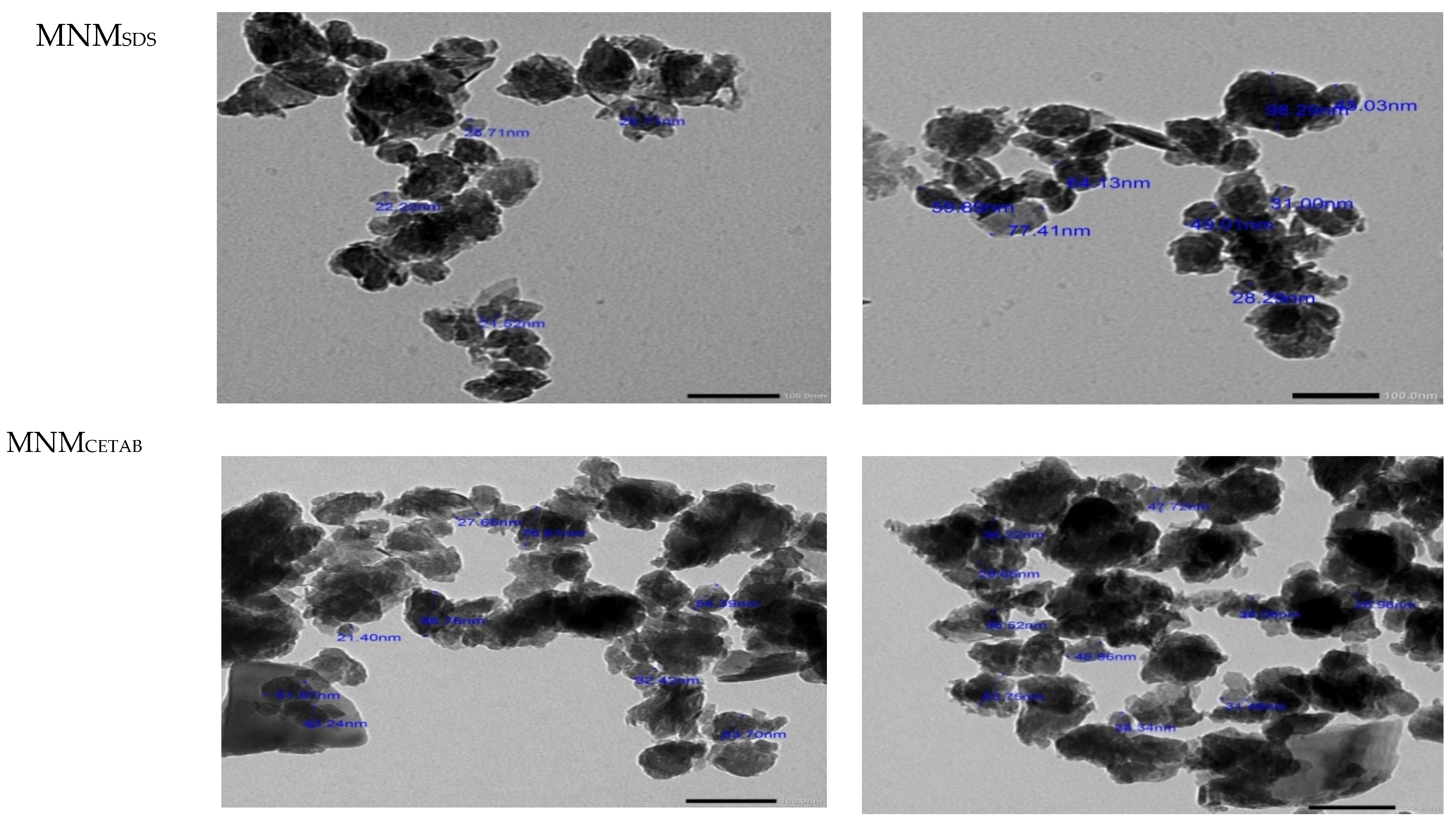
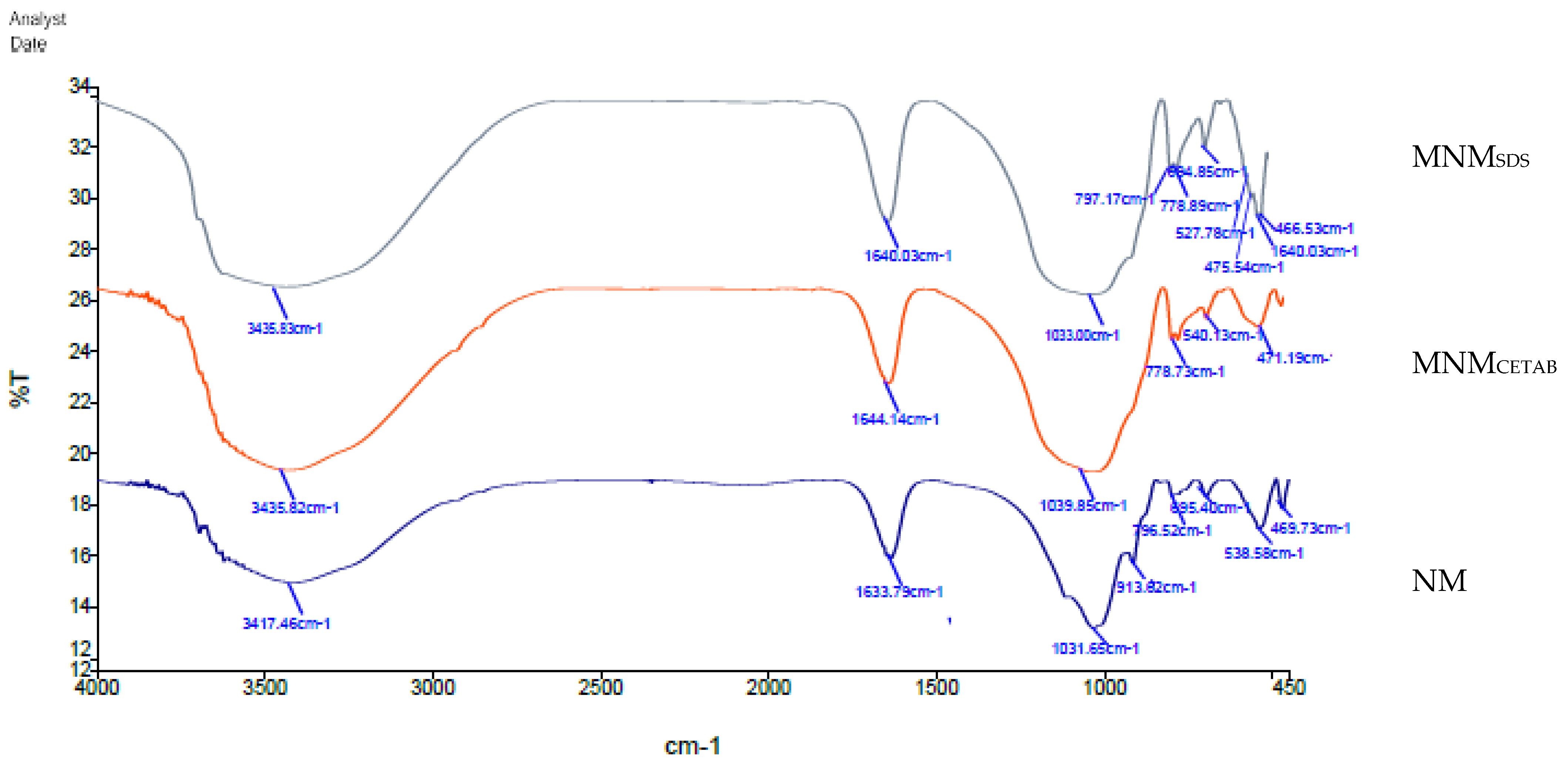
3.1. Physicochemical of NM, MNMsds, and MNMCETAB
3.2. Effect of MNM on In Vitro Ruminal Batch Culture GP, CH4, Nutrient Degradability, and Partitioning Factor
3.3. Effect of MNM Supplementation on In Vitro Ruminal Batch Culture pH, NH3-N and SCFAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Morsy, A.S.; Soltan, Y.A.; El-Zaiat, H.M.; Alencar, S.M.; Abdalla, A.L. Bee propolis extract as a phytogenic feed additive to enhance diet digestibility, rumen microbial biosynthesis, mitigating methane formation and health status of late pregnant ewes. Anim. Feed Sci. Technol. 2021, 273, 114834. [Google Scholar] [CrossRef]
- Soltan, Y.A.; Hashem, N.M.; Morsy, A.S.; El-Azrak, K.M.; Nour El-Din, A.; Sallam, S.M. Comparative effects of Moringa oleifera root bark and monensin supplementations on ruminal fermentation, nutrient digestibility and growth performance of growing lambs. Anim. Feed Sci. Technol. 2018, 235, 189–201. [Google Scholar] [CrossRef]
- Magaña, S.M.; Quintana, P.; Aguilar, D.H.; Toledo, J.A.; Ángeles-Chávez, C.; Cortés, M.A.; León, L.; Freile-Pelegrín, Y.; López, T.; Torres Sánchez, R.M. Antibacterial activity of montmorillonites modified with silver. J. Mol. Catal. A Chem. 2008, 281, 192–199. [Google Scholar] [CrossRef]
- Tate, K.; Yuan, G.; Theng, B.; Churchman, G.; Singh, J.; Berben, P. Can geophagy mitigate enteric methane emissions from cattle? J. Prelim. Res. 2015, 2, 1–8. [Google Scholar]
- Maryan, A.S.; Montazer, M. Natural and organo-montmorillonite as antibacterial nanoclays for cotton garment. J. Ind. Eng. Chem. 2015, 22, 164–170. [Google Scholar] [CrossRef]
- Maki, C.R.; Haney, S.; Wang, M.; Ward, S.H.; Bailey, R.H. Calcium montmorillonite clay for the reduction of aflatoxin residues in milk and dairy products. Dairy Vet. Sci. J. 2017, 2, 555587. [Google Scholar]
- Banat, F.; Al-Asheh, S.; Abu-Aitah, L. Examination of the effectiveness of Physical and chemical activation of natural bentonite for the removal of heavy metal ions from aqueous solutions. Adsorpt. Sci. Technol. 2002, 20, 151–167. [Google Scholar] [CrossRef]
- El-Nile, A.; Elazab, M.; El-Zaiat, H.; El-Azrak, K.E.; Elkomy, A.; Sallam, S.; Soltan, Y. In vitro and in vivo assessment of dietary supplementation of both natural or nano-zeolite in goat diets: Effects on ruminal fermentation and nutrients digestibility. Animals 2021, 11, 2215. [Google Scholar] [CrossRef]
- Bujdáková, H.; Bujdáková, V.; Májeková-Koščová, H.; Gaálová, B.; Bizovskác, V.; Boháčc, P.; Bujdák, J. Antimicrobial activity of organoclays based on quaternary alkylammonium and alkylphosphonium surfactants and montmorillonite. J. Appl. Clay Sci. 2018, 158, 21–28. [Google Scholar] [CrossRef]
- Rhoades, J.D. Cation exchange capacity. In Methods of Soil Analysis. Part 2: Chemical and Microbiological Methods; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar] [CrossRef] [Green Version]
- NRC (National Research Council). Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelid; The National Academy of Sciences: Washington, DC, USA, 2007. [Google Scholar]
- Association of Official Agricultural Chemists. Official Methods of Analysis, 20th ed.; Association of Official Agricultural Chemists: Arlington, VA, USA, 2006. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Bueno, I.C.S.; Cabral Filho, S.L.S.; Gobbo, S.P.; Louvandinin, H.; Vitti, D.M.S.S.; Abdallaa, A.L. Influence of inoculum source in a gas production method. Anim. Feed Sci. Technol. 2005, 123, 95–105. [Google Scholar] [CrossRef]
- Salama, H.A.S.; El-Zaiat, H.M.; Sallam, S.M.A.; Soltan, Y.A. Agronomic and qualitative characterization of multi-cut berseem clover (Trifolium alexandrinum L.) cultivars. J. Sci. Food Agric. 2020, 100, 3857–3865. [Google Scholar] [CrossRef] [PubMed]
- Sabry, M.; Nasser, M.E.A.; Kamel, H.E.M.; Abaza, M.A.; Soltan, Y.S. Effect of replacing corn grains with date palm kernels on ruminal fermentation, feed degradability, and methane production under different initial in vitro pH conditions. Anim. Biotechnol. 2021, in press. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Conra, H.R. Origin of plasma fatty acids in lactating cows fed high grain or high fat diets. J. Dairy Sci. 1971, 54, 1025–1033. [Google Scholar] [CrossRef]
- Dehority, B.A.; Damron, W.S.; McLaren, J.B. Occurrence of the rumen ciliate Oligoiso trichabubali in domestic cattle (Bostaurus). Appl. Environ. Microbiol. 1983, 45, 1394–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blümmel, M.; Steingab, H.; Becker, K. The relationship between in vitro gas production, in vitro microbial biomass yield and15N incorporations for the prediction of voluntary feed intake of roughages. Br. J. Nutr. 1997, 77, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Costa, E.C.B.; Araújo, G.G.L.; Oliveira, J.S.; Santos, E.M.; Henriques, L.T.; Perazzo, A.F.; Zanine, A.M.; Pereira, G.A.; Pinho, R.M.A. Effect of salt concentrations on in vitro rumen fermentation of cellulose, starch, and protein. South Afr. J. Anim. Sci. 2019, 49, 1139–1147. [Google Scholar] [CrossRef]
- Xue, W.; He, H.; Zhu, J.; Yuan, P. FTIR investigation of CTAB-Al-montmorillonite complexes. Spectrochim. Acta Part A 2007, 67, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Chouikhi, N.; Cecilia, J.A.; Vilarrasa-García, E.; Besghaier, S.; Chlendi, M.; Duro, F.I.F.; Castellon, E.R.; Bagane, M. CO2 adsorption of materials synthesized from clay minerals: A Review. Minerals 2019, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.B.; Strobel, H.J. Effect of ionophores on ruminal fermentation. Mini review. Appl. Environ. Microbiol. 1989, 55, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltan, Y.A.; Abdalla, A.L.; Silva, L.R.F.; Natel, A.S.; Morsy, A.S.; Louvandini, H. Response of different tropical pasture grass species to treatment with fibrolytic enzymes in terms of in vitro ruminal nutrient degradation and methanogenesis. Anim. Nutr. Feed Technol. 2013, 13, 551–568. [Google Scholar]
- Ruhee, R.T.; Roberts, L.A.; Ma, S.; Suzuki, K. Organosulfur compounds: A review of their anti-inflammatory effects in human health. Front. Nutr. 2020, 7, 64. [Google Scholar] [CrossRef]
- Hong, S.I.; Rhim, J.W. Antimicrobial activity of organically modified nano-clays. J. Nanosci. Nanotechnol. 2008, 8, 5818–5824. [Google Scholar] [CrossRef]
- Soltan, Y.; Adibe Filho, A.A.; Abdalla, A.; Berenchtein, B.; Schiavinatto, P.; Costa, C. Replacing maize with low tannin sorghum grains: Lamb growth performance, microbial protein synthesis and enteric methane production. Anim. Prod. Sci. 2021, 61, 1348–1355. [Google Scholar] [CrossRef]
- Ramos, S.C.; Jeong, C.D.; Mamuad, L.L.; Kim, S.H.; Son, A.R.; Miguel, M.A.; Islam, M.; Cho, Y.I.; Lee, S.S. Enhanced ruminal fermentation parameters and altered rumen bacterial community composition by formulated rumen buffer agents fed to dairy cows with a high-concentrate diet. Agriculture 2021, 11, 554. [Google Scholar] [CrossRef]
- Apajalahti, J.; Vienola, K.; Raatikainen, K.; Holder, V.; Moran, C.A. Conversion of branched-chain amino acids to corresponding isoacids—An in vitro tool for estimating ruminal protein degradability. Front. Vet. Sci. 2019, 6, 311. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, C.; Huang, Y.X.; Dong, K.H.; Yang, W.Z.; Zhangand, S.L.; Wang, H. Effects of isovalerate on ruminal fermentation, urinary excretion of purine derivatives and digestibility in steers. J. Anim. Physiol. Anim. Nutr. 2009, 93, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Soltan, Y.A.; Morsy, A.S.; Hashem, N.M.; Sallam, S.M. Boswellia sacra resin as a phytogenic feed supplement to enhance ruminal fermentation, milk yield, and metabolic energy status of early lactating goats. Anim. Feed Sci. Technol. 2021, 277, 114963. [Google Scholar] [CrossRef]
- Morsy, A.S.; Soltan, Y.A.; Sallam, S.M.A.; Kreuzer, M.; Alencar, S.M.; Abdalla, A.L. Comparison of the in vitro efficiency of supplementary bee propolis extracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation of methane. Anim. Feed Sci. Technol. 2015, 199, 51–60. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Item | Experimental Feed Substrate |
|---|---|
| (g/kg DM) | |
| Ingredients | |
| Trifolium alexandrinum clover | 500 |
| Ground maize | 233 |
| Wheat bran | 157 |
| Soybean meal | 90 |
| Calcium carbonate | 10 |
| Sodium chloride | 5 |
| Vitamins and minerals mixture 1 | 5 |
| Chemical composition | |
| Organic matter | 903 |
| Crude protein | 143 |
| Neutral detergent fiber | 395 |
| Acid detergent fiber | 194 |
| Acid detergent lignin | 49.9 |
| Ether extract | 32.1 |
| Items | NM | MNM | |
|---|---|---|---|
| MNMSDS | MNMCETAB | ||
| pH | 8.048 | 8.054 | 7.900 |
| Electrical conductivity (ppm) | 1408 | 960 | 230 |
| Cation exchange capacity (meq/100 g) | 77.5 | 117 | 81.9 |
| Zeta potential (mV) | −23.3 | −23.8 | −27.4 |
| Average particle size (nm) | 741.6 | 59.8 | 45.7 |
| Item | Treatments (T) | SEM | p Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Monensin | NM | MNM | T | Contrast 1 | Contrast 2 | |||||||
| MNMSDS | MNMCETAB | ||||||||||||
| Low | High | Low | High | Linear | Quadratic | Linear | Quadratic | ||||||
| Gas production (mL/g DM) | 113 ab | 99.2 c | 117 a | 109 abc | 102 bc | 102 bc | 97.7 c | 2.73 | <0.01 | <0.01 | 0.56 | <0.01 | 0.30 |
| Methane production | |||||||||||||
| (mL/g IOM) | 27.7 a | 19.5 bc | 25.9 ab | 20.1 bc | 17.3 cd | 20.2 bc | 11.9 d | 3.34 | <0.01 | <0.01 | 0.02 | <0.01 | 0.69 |
| (mL/g TDOM) | 43.8 a | 31.4 bc | 41.2 ab | 31.7 bc | 26.8 cd | 30.1 c | 17.4 d | 5.85 | <0.01 | <0.01 | 0.04 | <0.01 | 0.84 |
| Nutrient degradability | |||||||||||||
| TDOM | 632 ab | 614 b | 628 ab | 646 ab | 645 ab | 672 a | 652 a | 12.1 | 0.08 | 0.28 | 0.49 | 0.10 | 0.01 |
| TDNDF | 183 bcd | 143 d | 153 cd | 213 abc | 211 abc | 271 a | 227 ab | 20.74 | <0.01 | 0.29 | 0.49 | 0.10 | 0.01 |
| Partitioning factor | 2.80 cd | 3.13 abc | 2.69 d | 2.97 bcd | 3.17 ab | 3.33 a | 3.35 a | 0.07 | <0.01 | <0.01 | 0.84 | <0.01 | 0.01 |
| Item | Treatments (T) | SEM | p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Monensin | NM | MNM | T | Contrast 1 | Contrast 2 | |||||||
| MNMSDS | MNMCETAB | Linear | Quadratic | Linear | Quadratic | ||||||||
| Low | High | Low | High | ||||||||||
| pH | 5.59 b | 5.62 b | 5.63 b | 5.65 ab | 5.68 ab | 5.67 ab | 5.77 a | 0.025 | <0.01 | 0.13 | 0.51 | 0.13 | 0.01 |
| NH3-N (mg/100 mL) | 22.1 a | 16.3 b | 16.6 b | 17.5 ab | 14.5 b | 17.1 ab | 14.4 b | 1.02 | <0.01 | <0.01 | 0.69 | <0.01 | <0.01 |
| Protozoa (105/mL) | 7.02 ab | 6.00 b | 8.05 ab | 8.40 ab | 9.25 a | 9.00 a | 9.31 a | 0.597 | <0.01 | 0.03 | 0.74 | 0.02 | 0.28 |
| SCFAs | |||||||||||||
| Total (mM) | 104 | 105 | 98.6 | 107 | 106 | 102 | 103 | 2.5 | 0.51 | 0.62 | 0.51 | 0.73 | 0.41 |
| Acetate, % of total | 56.7 b | 56.4 b | 56.2 b | 61.1 a | 61.1 a | 60.5 a | 60.1 a | 0.58 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Propionate, % of total | 17.7 cd | 24.1 a | 18.2 bcd | 18.2 bcd | 18.0 bcd | 19.0 b | 18.9 b | 0.21 | <0.01 | 0.08 | <0.01 | <0.01 | <0.01 |
| Butyrate, % of total | 17.9 a | 14.0 b | 19.3 a | 13.9 b | 14.1 b | 13.1 b | 13.4 b | 0.38 | <0.01 | <0.01 | 0.03 | <0.01 | 0.01 |
| Isobutyrate, % of total | 1.77 ab | 1.33 b | 1.46 b | 1.43 b | 1.42 b | 2.47 ab | 2.58 a | 0.19 | 0.015 | 0.28 | 0.56 | 0.03 | 0.31 |
| Valerate, % of total | 1.56 ab | 1.08 c | 1.57 a | 1.59 a | 1.72 a | 1.24 c | 1.25 bc | 0.04 | <0.01 | 0.07 | 0.47 | <0.01 | <0.01 |
| Isovalerate, % of total | 4.17 a | 2.93 d | 3.83 b | 3.55 c | 3.69 bc | 3.52 c | 3.71 bc | 0.09 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| C2/C3 | 3.19 a | 2.33 b | 3.20 a | 3.32 a | 3.32 a | 3.18 a | 3.18 a | 0.02 | <0.01 | 0.01 | 0.13 | 0.88 | 0.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soltan, Y.; Morsy, A.; Hashem, N.; Elazab, M.; Sultan, M.; Marey, H.; Lail, G.A.E.; El-Desoky, N.; Hosny, N.; Mahdy, A.; et al. Modified Nano-Montmorillonite and Monensin Modulate In Vitro Ruminal Fermentation, Nutrient Degradability, and Methanogenesis Differently. Animals 2021, 11, 3005. https://doi.org/10.3390/ani11103005
Soltan Y, Morsy A, Hashem N, Elazab M, Sultan M, Marey H, Lail GAE, El-Desoky N, Hosny N, Mahdy A, et al. Modified Nano-Montmorillonite and Monensin Modulate In Vitro Ruminal Fermentation, Nutrient Degradability, and Methanogenesis Differently. Animals. 2021; 11(10):3005. https://doi.org/10.3390/ani11103005
Chicago/Turabian StyleSoltan, Yosra, Amr Morsy, Nesrein Hashem, Mahmoud Elazab, Mohamed Sultan, Haneen Marey, Gomaa Abo El Lail, Nagwa El-Desoky, Nourhan Hosny, Ahmed Mahdy, and et al. 2021. "Modified Nano-Montmorillonite and Monensin Modulate In Vitro Ruminal Fermentation, Nutrient Degradability, and Methanogenesis Differently" Animals 11, no. 10: 3005. https://doi.org/10.3390/ani11103005
APA StyleSoltan, Y., Morsy, A., Hashem, N., Elazab, M., Sultan, M., Marey, H., Lail, G. A. E., El-Desoky, N., Hosny, N., Mahdy, A., Hafez, E., & Sallam, S. (2021). Modified Nano-Montmorillonite and Monensin Modulate In Vitro Ruminal Fermentation, Nutrient Degradability, and Methanogenesis Differently. Animals, 11(10), 3005. https://doi.org/10.3390/ani11103005