
Welfare in Nile Tilapia Production: Dorsal Fin Erection as a Visual Indicator for Insensibility

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Rearing Conditions
2.2. Experimental Design and Procedures
2.3. Statistical Analysis
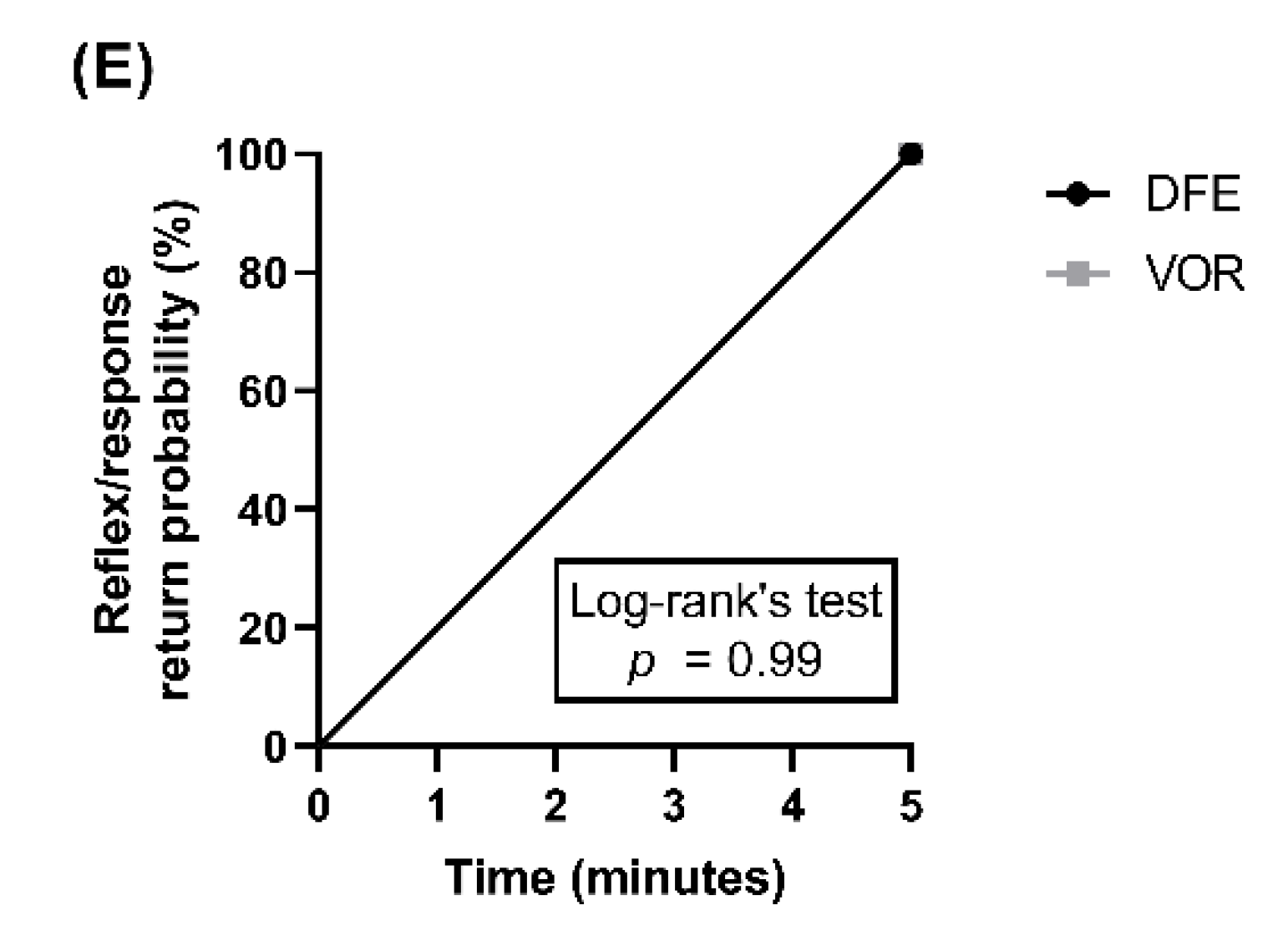
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Fish Not Submitted to Any Stunning Protocol-DFE and VOR Presence Control
References
- FAO. The State of Food and Agriculture 2020. Overcoming Water Challenges in Agriculture; FAO: Rome, Italy, 2020; p. 210. [Google Scholar] [CrossRef]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- European Food Safety (EFSA). General approach to fish welfare and to the concept of sentience in fish. EFSA J. 2009, 954, 1–27. [CrossRef]
- Frewer, L.J.; Kole, A.; Van de Kroon, S.M.A.; De Lauwere, C. Consumer Attitudes Towards the Development of Animal-Friendly Husbandry Systems. J. Agric. Environ. Ethics 2005, 18, 345–367. [Google Scholar] [CrossRef]
- Brijs, J.; Sandblom, E.; Axelsson, M.; Sundell, K.; Sundh, H.; Huyben, D.; Broström, R.; Kiessling, A.; Berg, C.; Gräns, A. The final countdown: Continuous physiological welfare evaluation of farmed fish during common aquaculture practices before and during harvest. Aquaculture 2018, 495, 903–911. [Google Scholar] [CrossRef]
- Saraiva, J.L.; Arechavala-Lopez, P. Welfare of fish—No longer the elephant in the room. Fishes 2019, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Clark, B.; Stewart, G.B.; Panzone, L.A.; Kyriazakis, I.; Frewer, L.J. Citizens, consumers and farm animal welfare: A meta-analysis of willingness-to-pay studies. Food Policy 2017, 68, 112–127. [Google Scholar] [CrossRef] [Green Version]
- Sneddon, L.U.; Lopez-Luna, J.; Wolfenden, D.C.C.; Leach, M.C.; Valentim, A.M.; Steenbergen, P.J.; Bardine, N.; Currie, A.D.; Broom, D.M.; Brown, C. Fish sentience denial: Muddying the waters. Anim. Sentience 2018, 3, 1. [Google Scholar] [CrossRef]
- Brown, C. Fish intelligence, sentience and ethics. Anim. Cogn. 2014, 18, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Branson, E.J. Fish Welfare, 1st ed.; Blackwell Publishing: Oxford, UK, 2008; p. 312. [Google Scholar]
- Pedrazzani, A.S.; Quintiliano, M.H.; Bolfe, F.; Sans, E.C.D.O.; Molento, C.F.M. Tilapia on-farm welfare assessment protocol for semi-intensive production systems. Front. Vet. Sci. 2020, 7, 991. [Google Scholar] [CrossRef] [PubMed]
- OIE. Welfare aspects of stunning and killing of farmed fish for human consumption. In OIE-Aquatic Animal Health Code, 22th ed.; OIE: Paris, France, 2019; Chapter 7; pp. 1–4. [Google Scholar]
- European Food Safety Authority (EFSA). Welfare aspects of the main systems of stunning and killing the main commercial species of animals. EFSA J. 2004, 45, 1–29. [Google Scholar]
- IBF; VetEffecT; Wageningen University; SANTE. Welfare of Farmed Fish: Common Practices during Transport and at Slaughter; European Commission: Luxembourg, 2017; Available online: http://publications.europa.eu/resource/cellar/facddd32-cda6-11e7-a5d5-01aa75ed71a1.0001.01/DOC_1 (accessed on 18 September 2021). [CrossRef]
- Oliveira Filho, P.R.C.; Oliveira, C.A.F.; Sobral, P.J.A.; Balieiro, J.C.C.; Natori, M.M.; Viegas, E.M.M. How stunning methods affect the quality of Nile tilapia meat. CYTA J. Food 2015, 13, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Poli, B.M.; Parisi, G.; Scappini, F.; Zampacavallo, G. Fish welfare and quality as affected by pre-slaughter and slaughter management. Aquac. Int. 2005, 13, 29–49. [Google Scholar] [CrossRef]
- Acerete, L.; Reig, L.; Alvarez, D.; Flos, R.; Tort, L. Comparison of two stunning/slaughtering methods on stress response and quality indicators of European sea bass (Dicentrarchus labrax). Aquaculture 2009, 287, 139–144. [Google Scholar] [CrossRef]
- Kestin, S.C.; van de Vis, J.W.; Robb, D.H.F. Protocol for assessing brain function in fish and the effectiveness of methods used to stun and kill them. Vet. Rec. 2002, 150, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Brijs, J.; Sundell, E.; Hjelmstedt, P.; Berg, C.; Senčić, I.; Sandblom, E.; Axelsson, M.; Lines, J.; Bouwsema, J.; Ellis, M.; et al. Humane slaughter of African sharptooth catfish (Clarias gariepinus): Effects of various stunning methods on brain function. Aquaculture 2021, 531, 735887. [Google Scholar] [CrossRef]
- Gregory, N.G.; Wotton, S.B. Studies on the central nervous system: Visually evoked cortical responses in sheep. Res. Vet. Sci. 1983, 34, 315–319. [Google Scholar] [CrossRef]
- Davis, M.W. Fish stress and mortality can be predicted using reflex impairment. Fish Fish. 2010, 11, 1–11. [Google Scholar] [CrossRef]
- Colgan, P. The Motivational Basis of Fish Behaviour. In The Behaviour of Teleost Fishes, 1st ed.; Pitcher, T., Ed.; Chapman and Hall: London, UK, 1993; pp. 23–46. [Google Scholar]
- Freitas, R.H.A.; Volpato, G.L. Behavioral response of Nile tilapia to an allopatric predator. Mar. Freshw. Behav. Physiol. 2008, 41, 267–272. [Google Scholar] [CrossRef]
- Giaquinto, P.C.; Volpato, G.L. Chemical Communication, Aggression, and Conspecific Recognition in the Fish Nile Tilapia. Physiol. Behav. 1997, 62, 1333–1338. [Google Scholar] [CrossRef]
- Rucinque, D.S.; Ferreira, P.F.; Leme, P.R.P.; Lapa-Guimarães, J.; Viegas, E.M.M. Ocimum americanum and Lippia alba essential oils as anaesthetics for Nile tilapia: Induction, recovery of apparent unconsciousness and sensory analysis of fillets. Aquaculture 2021, 531, 735902. [Google Scholar] [CrossRef]
- Algers, B.; Blokhuis, H.J.; Bøtner, A.; Broom, D.M.; Costa, P.; Domingo, M.; Greiner, M.; Hartung, J.; Koenen, F.; Müller-Graf, C.; et al. Scientific opinion. Species-specific welfare aspects of the main systems of stunning and killing of farmed carp Scientific Opinion of the Panel on Animal Health andWelfare (Question N◦ EFSA-Q-2008-439). EFSA J. 2009, 1013, 1–37. [Google Scholar]
- Mackerth, F. Water analysis: Some revised methods for limnologists. Freshw. Biol. Assoc. Sci. Publ. 1978, 36, 117. [Google Scholar]
- Van De Vis, H.; Kestin, S.; Robb, D.; Oehlenschläger, J.; Lambooij, B.; Münkner, W.; Kuhlmann, H.; Kloosterboer, K.; Tejada, M.; Huidobro, A.; et al. Is humane slaughter of fish possible for industry? Aquac. Res. 2003, 34, 211–220. [Google Scholar] [CrossRef]
- Zahl, I.H.; Samuelsen, O.; Kiessling, A. Anaesthesia of farmed fish: Implications for welfare. Fish Physiol. Biochem. 2012, 38, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Anil, M.H. Studies on the return of physical reflexes in pigs following electrical stunning. Meat Sci. 1991, 30, 13–21. [Google Scholar] [CrossRef]
- Laming, P.R. Behavioural arousal and its habituation in the squirrel fish, Holocentrus rufus: The role of the telecephalon. Behav. Neural Biol. 1987, 47, 80–104. [Google Scholar] [CrossRef]
- Noble, C.; Gismervik, K.; Iversen, M.H.; Kolarevic, J.; Nilsson, J.; Stien, L.H.; Turnbull, J.F. Welfare Indicators for Farmed Atlantic Salmon: Tools for Assessing Fish Welfare; Nofima: Tromsø, Norway, 2018; p. 351. ISBN 9788282965569. [Google Scholar]






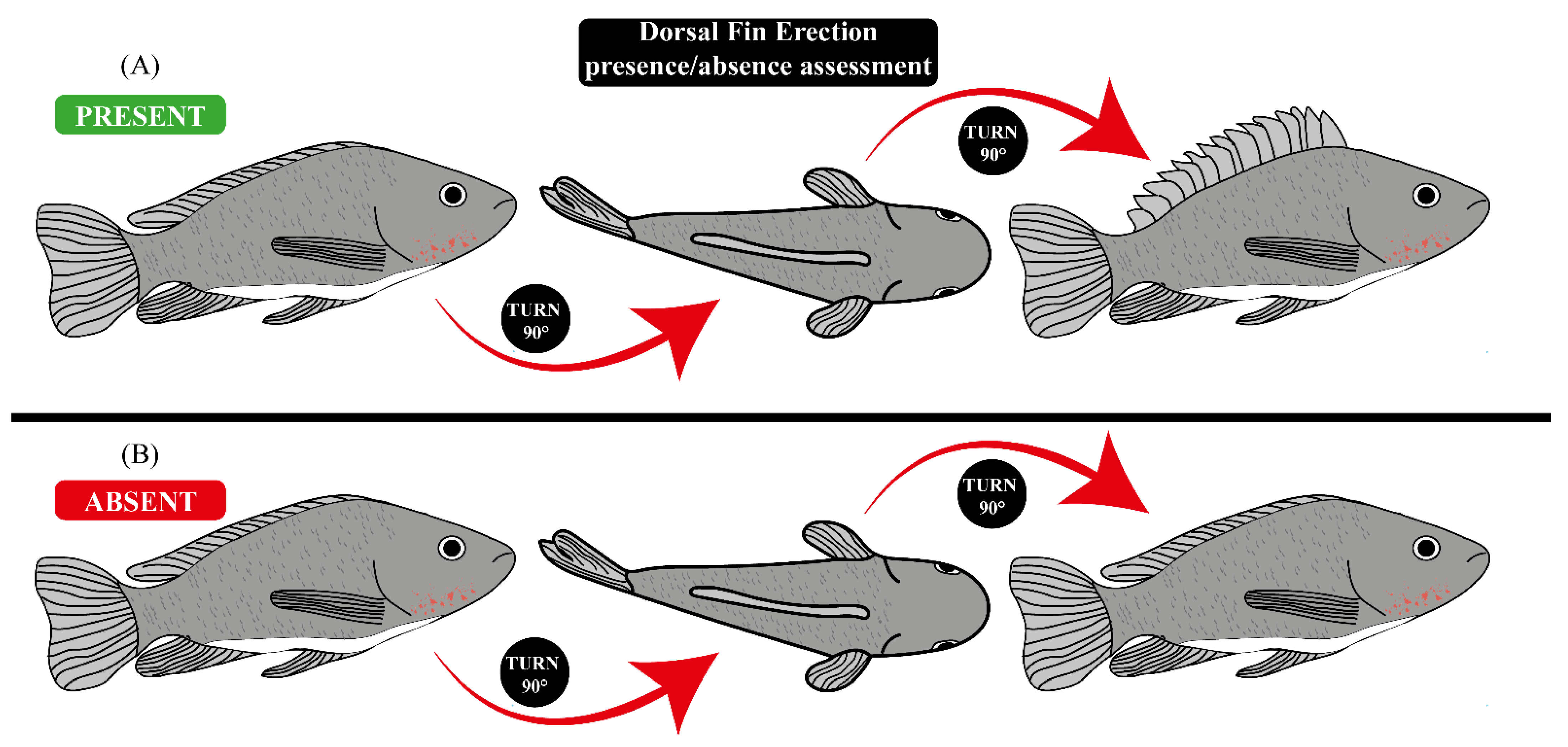{kind=link}
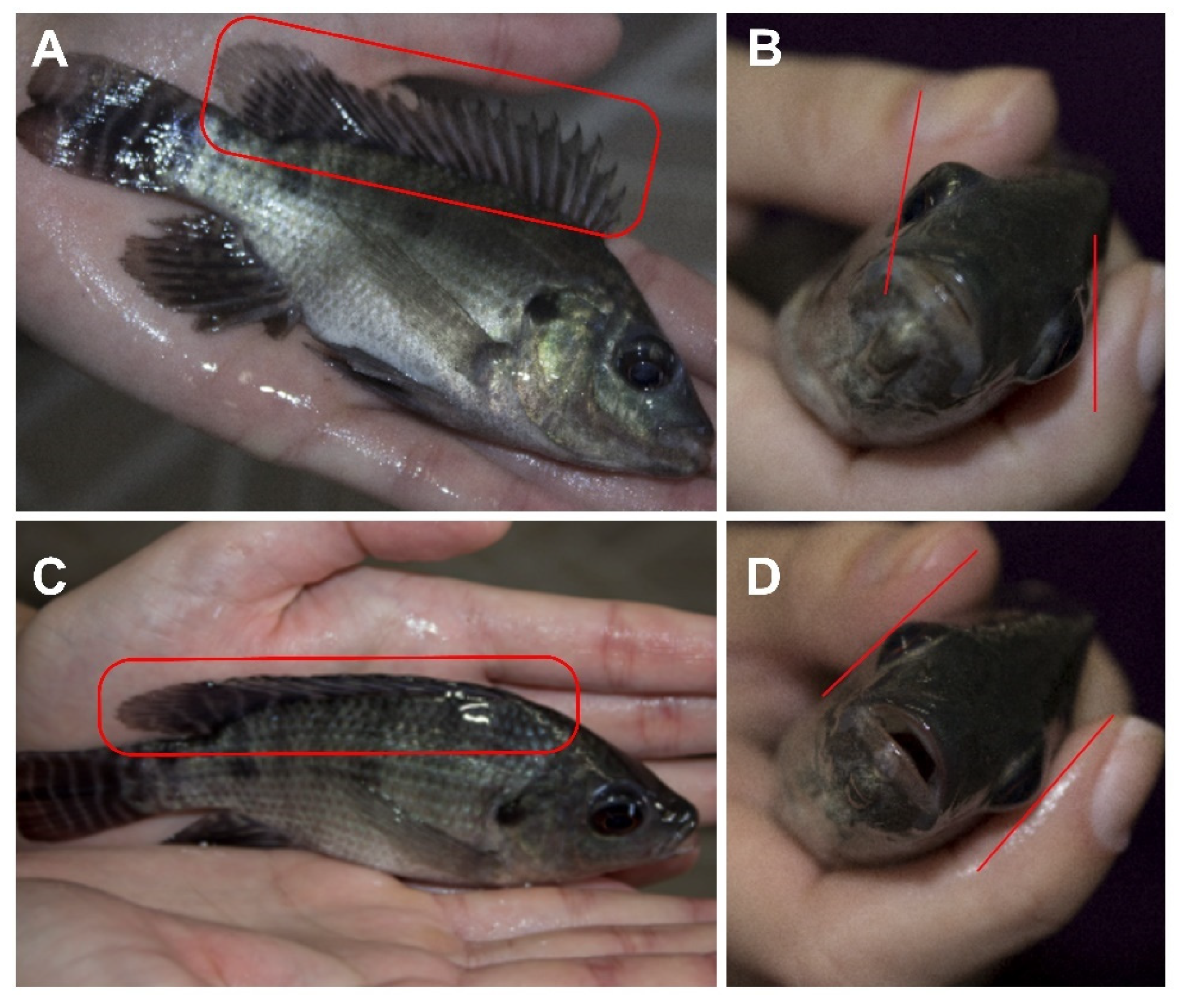{kind=link}
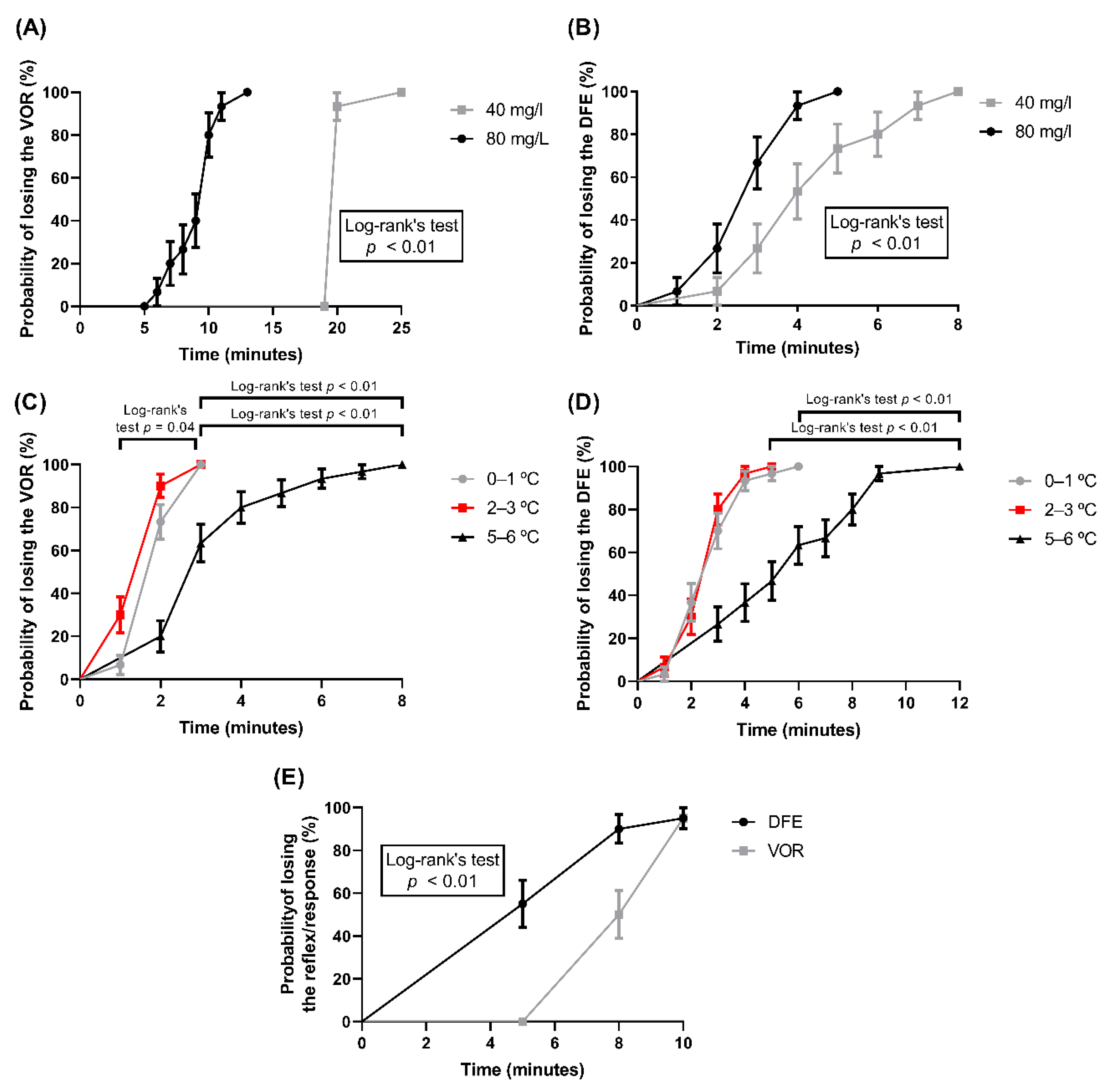{kind=link}
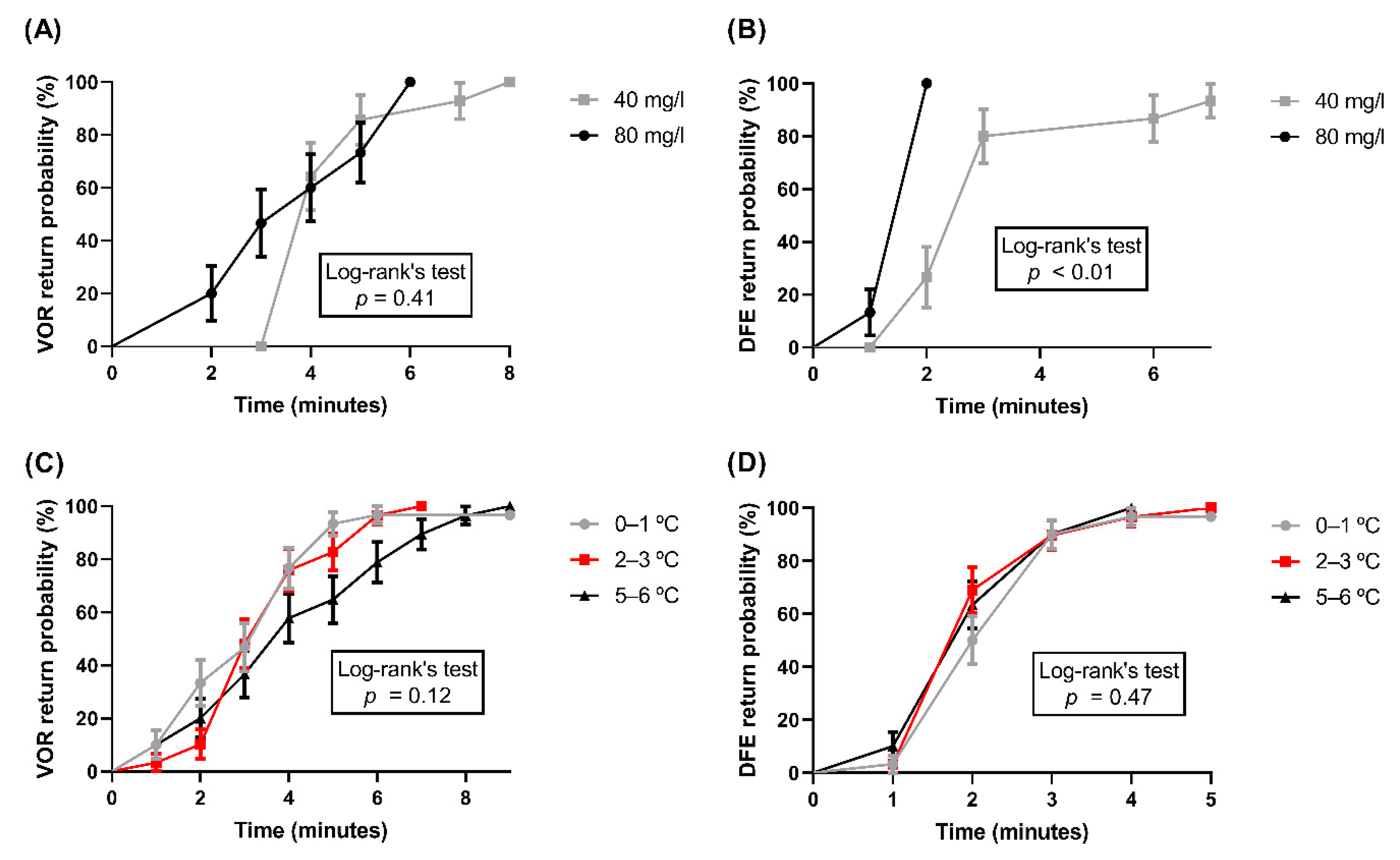{kind=link}
{kind=link}
| Stunning Protocol | VOR 1 | DFE 2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lost | Return | Lost | Return | |||||||||
| Range (min) | Median (min) | Fish (%) | Range (min) | Median (min) | Fish (%) | Range (min) | Median (min) | Fish (%) | Range (min) | Median (min) | Fish (%) | |
| Benzocaine (40 mg/L) | 20–25 | 20 | 100 | 0–8 | 4 | 100 | 2–8 | 4 | 100 | 0–7 | 3 | 93.33 |
| Benzocaine (80 mg/L) | 6–13 | 10 | 100 | 2–6 | 4 | 100 | 1–5 | 3 | 100 | 1–2 | 2 | 100 |
| Ice water (0–1 °C) | 1–3 | 2 | 100 | 1–6 | 4 | 96.67 | 1–6 | 3 | 100 | 1–4 | 2.5 | 96.67 |
| Ice water (2–3 °C) | 1–3 | 2 | 100 | 1–7 | 4 | 96.67 | 1–5 | 3 | 100 | 1–5 | 2 | 96.67 |
| Ice water (5–6 °C) | 2–8 | 3 | 100 | 2–9 | 4 | 96.67 | 3–12 | 6 | 100 | 1–4 | 2 | 96.67 |
| CO2 stunning | 8–10 | 9 | 95 | 5–5 | 5 | 100 | 5–10 | 5 | 95 | 5–5 | 5 | 100 |
| Stunning Protocol | Lost | Return | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| None VSI 1 (%) | Only One VSI (%) | Both VSI (%) | VOR 2 before DFE 3 (%) | DFE before VOR (%) | Both VSI at Same Time (%) | None VSI (%) | Only One VSI (%) | Both VSI (%) | VOR before DFE (%) | DFE before VOR (%) | Both VSI at Same Time (%) | |
| Benzocaine (40 mg/L) | 0 | 0 | 100 | 0 | 100 | 0 | 0 | 0 | 100 | 0 | 93.33 | 6.67 |
| Benzocaine (80 mg/L) | 0 | 0 | 100 | 0 | 100 | 0 | 0 | 0 | 100 | 0 | 86.67 | 13.33 |
| Ice water (0–1 °C) | 0 | 0 | 100 | 66.67 | 6.67 | 26.67 | 0 | 0 | 100 | 10.35 | 58.62 | 31.03 |
| Ice water (2–3 °C) | 0 | 0 | 100 | 80 | 3.3 | 16.67 | 0 | 0 | 100 | 3.45 | 86.21 | 10.34 |
| Ice water (5–6 °C) | 0 | 0 | 100 | 83.33 | 0 | 16.67 | 0 | 0 | 100 | 3.45 | 79.31 | 17.24 |
| CO2 stunning | 5 | 0 | 95 | 0 | 90 | 10 | 0 | 0 | 100 | 0 | 0 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camargo-dos-Santos, B.; Carlos, C.L.; Favero-Neto, J.; Alves, N.P.C.; Gonçalves, B.B.; Giaquinto, P.C. Welfare in Nile Tilapia Production: Dorsal Fin Erection as a Visual Indicator for Insensibility. Animals 2021, 11, 3007. https://doi.org/10.3390/ani11103007
Camargo-dos-Santos B, Carlos CL, Favero-Neto J, Alves NPC, Gonçalves BB, Giaquinto PC. Welfare in Nile Tilapia Production: Dorsal Fin Erection as a Visual Indicator for Insensibility. Animals. 2021; 11(10):3007. https://doi.org/10.3390/ani11103007
Chicago/Turabian StyleCamargo-dos-Santos, Bruno, Clarissa Lerois Carlos, João Favero-Neto, Nina Pacheco Capelini Alves, Bruno Bastos Gonçalves, and Percília Cardoso Giaquinto. 2021. "Welfare in Nile Tilapia Production: Dorsal Fin Erection as a Visual Indicator for Insensibility" Animals 11, no. 10: 3007. https://doi.org/10.3390/ani11103007
APA StyleCamargo-dos-Santos, B., Carlos, C. L., Favero-Neto, J., Alves, N. P. C., Gonçalves, B. B., & Giaquinto, P. C. (2021). Welfare in Nile Tilapia Production: Dorsal Fin Erection as a Visual Indicator for Insensibility. Animals, 11(10), 3007. https://doi.org/10.3390/ani11103007