
A Disorder of Sex Development in a Holstein–Friesian Heifer with a Rare Mosaicism (60,XX/90,XXY): A Genetic, Anatomical, and Histological Study

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Case Presentation
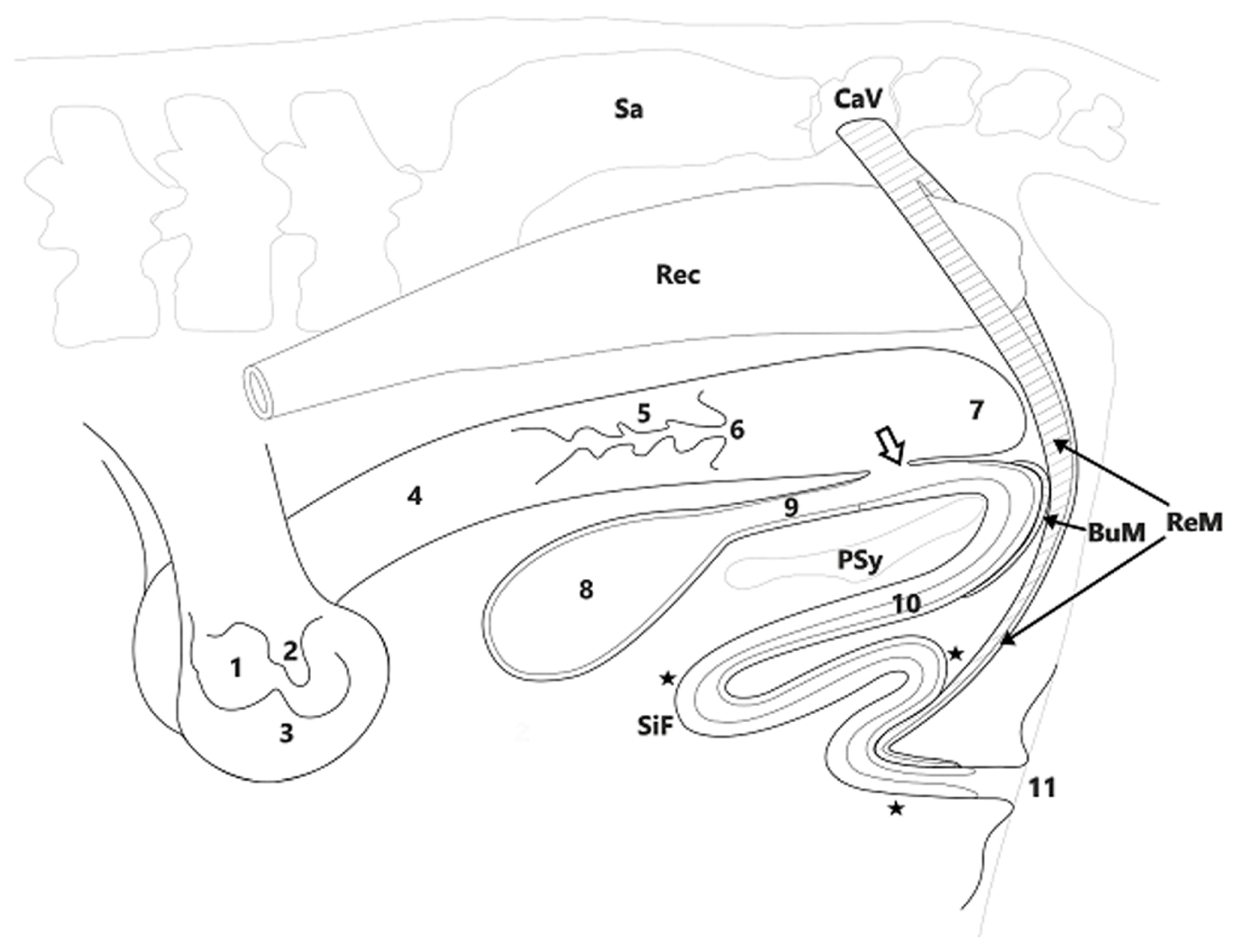
2.2. Anatomical and Histopathological Studies
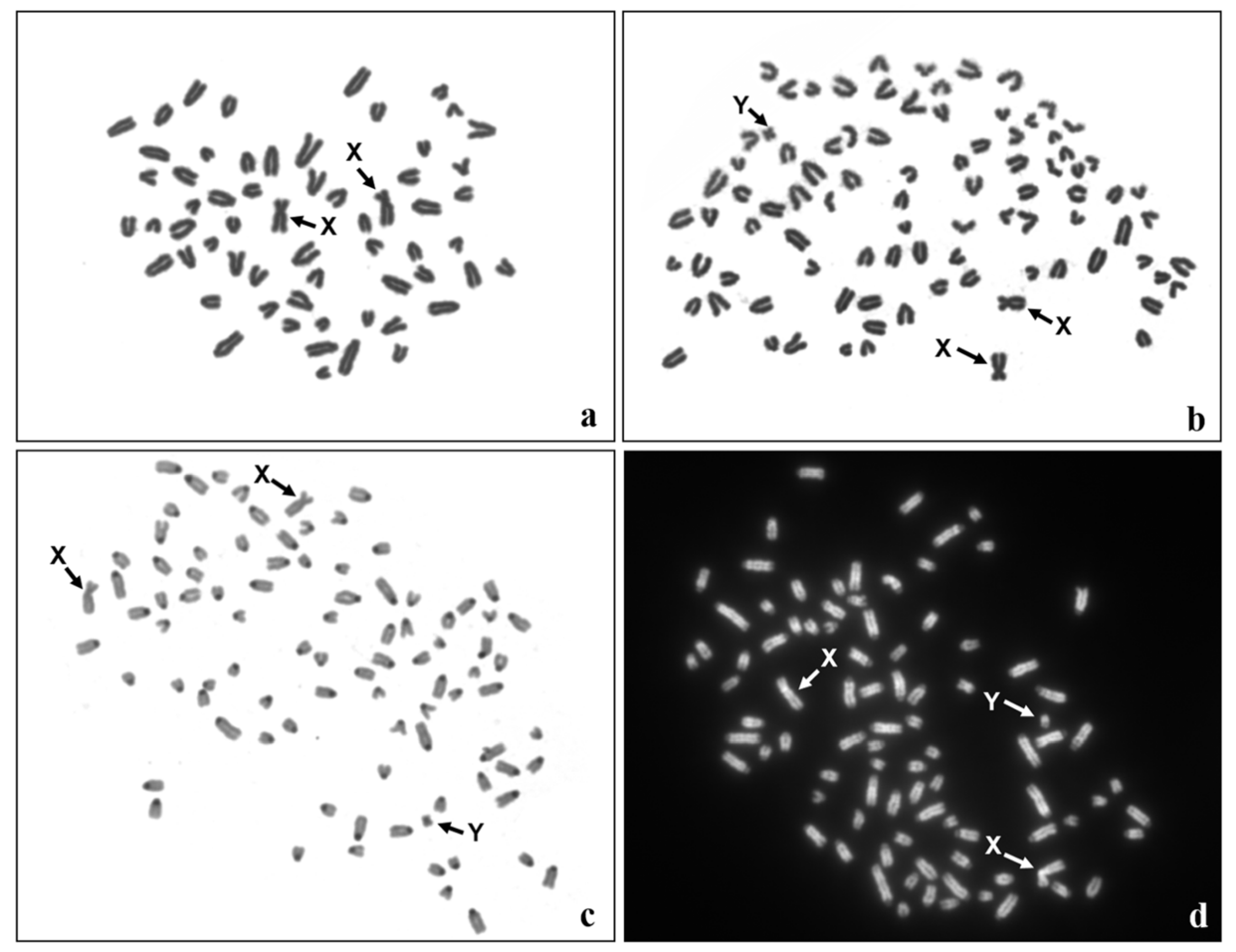
2.3. Cytogenetic Analysis
2.4. Molecular Analysis
3. Results
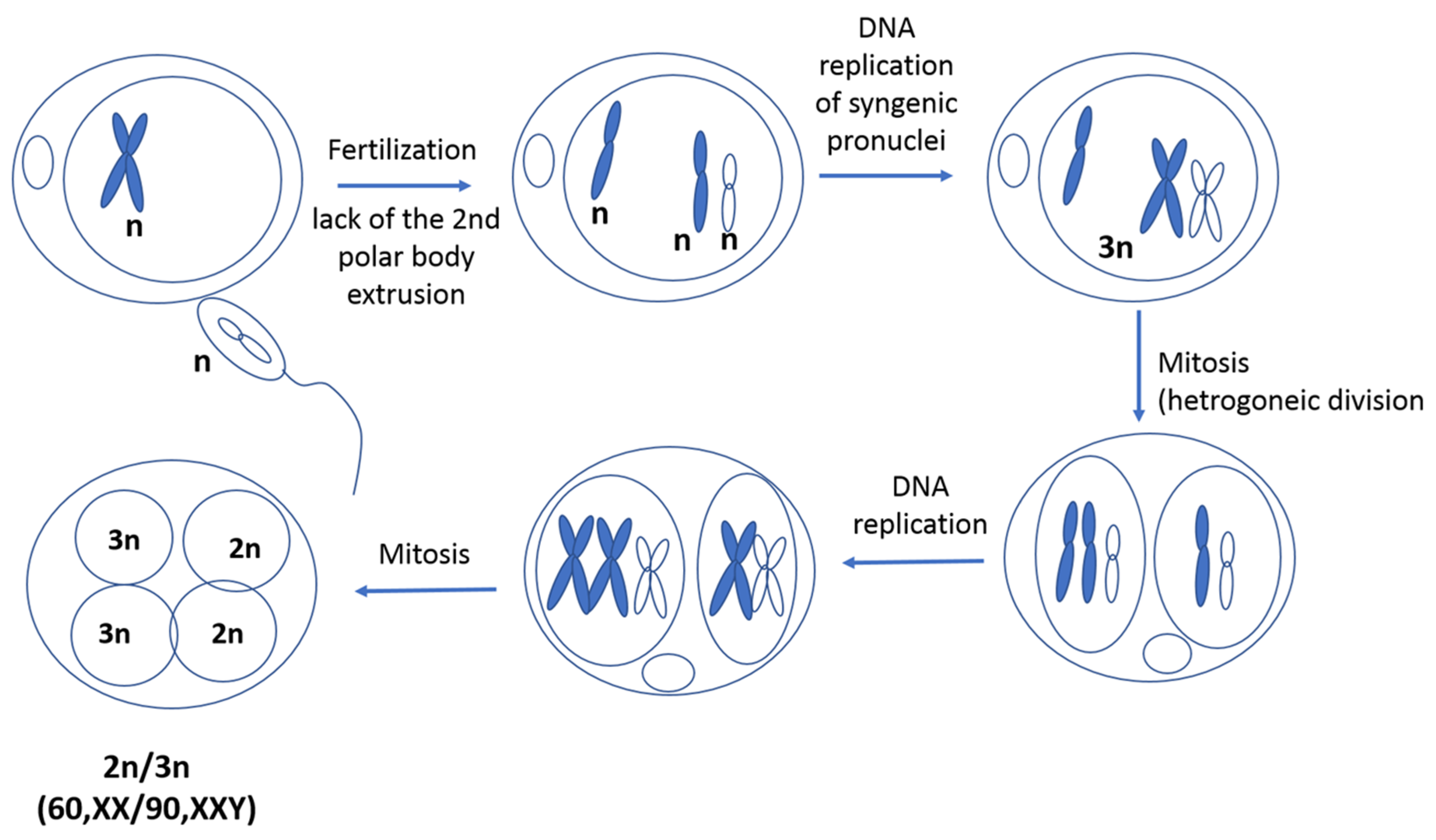
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Parma, P.; Veyrunes, F.; Pailhoux, E. Sex reversal in non-human placental mammals. Sex. Dev. 2016, 10, 326–344. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Switonski, M. Chromosome Abnormalities in Domestic Animals as Causes of Disorders of Sex Development or Impaired Fertility. In Insights from Animal Reproduction; Payan-Careira, R., Ed.; InTech: Rijeka, Croatia, 2016; pp. 207–225. [Google Scholar]
- Raudsepp, T.; Chowdhary, B.P. Chromosome aberrations and fertility disorders in domestic animals. Annu. Rev. Anim. Biosci. 2016, 4, 15–43. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Nowacka-Woszuk, J.; Albarella, S.; Switonski, M. Technical note: Droplet digital PCR as a new molecular method for a simple and reliable diagnosis of freemartinism in cattle. J. Dairy Sci. 2019, 102, 10100–10104. [Google Scholar] [CrossRef] [PubMed]
- Kozubska-Sobocińska, A.; Smołucha, G.; Danielak-Czech, B. Early diagnostics of freemartinism in Polish Holstein-Friesian female calves. Animals 2019, 9, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, H.O.; McEntee, K.; Hansel, W. Diploid-triploid chimerism in a bovine true hermaphrodite. Cytogenet. Genome Res. 1970, 9, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Rieck, G.W.; Höhn, H.; Schmidt, I. Vulvaaplasie und urethra masculina: Maskulinisierungseffekte des sinus urogenitalis bei genetisch weiblichen rindern durch chimärismen mit dem XXY-gonosomen-komplement [Aplastic vulva and masculine urethra: Masculinization effects of the urogenital sinus in genetically female cattle due to chimerism with the XXY gonosomal complement]. Berl. Munch. Tierarztl. Wochenschr. 1982, 95, 181–185. [Google Scholar]
- Meinecke, B.; Drogemuller, C.; Kuiper, H.; Burstel, D.; Wohlsein, P.; Ebeling, S.; Wehrend, A.; Meinecke-Tillmann, S. A diploid-triploid (60, XX/90, XXY) intersex in a Holstein heifer. Sex. Dev. 2007, 1, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Chu, E.H.; Thuline, H.C.; Norby, D.E. Triploid-diploid chimerism in a male tortoiseshell cat. Cytogenet. Genome Res. 1964, 3, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gregson, N.M.; Ishmael, J. Diploid-triploid chimerism in 3 tortoiseshell cats. Res. Vet. Sci. 1971, 2, 275–279. [Google Scholar] [CrossRef]
- Power, M.M.; Leadon, D.P. Diploid-triploid chimaerism (64, XX/96, XXY) in an intersex foal. Equine Vet. J. 1990, 22, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.C.; Hoffner, L.; Conlin, L.; Parks, W.T.; Fisher, R.A.; Spinner, N.; Yatsenko, S.A.; Bonadio, J.; Surti, U. Diploid/triploid mixoploidy: A consequence of asymmetric zygotic segregation of parental genomes. Am. J. Med. Genet. Part A 2018, 176, 2720–2732. [Google Scholar] [CrossRef] [PubMed]
- Van de Laar, I.; Rabelink, G.; Hochstenbach, R.; Tuerlings, J.; Hoogeboom, J.; Giltay, J. Diploid/triploid mosaicism in dysmorphic patients. Clin. Genet. 2002, 62, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Boonen, S.E.; Hoffmann, A.L.; Donnai, D.; Tümer, Z.; Ravn, K. Diploid/triploid mosaicism: A rare event or an under-diagnosed syndrome? Eur. J. Med. Genet. 2011, 54, 374–375. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Kociucka, B.; Nowacka-Woszuk, J.; Lach, Z.; Jaskowski, J.M.; Switonski, M. A high incidence of leukocyte chimerism (60, XX/60, XY) in single born heifers culled due to underdevelopment of internal reproductive tracts. Czech. J. Anim. Sci. 2014, 59, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Padula, A.M. The freemartin syndrome: An update. Anim. Reprod. Sci. 2005, 87, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Peretti, V.; Ciotola, F.; Albarella, S.; Paciello, O.; Dario, C.; Barbieri, V.; Iannuzzi, L. XX/XY chimerism in cattle: Clinical and cytogenetic studies. Sex. Dev. 2008, 2, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Destouni, A.; Zamani Esteki, M.; Catteeuw, M.; Tšuiko, O.; Dimitriadou, E.; Smits, K.; Kurg, A.; Salumets, A.; Van Soom, A.; Voet, T.; et al. Zygotes segregate entire parental genomes in distinct blastomere lineages causing cleavage-stage chimerism and mixoploidy. Genome. Res. 2016, 26, 567–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nes, N. Diploid-triploid chimerism in a true hermaphrodite mink (Mustela vison). Hereditas 1966, 56, 159–170. [Google Scholar] [CrossRef]
- Szczerbal, I.; Nowacka-Woszuk, J.; Dzimira, S.; Matuszczyk, A.; Iskrzak, P.; Switonski, M. Elevated incidence of freemartinism in pigs detected by droplet digital PCR and cytogenetic techniques. Livest. Sci. 2019, 219, 52–56. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytogenetic Analysis | ||||
| Tissue | Number of metaphase spreads | |||
| Total | 60,XX | 90,XXY | ||
| Lymphocytes | 400 | 388 | 12 | |
| Fibroblasts | 200 | 130 | 70 | |
| Molecular analysis | ||||
| PCR | ddPCR | |||
| SRY | AMELX | AMELY | ||
| Blood | + | + | + | |
| Skin | + | + | + | |
| Fibroblasts | + | + | + | |
| buccal epithelial cells | + | + | + | |
| hair follicles | not detectable | + | + | |
| Species | Karyotype | Information about Frequency of Triploid Metaphases in Different Tissues | Phenotype | Reference |
|---|---|---|---|---|
| Cattle | 60,XX/90,XXY | very low in leukocytes and elevated in fibroblasts (different organs) | Penis, empty scrotum, ovary with corpus luteum, ovotestis with epididymis; ductus deferens; uterus | [6] |
| 60,XX/90,XXY | absent from leukocytes and bone marrow, present in fibroblasts | aplasia of vulva, rudimentary penis, ovaries | [7] | |
| 60,XX/90,XXY | aplasia of vulva, rudimentary penis | [7] | ||
| 60,XX/90,XXY | absent from leukocytes, elevated in fibroblasts | aplastic vulva, penis and clitoris agenesis, a male-like urethra located in a pseudoprepuce opening between the mammary complexes, cycling ovaries | [8] | |
| 60,XX/90,XXY | low in leukocytes, elevated in fibroblasts | deformed vulva, blind vagina, extra urethra included in the shortened penis with sigmoid flexure; presence of bulbospongiosus muscle and retractor muscle of penis | This study | |
| Cat | 38,XX/57,XXY | not studied in leukocytes, present in fibroblasts (ear) | tortoiseshell male cat with spermatogenetically inactive testes | [9] |
| 38,XX/57,XXY | absent from leukocytes and low in fibroblasts (skin) | rudimentary penis, female genital tract, hypoplastic testis | [10] | |
| 38,XX/57,XXY | not studied in leukocytes, present in fibroblasts (testes) | tortoiseshell male cat with spermatogenetically inactive testes | ||
| 38,XY/57,XXY | not studied in leukocytes, low in fibroblasts (skin) | tortoiseshell male cat with spermatogenetically active testes | ||
| Horse | 64,XX/96,XXY | absent from leukocytes and very high in fibroblasts (skin) | enlarged clitoris, with partially sealed vulva lips, urinary tract orifice in a vestigial vagina | [11] |
| Mink | 30,XX/45,XXY | not studied in leukocytes, elevated in fibroblasts derived from different organs | enlarged clitoris with os penis, uterus, ovotestes | [19] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczerbal, I.; Komosa, M.; Nowacka-Woszuk, J.; Uzar, T.; Houszka, M.; Semrau, J.; Musial, M.; Barczykowski, M.; Lukomska, A.; Switonski, M. A Disorder of Sex Development in a Holstein–Friesian Heifer with a Rare Mosaicism (60,XX/90,XXY): A Genetic, Anatomical, and Histological Study. Animals 2021, 11, 285. https://doi.org/10.3390/ani11020285
Szczerbal I, Komosa M, Nowacka-Woszuk J, Uzar T, Houszka M, Semrau J, Musial M, Barczykowski M, Lukomska A, Switonski M. A Disorder of Sex Development in a Holstein–Friesian Heifer with a Rare Mosaicism (60,XX/90,XXY): A Genetic, Anatomical, and Histological Study. Animals. 2021; 11(2):285. https://doi.org/10.3390/ani11020285
Chicago/Turabian StyleSzczerbal, Izabela, Marcin Komosa, Joanna Nowacka-Woszuk, Tomasz Uzar, Marek Houszka, Jerzy Semrau, Magdalena Musial, Michal Barczykowski, Anna Lukomska, and Marek Switonski. 2021. "A Disorder of Sex Development in a Holstein–Friesian Heifer with a Rare Mosaicism (60,XX/90,XXY): A Genetic, Anatomical, and Histological Study" Animals 11, no. 2: 285. https://doi.org/10.3390/ani11020285
APA StyleSzczerbal, I., Komosa, M., Nowacka-Woszuk, J., Uzar, T., Houszka, M., Semrau, J., Musial, M., Barczykowski, M., Lukomska, A., & Switonski, M. (2021). A Disorder of Sex Development in a Holstein–Friesian Heifer with a Rare Mosaicism (60,XX/90,XXY): A Genetic, Anatomical, and Histological Study. Animals, 11(2), 285. https://doi.org/10.3390/ani11020285