
Paternal Dietary Methionine Supplementation Improves Carcass Traits and Meat Quality of Chicken Progeny

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Diets
2.2. Samples Collection and Carcass Traits
2.3. Meat Quality Traits
2.4. RNA Isolation and Real-Time Quantitative qRT-PCR
2.5. Statistical Analysis
3. Results
3.1. Effects of Paternal Methionine on Carcass Traits of Progeny
3.2. Effects of Paternal Methionine on Meat Quality of Progeny
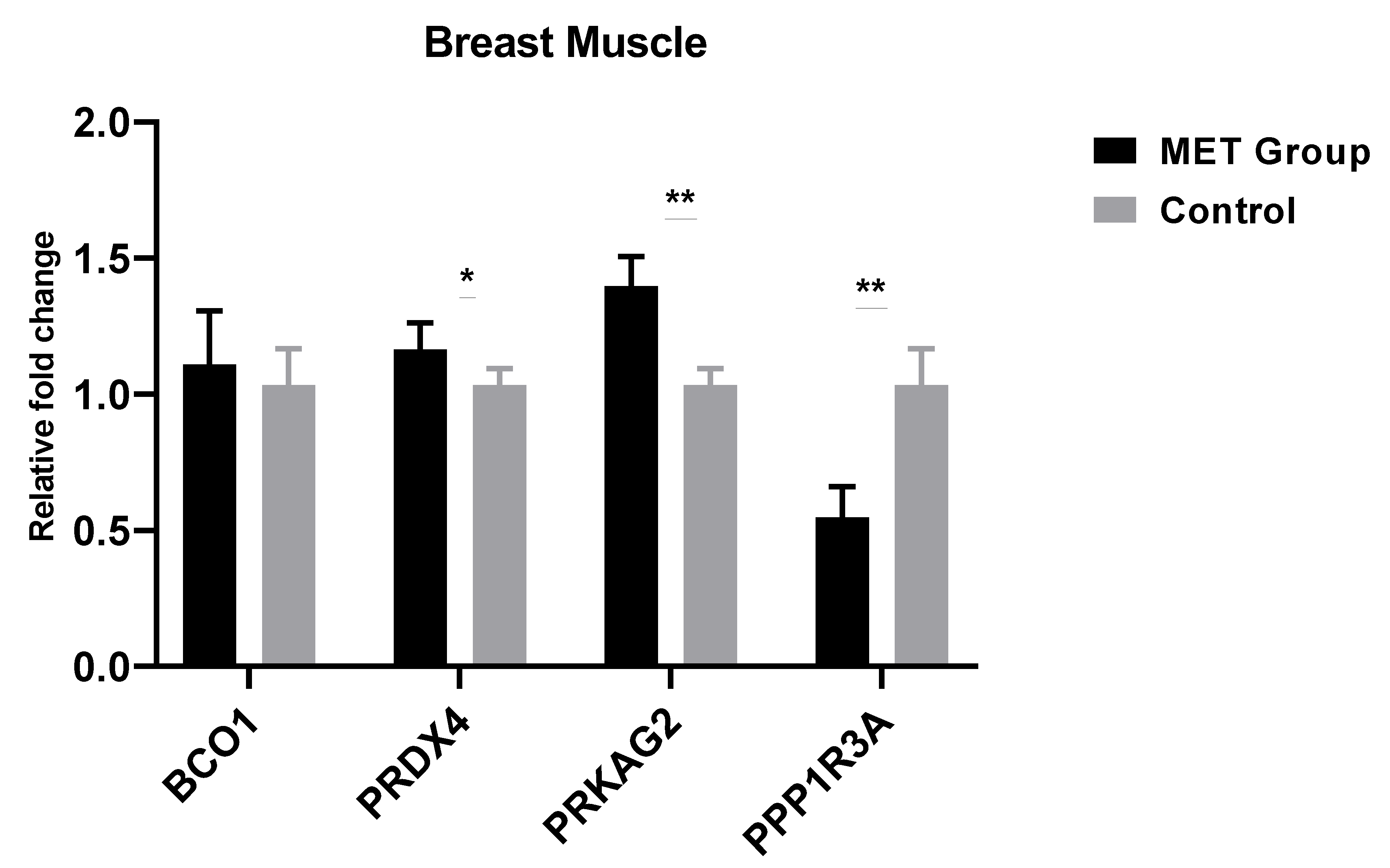
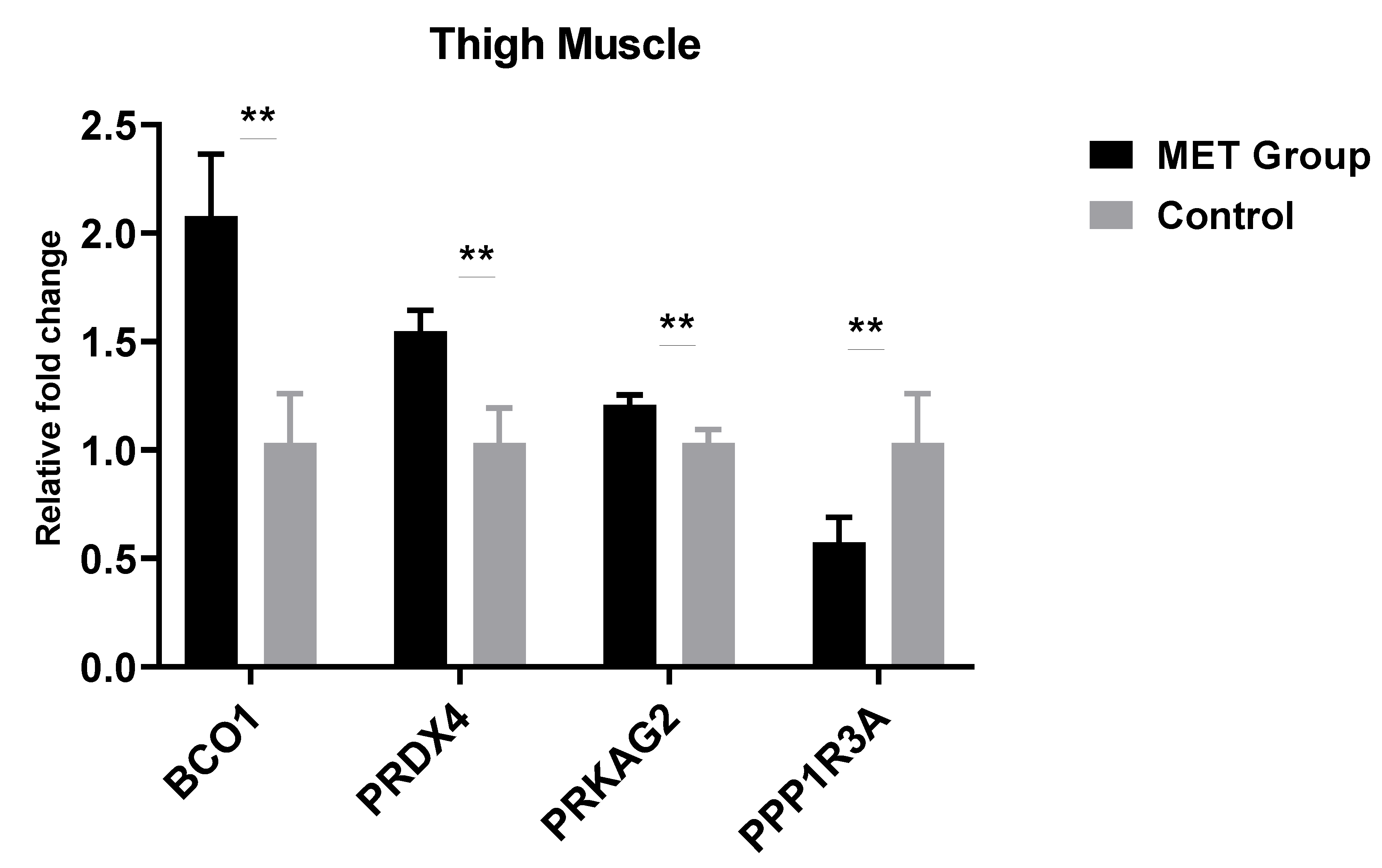
3.3. Effects of Methionine on Gene Expression of Breast and Thigh Muscles of Male’s Progeny
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Fanatico, A.C.; Pillai, P.B.; Emmert, J.L.; Owens, C.M. Meat quality of slow- and fast-growing chicken genotypes fed low-nutrient or standard diets and raised indoors or with outdoor access. Poult. Sci. 2007, 86, 2245–2255. [Google Scholar] [CrossRef] [PubMed]
- Dransfield, E.; Sosnicki, A.A. Relationship between muscle growth and poultry meat quality. Poult. Sci. 1999, 78, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, D.L. Poultry meat quality. World Poult. Sci. J. 2002, 58, 131–145. [Google Scholar] [CrossRef]
- Zhao, J.P.; Zhao, G.P.; Jiang, R.R.; Zheng, M.Q.; Chen, J.L.; Liu, R.R.; Wen, J. Effects of diet-induced differences in growth rate on metabolic, histological, and meat-quality properties of 2 muscles in male chickens of 2 distinct broiler breeds. Poult. Sci. 2012, 91, 237–247. [Google Scholar] [CrossRef]
- Drażbo, A.; Kozłowski, K.; Chwastowska-Siwiecka, I.; Sobczak, A.; Kwiatkowski, P.; Lemme, A. Effect of different dietary levels of DL-methionine and the calcium salt of DL- 2-hydroxy-4-[methyl] butanoic acid on the growth performance, carcass yield and meat quality of broiler chickens. Eur. Poult. Sci. 2015, 79, 1–14. [Google Scholar] [CrossRef]
- Wen, C.; Jiang, X.Y.; Ding, L.R.; Wang, T.; Zhou, Y.M. Effects of dietary methionine on growth performance, meat quality and oxidative status of breast muscle in fast- and slow-growing broilers. Poult. Sci. 2017, 96, 1707–1714. [Google Scholar] [CrossRef]
- Alagawany, M.; Abd El-Hack, M.E.; Arif, M.; Ashour, E.A. Individual and combined effects of crude protein, methionine, and probiotic levels on laying hen productive performance and nitrogen pollution in the manure. Environ. Sci. Pollut. Res. Int. 2016, 23, 22906–22913. [Google Scholar] [CrossRef]
- Bunchasak, C. Role of dietary methionine in poultry production. J. Poult. Sci. 2009, 46, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Niculescu, M.D.; Zeisel, S.H. Diet, methyl donors and DNA methylation: Interactions between dietary folate, methionine and choline. J. Nutr. 2002, 132, 2333S–2335S. [Google Scholar] [CrossRef]
- Schutte, J.B.; De Jong, J.; Smink, W.; Pack, M. Replacement value of betaine for DL-methionine in male broiler chicks. Poult. Sci. 1997, 76, 321–325. [Google Scholar] [CrossRef]
- Binder, M. Life cycle analysis of DL-methionine in broiler meat production. Amino News 2003, 4, 1–8. [Google Scholar]
- Elwan, H.A.M.; Elnesr, S.S.; Xu, Q.; Xie, C.; Dong, X.; Zou, X. Effects of In Ovo Methionine-Cysteine Injection on Embryonic Development, Antioxidant Status, IGF-I and TLR4 Gene Expression, and Jejunum Histomorphometry in Newly Hatched Broiler Chicks Exposed to Heat Stress during Incubation. Animals (Basel) 2019, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nockels, C. Protective effects of supplemental vitamin E against infection. Fed. Proc. 1979, 38, 2134–2138. [Google Scholar] [PubMed]
- Gao, J.; Lv, Z.; Li, C.; Yue, Y.; Zhao, X.; Wang, F.; Guo, Y. Maternal zinc supplementation enhanced skeletal muscle development through increasing protein synthesis and inhibiting protein degradation of their offspring. Biol. Trace. Elem. Res. 2014, 162, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Bosagna, C.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of phenotype and disease. Mol. Cell Endocrinol. 2012, 354, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Tan, X.; Zhao, G.; Chen, Y.; Zhao, D.; Li, W.; Zheng, M.; Wen, J. Maternal dietary methionine supplementation influences egg production, and the growth performance and meat quality of offspring. Poult. Sci. 2020, 99, 3550–3556. [Google Scholar] [CrossRef]
- Ruan, D.; Fouad, A.M.; Fan, Q.; Xia, W.; Wang, S.; Chen, W.; Lin, C.; Wang, Y.; Yang, L.; Zheng, C. Effects of dietary methionine on productivity, reproductive performance, antioxidant capacity, ovalbumin and antioxidant-related gene expression in laying duck breeders. Br. J.Nutr. 2018, 119, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Kalvandi, O.; Sadeghi, A.; Karimi, A. Methionine supplementation improves reproductive performance, antioxidant status, immunity and maternal antibody transmission in breeder Japanese quail under heat stress conditions. Arch. Anim. Breed. 2019, 62, 275–286. [Google Scholar] [CrossRef]
- Hayat, Z.; Rehman, A.U.; Akram, K.; Farooq, U.; Saleem, G. Evaluation of a natural methionine source on broiler growth performance. J. Sci. Food. Agric. 2015, 95, 2462–2466. [Google Scholar] [CrossRef]
- Wen, C.; Jiang, X.; Ding, L.; Wang, T.; Zhou, Y. Effects of dietary methionine on breast muscle growth, myogenic gene expression and IGF-I signaling in fast- and slow-growing broilers. Sci. Rep. 2017, 7, 1924. [Google Scholar] [CrossRef]
- Zhang, S.; Saremi, B.; Gilbert, E.R.; Wong, E.A. Physiological and biochemical aspects of methionine isomers and a methionine analogue in broilers. Poult. Sci. 2017, 96, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Conde-Aguilera, J.A.; Cholet, J.C.; Lessire, M.; Mercier, Y.; Tesseraud, S.; van Milgen, J. The level and source of free-methionine affect body composition and breast muscle traits in growing broilers. Poult. Sci. 2016, 95, 2322–2331. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.S.; Teshima, S.-I.; Koshio, S.; Ishikawa, M.; Uyan, O.; Hernandez, L.H.H.; Michael, F.R. Supplemental effects of coated methionine and/or lysine to soy protein isolate diet for juvenile kuruma shrimp, Marsupenaeus japonicus. Aquaculture 2005, 248, 13–19. [Google Scholar] [CrossRef]
- Smith, S.; Boling, J. Lipid coating as a mode of protecting free methionine from ruminal degradation. J. Anim. Sci. 1984, 58, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triques, G.E.; Cristo, A.B.D.; Canevese, M.; Marques, P.F.D.S.; Burin Junior, A.M.; Fernandes, J.I.M. Effect of Antioxidant Supplementation in Diets of Roosters during the Post-Peak Phase on Reproduction and Production Characteristics of Offspring. Ciênc. Anim. Bras. 2019, 20, 1–13. [Google Scholar] [CrossRef]
- Alderey, A.A.; Nasr, A.M.E.; Abu khashaba, H.A.; Samak, H.R. Performance of Gimmizah Cockerels Fed Two Levels of Methionine and Lysine and Two Levels of Protein. J. Anim. Poult. Prod. Mansoura Univ. 2019, 10, 223–229. [Google Scholar] [CrossRef]
- Shanmugam, M.; Prakash, B.; Rao, S.V.R.; Panda, A.K. Effect of dietary energy and crude protein on semen parameters and fertility in layer breeders males. Indian J. Anim. Sci. 2016, 51. [Google Scholar] [CrossRef]
- Gómez-Polo, C.; Montero, J.; Gómez-Polo, M.; Martin Casado, A. Comparison of the CIELab and CIEDE 2000 color difference formulas on gingival color space. J. Prosthodont 2020, 29, 401–408. [Google Scholar] [CrossRef]
- O’brien, W.; Groh, C.; Boenke, K. A new, small-color-difference equation for dental shades. J. Dent. Res. 1990, 69, 1762–1764. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhao, G.; Liu, R.; Zheng, M.; Hu, Y.; Wu, D.; Zhang, L.; Li, P.; Wen, J. The identification of 14 new genes for meat quality traits in chicken using a genome-wide association study. BMC Genomics 2013, 14, 458. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, X.; Tan, H.; Zhang, D.; Zhang, H.; Wei, S.; Yan, H.C. Nutrient density and slaughter age have differential effects on carcase performance, muscle and meat quality in fast and slow growing broiler genotypes. Poult. Sci. 2013, 54, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Devatkal, S.K.; Vishnuraj, M.R.; Kulkarni, V.V.; Kotaiah, T. Carcass and meat quality characterization of indigenous and improved variety of chicken genotypes. Poult. Sci. 2018, 97, 2947–2956. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Z.; Liu, R.; Wang, J.; Zheng, M.; Li, Q.; Cui, H.; Zhao, G.; Wen, J. Alteration of Hepatic Gene Expression along with the Inherited Phenotype of Acquired Fatty Liver in Chicken. Genes (Basel) 2018, 9, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D.J.m. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.; Yi, Y.J.; Wickramasuriya, S.S.; Kim, E.; Shin, T.K.; Cho, H.M.; Kim, N.; Heo, J.M. Evaluation of sulphur amino acid requirement of male Korean native ducklings from hatch to 21 day of age. Br. Poult. Sci. 2017, 58, 272–277. [Google Scholar] [CrossRef]
- Majdeddin, M.; Golian, A.; Kermanshahi, H.; Michiels, J.; De Smet, S. Effects of methionine and guanidinoacetic acid supplementation on performance and energy metabolites in breast muscle of male broiler chickens fed corn-soybean diets. Br. Poult. Sci. 2019, 60, 554–563. [Google Scholar] [CrossRef]
- Rehman, A.U.; Arif, M.; Husnain, M.M.; Alagawany, M.; Abd El-Hack, M.E.; Taha, A.E.; Elnesr, S.S.; Abdel-Latif, M.A.; Othman, S.I.; Allam, A.A. Growth Performance of Broilers as Influenced by Different Levels and Sources of Methionine Plus Cysteine. Animals (Basel) 2019, 9, 1056. [Google Scholar] [CrossRef] [Green Version]
- Hickling, D.; Guenter, W.; Jackson, M.E. The effects of dietary methionine and lysine on broiler chicken performance and breast meat yield. Can. J. Anim. Sci. 1990, 70, 673–678. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Hua, Y.; Fu, W.-Z.; Zhou, F.; Yang, B.-B.; Xiao, J.-X.; Liu, M.-H.; Shao, Q.-J. Effects of supplementation coated lysine and methionine in mixture protein diets on growth performance, digestibility and serum biochemical indices of juvenile black sea bream, Acanthopagrus schlegelii. Turk. J. Fish. Aquat. Sci. 2014, 14, 633–642. [Google Scholar]
- Mandal, A.; Elangovan, A.; Johri, T.S. Comparing bio-efficacy of liquid DL-methionine hydroxy analogue free acid with DL-methionine in broiler chickens. Asian Austral. J. Anim. 2004, 17, 102–108. [Google Scholar] [CrossRef]
- Liu, Y.; Song, G.; Yi, G.; Hou, Y.; Huang, J.; Knight, C.D. Effect of supplementing 2-hydroxy-4-(methylthio) butanoic acid and DL-methionine in corn-soybean-cottonseed meal diets on growth performance and carcass quality of broilers. Asian Austral. J. Anim. 2006, 19, 1197–1205. [Google Scholar] [CrossRef]
- Zhan, X.A.; Li, J.X.; Xu, Z.R.; Zhao, R.Q. Effects of methionine and betaine supplementation on growth performance, carcase composition and metabolism of lipids in male broilers. Br. Poult. Sci. 2006, 47, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Woelfel, R.L.; Owens, C.M.; Hirschler, E.M.; Martinez-Dawson, R.; Sams, A.R. The characterization and incidence of pale, soft, and exudative broiler meat in a commercial processing plant. Poult. Sci. 2002, 81, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, A.; Herbert, U.; Miskel, D.; Heinemann, C.; Braun, C.; Dohlen, S.; Zeitz, J.O.; Eder, K.; Saremi, B.; Kreyenschmidt, J. Effect of methionine supplementation in chicken feed on the quality and shelf life of fresh poultry meat. Poult. Sci. 2017, 96, 2853–2861. [Google Scholar] [CrossRef]
- Zonenberg, Ł.; Drażbo, A. The effect of increased methionine in broiler chicken diets on the quality of breast muscles at different times of vacuum storage under refrigeration. Sci. Ann. Pol. Soc. Anim. Prod. 2018, 14, 49–60. [Google Scholar] [CrossRef]
- Albrecht, A.; Hebel, M.; Heinemann, C.; Herbert, U.; Miskel, D.; Saremi, B.; Kreyenschmidt, J. Assessment of Meat Quality and Shelf Life from Broilers Fed with Different Sources and Concentrations of Methionine. J. Food Qual. 2019, 2019, 6182580. [Google Scholar] [CrossRef]
- Wang, Z.G.; Pan, X.J.; Peng, Z.Q.; Zhao, R.Q.; Zhou, G.H. Methionine and selenium yeast supplementation of the maternal diets affects color, water-holding capacity, and oxidative stability of their male offspring meat at the early stage. Poult. Sci. 2009, 88, 1096–1101. [Google Scholar] [CrossRef]
- Mugler, D.; Cunningham, F.E. Factors affecting poultry meat color—A review. World Poult. Sci. J. 1972, 28, 400–406. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Nadaf, J.; Le Bihan-Duval, E.; Berri, C.; Dunn, I.; Talbot, R.; De Koning, D.-J. Using targeted resequencing for identification of candidate genes and SNPs for a QTL Affecting the pH value of chicken meat. G3 Genes Genomes Genet. 2015, 5, 2085–2089. [Google Scholar] [CrossRef] [Green Version]
- Nadaf, J.; Berri, C.; Dunn, I.; Godet, E.; Le Bihan-Duval, E.; De Koning, D.J. An expression QTL of closely linked candidate genes affects pH of meat in chickens. Genetics 2014, 196, 867–874. [Google Scholar] [CrossRef] [Green Version]
- Sibut, V.; Hennequet-Antier, C.; Le Bihan-Duval, E.; Marthey, S.; Duclos, M.J.; Berri, C. Identification of differentially expressed genes in chickens differing in muscle glycogen content and meat quality. BMC Genomics 2011, 12, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abasht, B.; Zhou, N.; Lee, W.R.; Zhuo, Z.; Peripolli, E. The metabolic characteristics of susceptibility to wooden breast disease in chickens with high feed efficiency. Poult. Sci. 2019, 98, 3246–3256. [Google Scholar] [CrossRef] [PubMed]
- Jlali, M.; Graulet, B.; Chauveau-Duriot, B.; Chabault, M.; Godet, E.; Leroux, S.; Praud, C.; Le Bihan-Duval, E.; Duclos, M.J.; Berri, C. A mutation in the promoter of the chicken β, β-carotene 15, 15′-monooxygenase 1 gene alters xanthophyll metabolism through a selective effect on its mRNA abundance in the breast muscle. J. Anim. Sci. 2012, 90, 4280–4288. [Google Scholar] [CrossRef] [PubMed]
- Le Bihan-Duval, E.; Nadaf, J.; Berri, C.; Pitel, F.; Graulet, B.; Godet, E.; Leroux, S.Y.; Demeure, O.; Lagarrigue, S.; Duby, C.J.P.O. Detection of a Cis eQTL controlling BMCO1 gene expression leads to the identification of a QTG for chicken breast meat color. PLoS ONE 2011, 6, e14825. [Google Scholar] [CrossRef]
- Chang, A.; Halley, J.; Silva, M. Can feeding the broiler breeder improve chick quality and offspring performance? Anim. Prod. Sci. 2016, 56, 1254–1262. [Google Scholar] [CrossRef]
- Schagdarsurengin, U.; Steger, K. Epigenetics in male reproduction: Effect of paternal diet on sperm quality and offspring health. Nat. Rev. Urol. 2016, 13, 584–595. [Google Scholar] [CrossRef]
- Zhang, N. Epigenetic modulation of DNA methylation by nutrition and its mechanisms in animals. Anim. Nutr. 2015, 1, 144–151. [Google Scholar] [CrossRef]
- Tehlivets, O.; Malanovic, N.; Visram, M.; Pavkov-Keller, T.; Keller, W. S-adenosyl-L-homocysteine hydrolase and methylation disorders: Yeast as a model system. Biochim. Biophys. Acta 2013, 1832, 204–215. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N. Role of methionine on epigenetic modification of DNA methylation and gene expression in animals. Anim. Nutr. 2018, 4, 11–16. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Item | Control Group | MET Group |
|---|---|---|
| Ingredient (%) | ||
| Corn | 65.37 | 65.37 |
| Wheat bran | 15.00 | 15.00 |
| Soybean meal, 44% CP | 13.60 | 13.50 |
| Soybean oil | 2.00 | 2.00 |
| Limestones | 1.50 | 1.50 |
| Dicalcium phosphate | 1.23 | 1.23 |
| Salt | 0.30 | 0.30 |
| Coated methionine | 0.00 | 0.10 |
| Premix 2 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 |
| Calculated composition | ||
| ME (Mcal/kg) | 2.87 | 2.87 |
| CP (%) | 13.82 | 13.82 |
| Ca (%) | 0.94 | 0.94 |
| TP (%) | 0.61 | 0.61 |
| Met (%) | 0.28 | 0.38 |
| Analyzed composition | ||
| CP (%) | 13.85 | 13.89 |
| Met (%) | 0.29 | 0.40 |
| Accession Numbers | Genes 1 | Primers | Amplicon Size (bp) |
|---|---|---|---|
| XM_015281206.2 | PRKAG2 | F:5’-TGCCTTCATACATCCAGACACTCCTAT-3’ | 279 |
| R:5’-ACCTCAGCCTTCACTATCCTATCAACA-3’ | |||
| XM_416800.6 | PRDX4 | F:5’-CCACCCTAGCCATGGATTACC-3’ | 197 |
| R:5’-AGGCATGGCTACATCTTCGAG-3’ | |||
| XM_015292519.2 | BCO1 | F:5’-TCCAACTTCCGCAACTGCTGTA-3’ | 314 |
| R:5’-TTGGCTCAGACACCACAACACA-3’ | |||
| XM_004937541.2 | PPP1R3A | F:5’-TGAACGGCATTATACGAGTCCTCAA-3’ | 195 |
| R:5’-ATTCCACTTTGGCTCCATCTCTCTG-3’ | |||
| XM_015293128.2 | RPL32 | F:5’-AGTTCATCCGCCACCAGTCTGAT-3’ | 147 |
| R:5’-GCTTCGTCTTCTTGTTGCTCCCATA-3’ | |||
| NM_001006685.1 | HSP70 | F:5’-TCTGCTCCTGTTGGATGTC-3’ | 95 |
| R:5’-TGGGAATGGTGGTGTTACG-3’ |
| Items | Treatments | Gender | SEM | Significance | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control | MET Group | Male | Female | Treatments | Gender | Interaction | |||
| Live weight(g) | 2002.5 a | 1942.9 b | 2009.8 A | 1871.7 B | 17.3 | * | ** | NS | |
| Eviscerated weight(g) | 1631.0 | 1600.0 | 1663.1 A | 1516.4 B | 14.9 | NS | ** | NS | |
| Eviscerated yield (%) | 81.47 b | 82.33 a | 82.79 A | 81.01 B | 0.22 | * | ** | NS | |
| Breast muscle (%) | 26.65 | 26.41 | 26.10 A | 26.97 B | 0.14 | NS | ** | NS | |
| Thigh muscle (%) | 29.09 b | 29.99 a | 29.74 A | 29.03 B | 0.09 | ** | ** | NS | |
| AF (%) | 0.97 a | 0.88 b | 0.83 B | 1.02 A | 0.02 | ** | ** | NS | |
| Items 4 | Treatments | Gender | SEM | Significance | ||||
|---|---|---|---|---|---|---|---|---|
| Control | MET Group | Male | Female | Treatments | Gender | Interaction | ||
| L*45 min | 47.53 b | 48.13 a | 47.85 | 47.81 | 0.10 | ** | NS | NS |
| a*45 min | 1.82 b | 2.13 a | 2.09 A | 1.87 B | 0.04 | ** | ** | NS |
| b*45 min | 3.75 | 3.99 | 3.84 | 3.82 | 0.05 | NS | NS | NS |
| pH45 min | 6.45 | 6.43 | 6.43 | 6.45 | 0.01 | NS | NS | NS |
| L*24 h | 54.43 b | 55.39 a | 55.03 | 54.81 | 0.14 | * | NS | NS |
| a*24 h | 2.19 b | 2.31 a | 2.48 A | 2.02 B | 0.06 | * | ** | NS |
| b*24 h | 6.19 | 6.10 | 6.33 | 5.97 | 0.09 | NS | NS | NS |
| pH24 h | 5.81 b | 5.99 a | 5.87 | 5.84 | 0.01 | ** | NS | NS |
| Item 4 | Treatments | Gender | SEM | Significance | ||||
|---|---|---|---|---|---|---|---|---|
| Control | MET Group | Male | Female | Treatments | Gender | Interaction | ||
| L*45 min | 52.14 | 52.19 | 52.28 | 52.05 | 0.18 | NS | NS | NS |
| a*45 min | 3.44 | 3.39 | 3.57A | 3.26B | 0.06 | NS | ** | NS |
| b*45 min | 1.63 | 1.69 | 1.56 | 1.76 | 0.07 | NS | NS | NS |
| pH45 min | 6.45 | 6.44 | 6.45 | 6.44 | 0.01 | NS | NS | NS |
| L*24 h | 54.36 | 54.73 | 54.53 | 54.55 | 0.17 | NS | NS | NS |
| a*24 h | 3.22 b | 3.51 a | 3.45 | 3.28 | 0.05 | ** | NS | NS |
| b*24 h | 2.75 b | 3.32 a | 3.01 | 3.06 | 0.07 | ** | NS | NS |
| pH24 h | 6.22 b | 6.40 a | 6.29 | 6.27 | 0.01 | ** | NS | NS |
| Items | Control | MET Group | SEM | Significance |
|---|---|---|---|---|
| Breast drip loss (%) | 3.68 a | 3.47 b | 0.12 | * |
| Thigh drip loss (%) | 3.64 a | 2.71 b | 0.11 | ** |
| Breast shear force(g) | 3028.1 a | 2543.0 b | 56.0 | ** |
| Thigh shear force(g) | 1616.5 a | 1465.0 b | 76.8 | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsharkawy, M.S.; Chen, Y.; Liu, R.; Tan, X.; Li, W.; El-Wardany, I.; Zhao, D.; Zheng, M.; Wen, J.; Zhao, G. Paternal Dietary Methionine Supplementation Improves Carcass Traits and Meat Quality of Chicken Progeny. Animals 2021, 11, 325. https://doi.org/10.3390/ani11020325
Elsharkawy MS, Chen Y, Liu R, Tan X, Li W, El-Wardany I, Zhao D, Zheng M, Wen J, Zhao G. Paternal Dietary Methionine Supplementation Improves Carcass Traits and Meat Quality of Chicken Progeny. Animals. 2021; 11(2):325. https://doi.org/10.3390/ani11020325
Chicago/Turabian StyleElsharkawy, Mohamed Shafey, Ying Chen, Ranran Liu, Xiaodong Tan, Wei Li, Ibrahim El-Wardany, Dongqin Zhao, Maiqing Zheng, Jie Wen, and Guiping Zhao. 2021. "Paternal Dietary Methionine Supplementation Improves Carcass Traits and Meat Quality of Chicken Progeny" Animals 11, no. 2: 325. https://doi.org/10.3390/ani11020325
APA StyleElsharkawy, M. S., Chen, Y., Liu, R., Tan, X., Li, W., El-Wardany, I., Zhao, D., Zheng, M., Wen, J., & Zhao, G. (2021). Paternal Dietary Methionine Supplementation Improves Carcass Traits and Meat Quality of Chicken Progeny. Animals, 11(2), 325. https://doi.org/10.3390/ani11020325