
Processing Matters in Nutrient-Matched Laboratory Diets for Mice—Energy and Nutrient Digestibility

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Animals, Materials and Methods
2.1. Animals and Diets
2.2. Digestibility Trial
2.3. Organic Acids
2.4. Post-Mortem Sampling
2.5. Statistics
3. Results
3.1. Diets
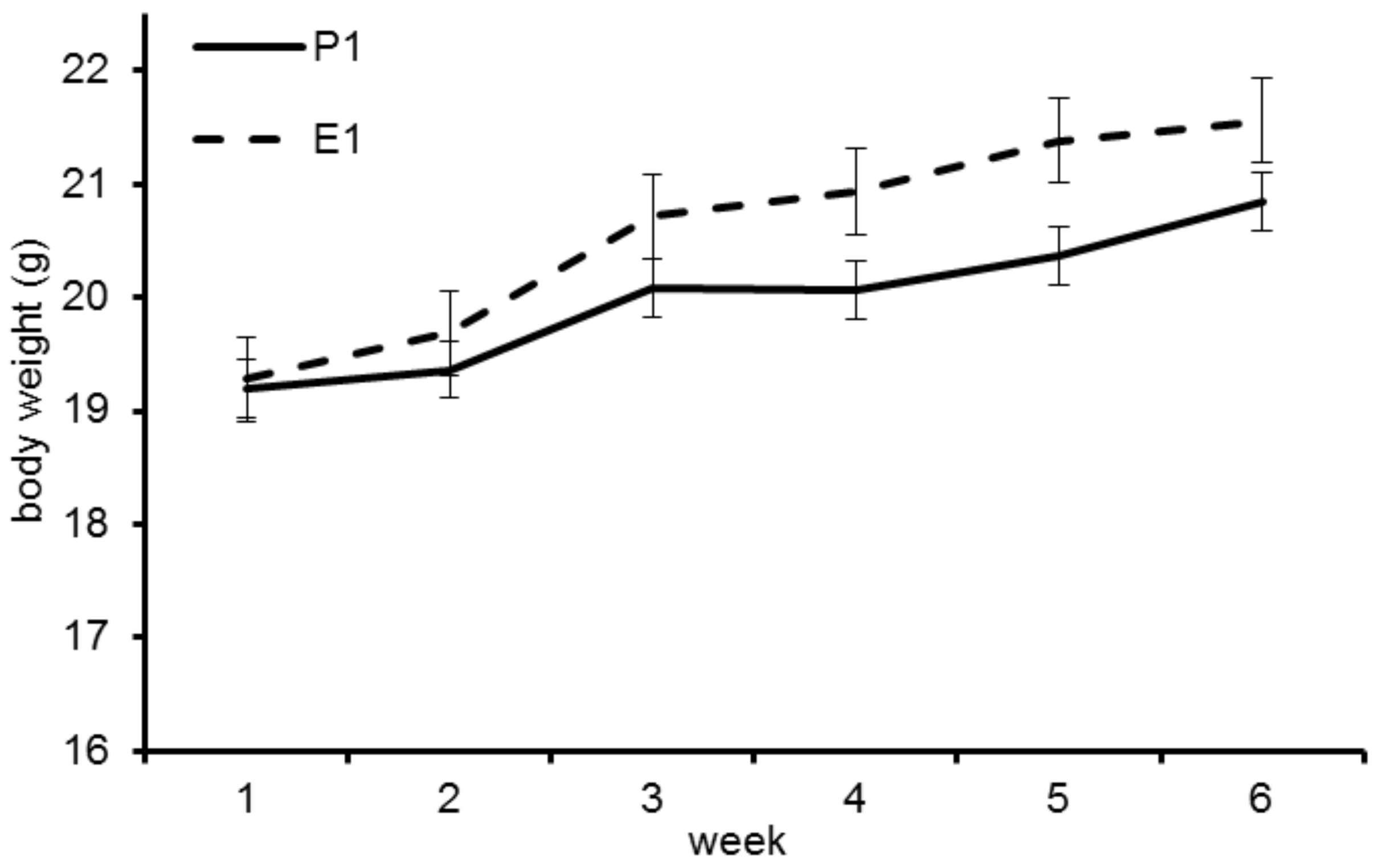
3.2. Trial 1
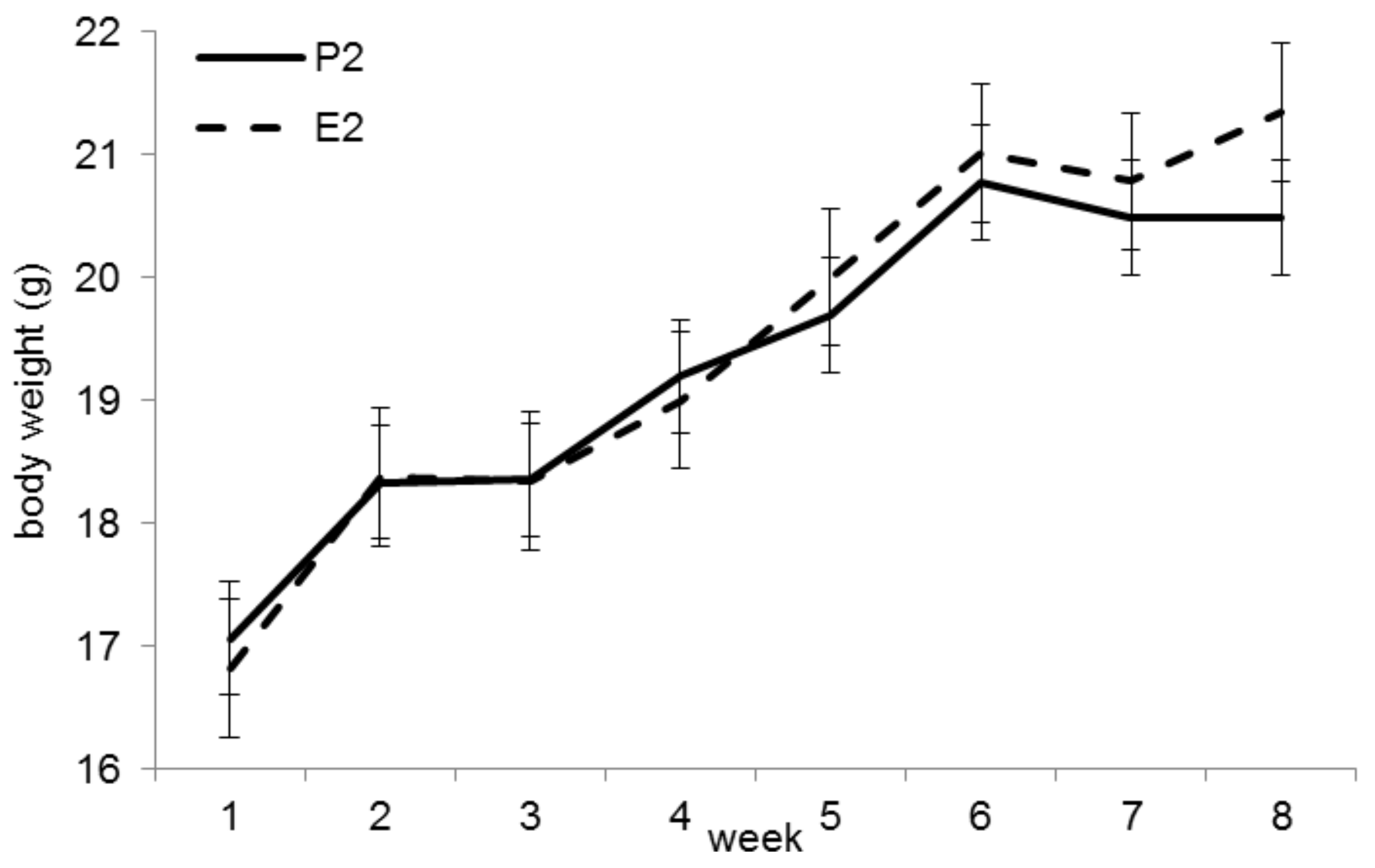
3.3. Trial 2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, L.; Combs, G.F., Jr.; DeMars, L.C.; Johnson, L.K. Effects of the physical form of the diet on food intake, growth, and body composition changes in mice. J. Am. Assoc. Anim. Sci. 2011, 50, 488–494. [Google Scholar]
- Levy, D.R.; Flores, R.; Garcia, G.E.; Craig, S.L.; Jensen, V.B. Effects of Extruded Compared with Pelleted Diets on Laboratory Mice Housed in Individually Ventilated Cages and the Cage Environment. J. Am. Assoc. Anim. Sci. 2018, 57, 686–694. [Google Scholar] [CrossRef]
- Jane, J. Starch properties, modifications, and applications. J. Macromol. Sci. Part A Pure Appl. Chem. 1995, 32, 751–757. [Google Scholar] [CrossRef]
- Zhu, L.; Jones, C.; Guo, Q.; Lewis, L.; Stark, C.R.; Alavi, S. An evaluation of total starch and starch gelatinization methodologies in pelleted animal feed. J. Anim. Sci. 2016, 94, 1501–1507. [Google Scholar] [CrossRef] [Green Version]
- Case, S.; Hamann, D.; Schwartz, S. Effect of starch gelatinization on physical properties of extrused wheat-and corn-based products. Cereal Chem. 1992, 69, 401–404. [Google Scholar]
- Zimonja, O.; Svihus, B. Effects of processing of wheat or oats starch on physical pellet quality and nutritional value for broilers. Anim. Feed Sci. Technol. 2009, 149, 287–297. [Google Scholar] [CrossRef]
- Tran, Q.; van Lin, C.; Hendriks, W.; van der Poel, A. Lysine reactivity and starch gelatinization in extruded and pelleted canine diets. Anim. Feed Sci. Technol. 2007, 138, 162–168. [Google Scholar] [CrossRef]
- Böswald, L.; Kienzle, E. Differences in starch gelatinisation between differently processed laboratory rodent maintenance diets labelled as identical. In Proceedings of the 73rd Society of Nutrition Physiology (GfE) Conference, Goettingen, Germany, 13–15 March 2019. [Google Scholar]
- Bergman, E. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.M.; Jenkins, D.J. Carbohydrate digestibility and metabolic effects. J. Nutr. 2007, 137, 2539S–2546S. [Google Scholar] [CrossRef]
- Houdijk, J.; Hartemink, R.; Verstegen, M.; Bosch, M. Effects of dietary non-digestible oligosaccharides on microbial characteristics of ileal chyme and faeces in weaner pigs. Arch. Anim. Nutr. 2002, 56, 297–307. [Google Scholar] [CrossRef]
- Zentek, J. Influence of diet composition on the microbial activity in the gastro-intestinal tract of dogs. I. Effects of varying protein intake on the composition of the ileum chyme and the faeces. J. Anim. Physiol. Anim. Nutr. 1995, 74, 43–52. [Google Scholar] [CrossRef]
- Drochner, W.; Kerler, A.; Zacharias, B. Pectin in pig nutrition, a comparative review. J. Anim. Physiol. Anim. Nutr. 2004, 88, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Kienzle, E.; Ahlborn, H.H.; Meyer, H.; Ganter, M. Effect of lactose on apparent prececal and fecal digestibility of crude nutrients and minerals in minipig. J. Anim. Physiol. Anim. Nutr. 1995, 74, 227–234. [Google Scholar] [CrossRef]
- Jiang, F.; Du, C.; Jiang, W.; Wang, L.; Du, S.-K. The preparation, formation, fermentability, and applications of resistant starch. Int. J. Biol. Macromol. 2020, 150, 1155–1161. [Google Scholar] [CrossRef]
- Holm, J.; Lundquist, I.; Björck, I.; Eliasson, A.-C.; Asp, N.-G. Degree of starch gelatinization, digestion rate of starch in vitro, and metabolic response in rats. Am. J. Clin. Nutr. 1988, 47, 1010–1016. [Google Scholar] [CrossRef]
- Naumann, C.; Bassler, R. Die Chemische Untersuchung von Futtermitteln. Band III: Methodenbuch; Verlag J. Naumann: Neudamm, Germany, 1986. [Google Scholar]
- FEDIAF. Nutritional Guidelines for Complete and Complementary Pet Food for Cats and Dogs; Fediaf: Brussels, Belgium, 2020. [Google Scholar]
- GfE. Opinion on the indispensability of animal experiments in animal nutrition research and suitability of alternative methods. Proc. Soc. Nutr. Physiol. 2017, 26, 218–224. [Google Scholar]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen & Co: London, UK, 1959. [Google Scholar]
- Hald, S.; Schioldan, A.G.; Moore, M.E.; Dige, A.; Lærke, H.N.; Agnholt, J.; Bach Knudsen, K.E.; Hermansen, K.; Marco, M.L.; Gregersen, S. Effects of arabinoxylan and resistant starch on intestinal microbiota and short-chain fatty acids in subjects with metabolic syndrome: A randomised crossover study. PLoS ONE 2016, 11, e0159223. [Google Scholar] [CrossRef] [PubMed]
- Haidar, M.N.; Petie, M.; Heinsbroek, L.T.; Verreth, J.A.; Schrama, J.W. The effect of type of carbohydrate (starch vs. nonstarch polysaccharides) on nutrients digestibility, energy retention and maintenance requirements in Nile tilapia. Aquaculture 2016, 463, 241–247. [Google Scholar] [CrossRef]
- Högberg, A.; Lindberg, J.E. Influence of cereal non-starch polysaccharides and enzyme supplementation on digestion site and gut environment in weaned piglets. Anim. Feed Sci. Technol. 2004, 116, 113–128. [Google Scholar] [CrossRef]
- Sasaki, T.; Kohyama, K. Influence of non-starch polysaccharides on the in vitro digestibility and viscosity of starch suspensions. Food Chem. 2012, 133, 1420–1426. [Google Scholar] [CrossRef]
- Spiller, G.A.; Chernoff, M.C.; Hill, R.A.; Gates, J.E.; Nassar, J.J.; Shipley, E.A. Effect of purified cellulose, pectin, and a low-residue diet on fecal volatile fatty acids, transit time, and fecal weight in humans. Am. J. Clin. Nutr. 1980, 33, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Bjornvad, C.R.; Nielsen, D.H.; Armstrong, P.J.; McEvoy, F.; Hoelmkjaer, K.M.; Jensen, K.S.; Pedersen, G.F.; Kristensen, A.T. Evaluation of a nine-point body condition scoring system in physically inactive pet cats. Am. J. Vet. Res. 2011, 72, 433–437. [Google Scholar] [CrossRef]
- Foulis, S.A.; Hughes, J.M.; Friedl, K.E. New concerns about military recruits with metabolic obesity but normal weight (“skinny fat”). Obesity 2020, 28, 223. [Google Scholar] [CrossRef]
- Stewart, R.; Preece, R.; Sheppard, H.G. Twelve generations of marginal protein deficiency. Br. J. Nutr. 1975, 33, 233–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Leu, R.K.; Brown, I.L.; Hu, Y.; Morita, T.; Esterman, A.; Young, G.P. Effect of dietary resistant starch and protein on colonic fermentation and intestinal tumourigenesis in rats. Carcinogenesis 2007, 28, 240–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Diet Parameter | Labeled Content | Trial 1 | Trial 2 | ||
|---|---|---|---|---|---|
| P1 | E1 | P2 | E2 | ||
| Gross energy (MJ/kg) | - | 17.23 | 17.56 | 16.6 | 17.3 |
| Dry matter (%) | 88.8 | 89.8 | 89.8 | 89.5 | 88.5 |
| Crude protein (%) | 20.6 | 23.4 * | 19.5 | 18.7 | 22.5 |
| Crude fat (%) | 4.1 | 3.6 | 4.3 | 2.5 * | 5.1 * |
| Crude ash (%) | 5.9 | 5.8 | 5.6 | 4.8 | 6.1 |
| Crude fiber (%) | 6.1 | 6.1 | 8.2 * | 5.2 | 5.8 |
| Nitrogen-free extracts (%, calculated) | 55.0 | 50.9 | 52.2 | 58.3 | 49.0 |
| Starch (%) | - | 27 | 27 | 43 | 28 |
| Starch gelatinization (%) | |||||
| - before autoclaving | - | 22 | 64 | 22 | 50 |
| - after autoclaving | 15 | 57 | 17 | 70 | |
| Total dietary fiber (%) | - | 27.8 | 24.5 | 15.9 | 23.3 |
| Soluble dietary fiber (%) | - | 3.4 | 4.0 | 2.0 | 3.9 |
| Insoluble dietary fiber (%) | - | 24.4 | 20.5 | 13.9 | 19.4 |
| Sugars (%) | - | 6.2 | 6.1 | 4.6 | 6.5 |
| aD [%] | Trial 1 | Trial 2 | ||||
|---|---|---|---|---|---|---|
| Diet P1 n = 11 | Diet E1 n = 11 | p | Diet P2 n = 16 | Diet E2 n = 16 | p | |
| GE | 74.8 ± 0.5 | 80.9 ± 0.6 | <0.001 | 84.4 ± 0.3 | 81.9 ± 0.6 | <0.001 |
| DM | 70.8 ± 0.8 | 77.2 ± 0.7 | <0.001 | 81.3 ± 0.2 | 77.6 ± 0.8 | <0.001 |
| OM | 73.7 ± 0.6 | 80.3 ± 0.5 | <0.001 | 84.7 ± 0.2 | 80.8 ± 0.7 | <0.001 |
| CP | 82.2 ± 0.5 | 81.7 ± 0.9 | 0.31 | 83.8 ± 0.3 | 85.1 ± 0.4 | <0.001 |
| EE | 91.2 ± 0.7 | 92.0 ± 0.5 | 0.09 | 92.9 ± 1.4 | 93.4 ± 1.0 | 0.71 |
| CH+F | 69.2 ± 0.7 | 79.1 ± 0.4 | <0.001 | 85.1 ± 1.7 | 81.7 ± 4.5 | <0.05 |
| Sample | Trial 1 | Trial 2 | ||||
|---|---|---|---|---|---|---|
| Diet P1 n = 11 | Diet E1 n = 11 | p | Diet P2 n = 16 | Diet E2 n = 16 | p | |
| stomach | 3.9 ± 0.5 | 3.7 ± 0.7 | 0.46 | 3.9 ± 0.9 | 3.6 ± 0.5 | 0.33 |
| anterior small intestine | 6.7 ± 0.3 | 6.6 ± 0.3 | 0.54 | 6.7 ± 0.5 | 6.7 ± 0.3 | 0.76 |
| posterior small intestine | 7.0 ± 0.4 | 7.0 ± 0.3 | 0.70 | 7.3 ± 0.3 | 7.2 ± 0.2 | 0.11 |
| cecum | 6.5 ± 1.1 | 6.9 ± 0.4 | 0.24 | 7.2 ± 0.3 | 7.1 ± 0.2 | 0.16 |
| colon | 6.9 ± 0.3 | 7.2 ± 0.1 | <0.05 | 7.6 ± 0.3 | 7.5 ± 0.3 | 0.14 |
| faeces | 7.6 ± 0.1 | 7.4 ± 0.1 | <0.05 | 7.9 ± 0.2 | 7.7 ± 0.4 | 0.33 |
| mmol/L | Trial 1 | Trial 2 | ||||
|---|---|---|---|---|---|---|
| Diet P1 n = 11 | Diet E1 n = 11 | p | Diet P2 n = 16 | Diet E2 n = 16 | p | |
| acetic acid | 8.37 ± 1.52 | 6.95 ± 1.61 | 0.09 | 6.44 ± 1.16 | 6.73 ± 0.97 | 0.60 |
| propionic acid | 1.16 ± 0.15 | 1.17 ± 0.15 | 0.92 | 1.19 ± 0.17 | 1.25 ± 0.18 | 0.47 |
| n-butyric acid | 1.18 ± 0.21 | 1.09 ± 0.02 | 0.57 | 0.47 ± 0.08 | 0.55 ± 0.08 | 0.06 |
| acetic acid/propionic acid | 7.15 / 1 | 5.92/1 | 5.41/1 | 5.38/1 | ||
| Parameter | Diet P2 n = 16 | Diet E2 n = 16 | p |
|---|---|---|---|
| Gross energy (MJ/kg DM) | 25.1 ± 1.9 | 23.7 ± 1.2 | <0.05 |
| Crude protein (% DM) | 46.1 ± 7.3 | 51.9 ± 4.7 | <0.05 |
| Fat (% DM) | 35.4 ± 9.6 | 28.4 ± 5.9 | <0.05 |
| Ash (% DM) | 10.9 ± 1.6 | 12.1 ± 1.3 | <0.05 |
| Liver energy content (MJ/kg) | 7.1 ± 0.3 | 6.9 ± 0.2 | 0.26 |
| Liver weight in % final BW | 4.7 ± 0.6 | 5.2 ± 0.3 | <0.01 |
| GIT weight in % final BW | 13.1 ± 1.6 | 13.8 ± 3.1 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Böswald, L.F.; Wenderlein, J.; Straubinger, R.K.; Ulrich, S.; Kienzle, E. Processing Matters in Nutrient-Matched Laboratory Diets for Mice—Energy and Nutrient Digestibility. Animals 2021, 11, 523. https://doi.org/10.3390/ani11020523
Böswald LF, Wenderlein J, Straubinger RK, Ulrich S, Kienzle E. Processing Matters in Nutrient-Matched Laboratory Diets for Mice—Energy and Nutrient Digestibility. Animals. 2021; 11(2):523. https://doi.org/10.3390/ani11020523
Chicago/Turabian StyleBöswald, Linda F., Jasmin Wenderlein, Reinhard K. Straubinger, Sebastian Ulrich, and Ellen Kienzle. 2021. "Processing Matters in Nutrient-Matched Laboratory Diets for Mice—Energy and Nutrient Digestibility" Animals 11, no. 2: 523. https://doi.org/10.3390/ani11020523
APA StyleBöswald, L. F., Wenderlein, J., Straubinger, R. K., Ulrich, S., & Kienzle, E. (2021). Processing Matters in Nutrient-Matched Laboratory Diets for Mice—Energy and Nutrient Digestibility. Animals, 11(2), 523. https://doi.org/10.3390/ani11020523