
A De Novo Mutation in COL1A1 in a Holstein Calf with Osteogenesis Imperfecta Type II

 ,
,  , and
, and 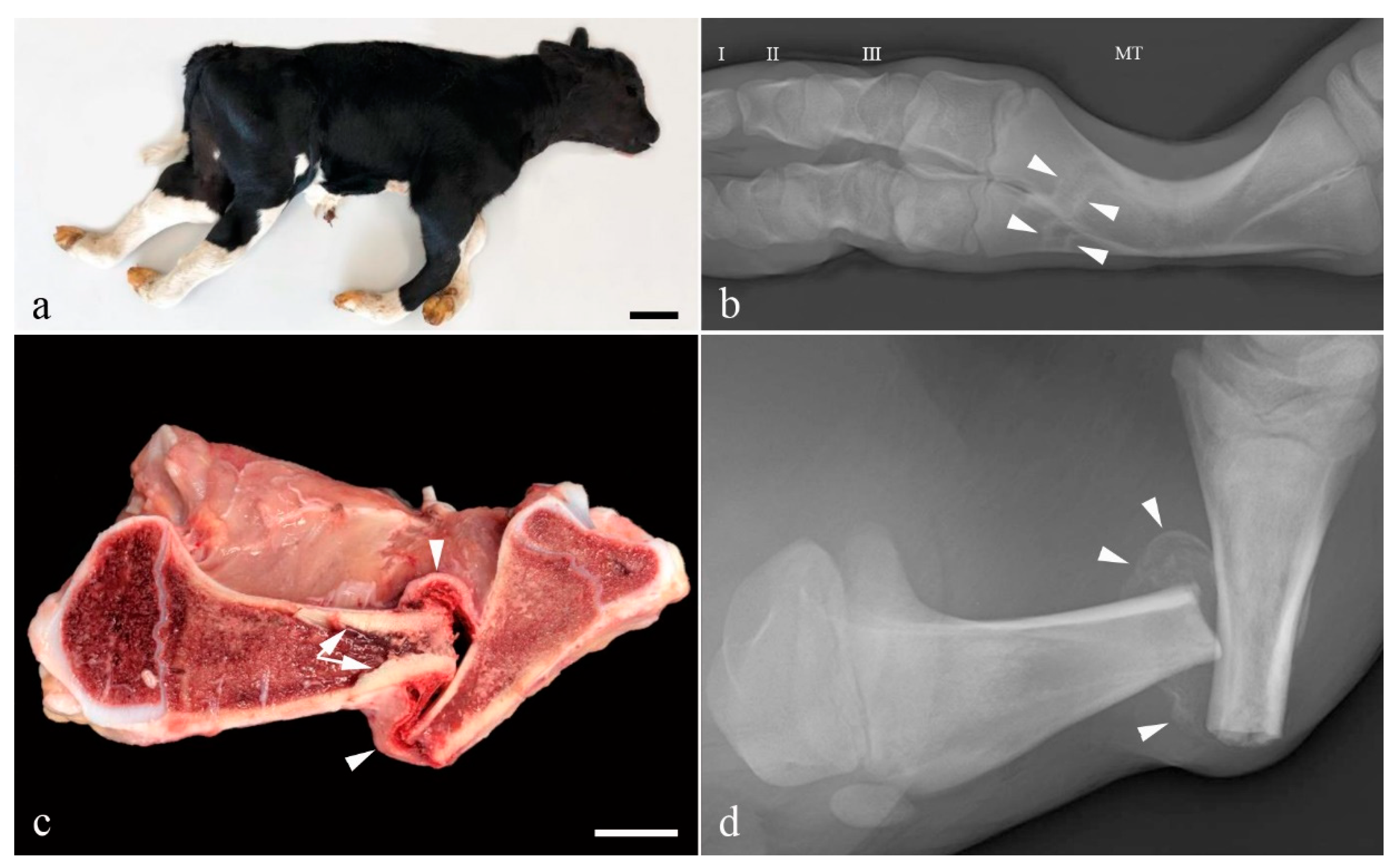{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pathological Investigation
2.2. DNA Samples
2.3. Whole-Genome Sequencing
2.4. Targeted Genotyping
2.5. Evaluation of the Molecular Consequences of Amino Acid Substitutions
2.6. Sequence Accessions
3. Results
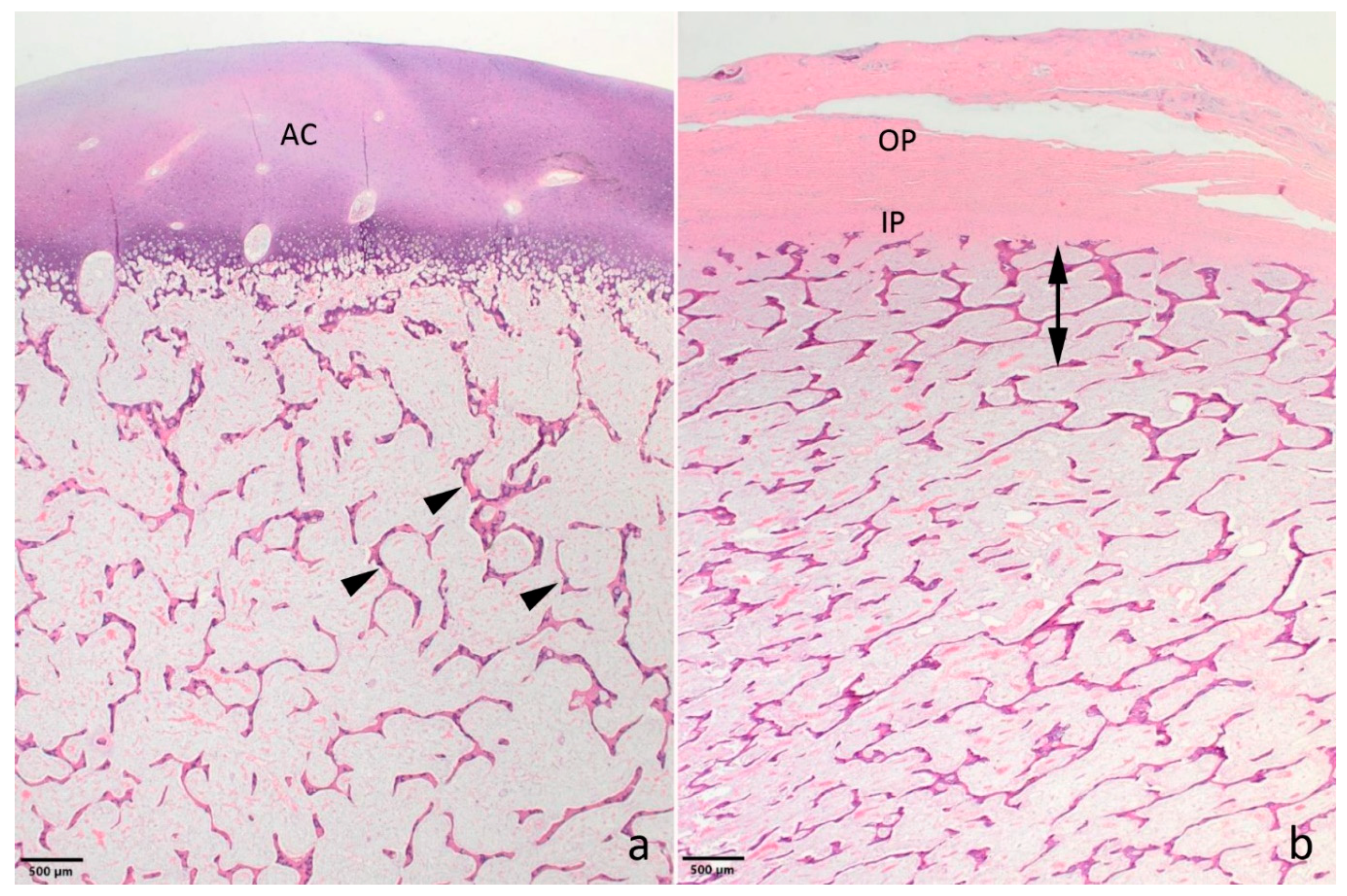
3.1. Pathological Phenotype
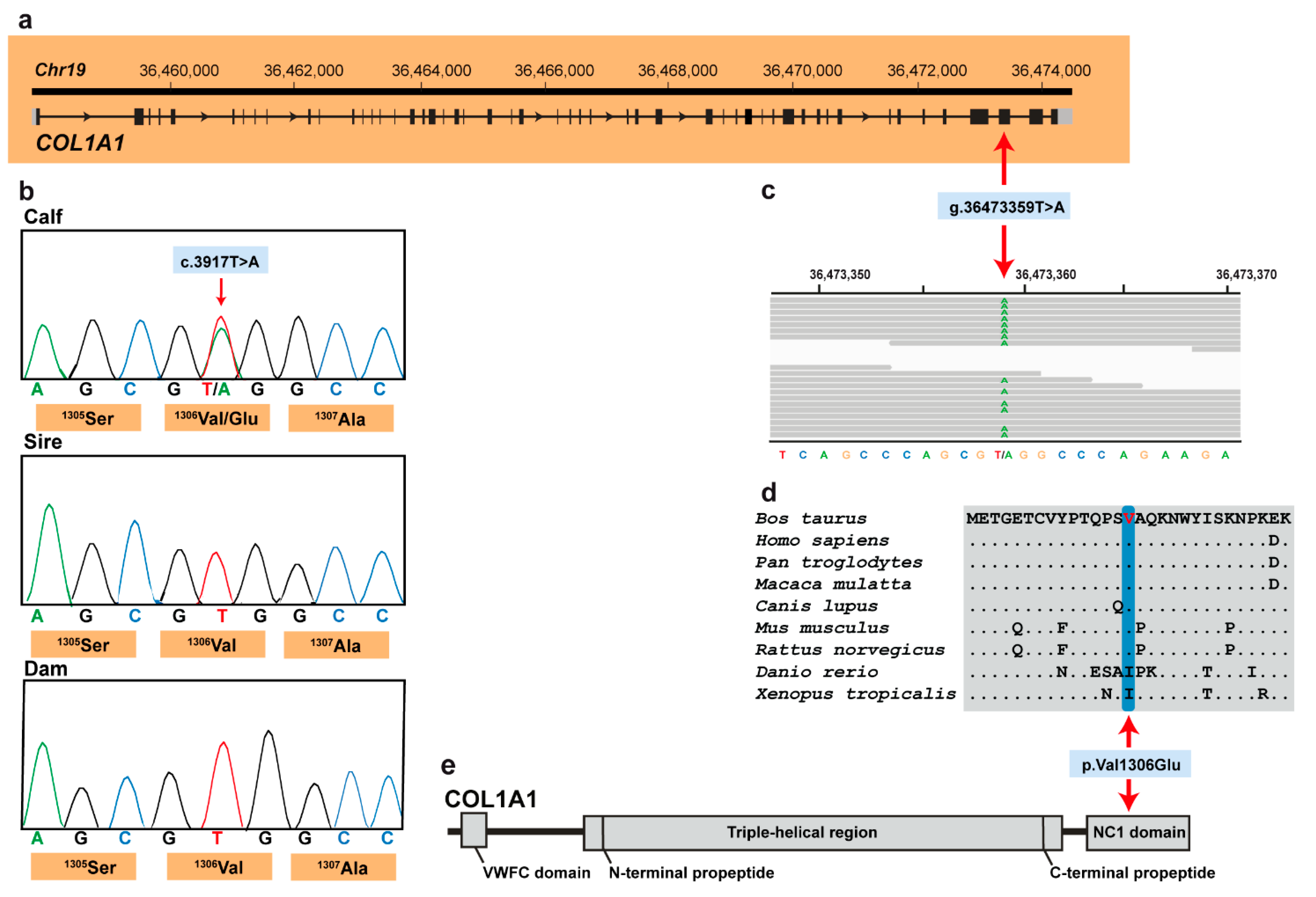
3.2. Genetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marini, J.C.; Forlino, A.; Cabral, W.A.; Barnes, A.M.; Antonio, J.D.S.; Milgrom, S.; Hyland, J.C.; Körkkö, J.; Prockop, D.J.; De Paepe, A.; et al. Consortium for osteogenesis imperfecta mutations in the helical domain of type I collagen: Regions rich in lethal mutations align with collagen binding sites for integrins and proteoglycans. Hum. Mutat. 2007, 28, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Rossi, V.; Lee, B.; Marom, R. Osteogenesis imperfecta: Advancements in genetics and treatment. Curr. Opin. Pediatr. 2019, 31, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Peddada, K.V.; Sullivan, B.T.; Margalit, A.; Sponseller, P.D. Fracture Patterns Differ Between Osteogenesis Imperfecta and Routine Pediatric Fractures. J. Pediatr. Orthop. 2018, 38, e207–e212. [Google Scholar] [CrossRef] [PubMed]
- Marini, J.C.; Cabral, W.A. Osteogenesis Imperfecta Genetics of Bone Biology and Skeletal Disease; Elsevier Inc: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Kater, J.C.; Hartley, W.; Dysart, T.; Campbell, A. Osteogenesis imperfecta and bone resorption: Two unusual skeletal abnormalities in young lambs. New Zealand Veter- J. 1963, 11, 41–46. [Google Scholar] [CrossRef]
- Cohn, L.; Meuten, D.J. Bone fragility in a kitten: An osteogenesis imperfecta-like syndrome. J. Am. Veter- Med. Assoc. 1990, 197, 98–100. [Google Scholar]
- Calkins, E.; Kalm, D.; Diner, W.C. Idiopathic familial osteoporosis in dogs: “Osteogenesis imperfecta”. Ann. N. Y. Acad. Sci. 1956, 64, 410–423. [Google Scholar] [CrossRef]
- Jensen, P.T.; Rusmussen, P.G.; Basse, A. Congenital osteogenesis imperfecta in Charolais cattle. Nord. Vet. Med. 1976, 28, 304–308. [Google Scholar]
- Campbell, B.G.; Wootton, J.A.; MacLeod, J.N.; Minor, R.R. Sequence of Normal Canine COL1A1 cDNA and Identification of a Heterozygous α1(I) Collagen Gly208AIa Mutation in a Severe Case of Canine Osteogenesis Imperfecta. Arch. Biochem. Biophys. 2000, 384, 37–46. [Google Scholar] [CrossRef]
- Petersen, J.L.; Tietze, S.M.; Burrack, R.M.; Steffen, D.J. Evidence for a de novo, dominant germ-line mutation causative of osteogenesis imperfecta in two Red Angus calves. Mamm. Genome 2019, 30, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Bourneuf, E.; Otz, P.; Pausch, H.; Jagannathan, V.; Michot, P.; Grohs, C.; Piton, G.; Ammermüller, S.; Deloche, M.C.; Fritz, S.; et al. Rapid Discovery of De Novo Deleterious Mutations in Cattle Enhances the Value of Livestock as Model Species. Sci. Rep. 2017, 7, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letko, A.; Zdora, I.; Hitzler, V.; Jagannathan, V.; Beineke, A.; Möhrke, C.; Drögemüller, C. A de novo in-frame duplication in the COL 1A2 gene in a Lagotto Romagnolo dog with osteogenesis imperfecta. Anim. Genet. 2019, 50, 786–787. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.G.; Wootton, J.A.M.; MacLeod, J.N.; Minor, R.R. Canine COL1A2 Mutation Resulting in C-Terminal Truncation of Pro-α2(I) and Severe Osteogenesis Imperfecta. J. Bone Miner. Res. 2001, 16, 1147–1153. [Google Scholar] [CrossRef] [PubMed]
- Quist, E.M.; Doan, R.; Pool, R.R.; Porter, B.F.; Bannasch, D.L.; Dindot, S.V. Identification of a Candidate Mutation in the COL1A2 Gene of a Chow Chow With Osteogenesis Imperfecta. J. Hered. 2017, 109, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Drögemüller, C.; Becker, D.; Brunner, A.; Haase, B.; Kircher, P.; Seeliger, F.; Fehr, M.; Baumann, U.; Lindblad-Toh, K.; Leeb, T. A Missense Mutation in the SERPINH1 Gene in Dachshunds with Osteogenesis Imperfecta. PLoS Genet. 2009, 5, e1000579. [Google Scholar] [CrossRef] [Green Version]
- Rosen, B.D.; Bickhart, D.M.; Schnabel, R.D.; Koren, S.; Elsik, C.G.; Tseng, E.; Rowan, T.N.; Low, W.Y.; Zimin, A.; Couldrey, C.; et al. De novo assembly of the cattle reference genome with single-molecule sequencing. GigaScience 2020, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hayes, B.J.; Daetwyler, H.D. 1000 Bull Genomes Project to Map Simple and Complex Genetic Traits in Cattle: Applications and Outcomes. Annu. Rev. Anim. Biosci. 2019, 7, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinform. 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Häfliger, I.M.; Wiedemar, N.; Švara, T.; Starič, J.; Cociancich, V.; Šest, K.; Gombač, M.; Paller, T.; Agerholm, J.S.; Drögemüller, C. Identification of small and large genomic candidate variants in bovine pulmonary hypoplasia and anasarca syndrome. Anim. Genet. 2020, 51, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Wenger, A.M.; Zehir, A.; Mesirov, J.P. Variant Review with the Integrative Genomics Viewer. Cancer Res. 2017, 77, e31–e34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinform. 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.H.M.; Pires, D.E.V.; Ascher, D.B. DynaMut: Predicting the impact of mutations on protein conformation, flexibility and stability. Nucleic Acids Res. 2018, 46, W350–W355. [Google Scholar] [CrossRef]
- Lamandé, S.R.; Bateman, J.F. Genetic Disorders of the Extracellular Matrix. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2019, 303, 1527–1542. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Shi, L.; Wang, W.; Zuo, R.; Wang, M.; Kang, Q. Identification of two novel COL10A1 heterozygous mutations in two Chinese pedigrees with Schmid-type metaphyseal chondrodysplasia. BMC Med. Genet. 2019, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.; Cole, W.G.; Rogers, J.; Bateman, J.F. A mutation in the conserved NC1 domain of type X collagen prevents in vitro multimer assembly resulting in a Schmid-type metaphyseal chondrodysplasia. Matrix Biol. 1994, 14, 396–397. [Google Scholar] [CrossRef]
- Chan, D.; Weng, Y.M.; Golub, S.; Bateman, J.F. Type X collagen NC1 mutations produced by site-directed mutagenesis prevent in vitro assembly. Ann. N. Y. Acad. Sci. 1996, 785, 231–233. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Freddi, S.; Bateman, J.F. Collagen X Chains Harboring Schmid Metaphyseal Chondrodysplasia NC1 Domain Mutations Are Selectively Retained and Degraded in Stably Transfected Cells. J. Biol. Chem. 2002, 277, 12516–12524. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacinto, J.G.P.; Häfliger, I.M.; McEvoy, F.J.; Drögemüller, C.; Agerholm, J.S. A De Novo Mutation in COL1A1 in a Holstein Calf with Osteogenesis Imperfecta Type II. Animals 2021, 11, 561. https://doi.org/10.3390/ani11020561
Jacinto JGP, Häfliger IM, McEvoy FJ, Drögemüller C, Agerholm JS. A De Novo Mutation in COL1A1 in a Holstein Calf with Osteogenesis Imperfecta Type II. Animals. 2021; 11(2):561. https://doi.org/10.3390/ani11020561
Chicago/Turabian StyleJacinto, Joana G. P., Irene M. Häfliger, Fintan J. McEvoy, Cord Drögemüller, and Jørgen S. Agerholm. 2021. "A De Novo Mutation in COL1A1 in a Holstein Calf with Osteogenesis Imperfecta Type II" Animals 11, no. 2: 561. https://doi.org/10.3390/ani11020561
APA StyleJacinto, J. G. P., Häfliger, I. M., McEvoy, F. J., Drögemüller, C., & Agerholm, J. S. (2021). A De Novo Mutation in COL1A1 in a Holstein Calf with Osteogenesis Imperfecta Type II. Animals, 11(2), 561. https://doi.org/10.3390/ani11020561