
The Identification of Mutation in BMP15 Gene Associated with Litter Size in Xinjiang Cele Black Sheep

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. DNA Extraction
2.3. Detection of FecXI, FecXH, FecXG, and FecXB Mutation Sites of the BMP15 Gene
2.4. Exons 1 and 2 of BMP15 Gene Sequence Analysis
2.5. Mutation Detection Assays
2.6. Statistical Analysis
3. Results
3.1. Detection of FecXI, FecXH, FecXG, and FecXB Mutation Sites of the BMP15 Gene
3.2. Sequencing Results of the BMP15 Gene Coding Region
3.2.1. Exon 1 of the BMP15 Gene Sequence
3.2.2. Exon 2 of the BMP15 Gene Sequence
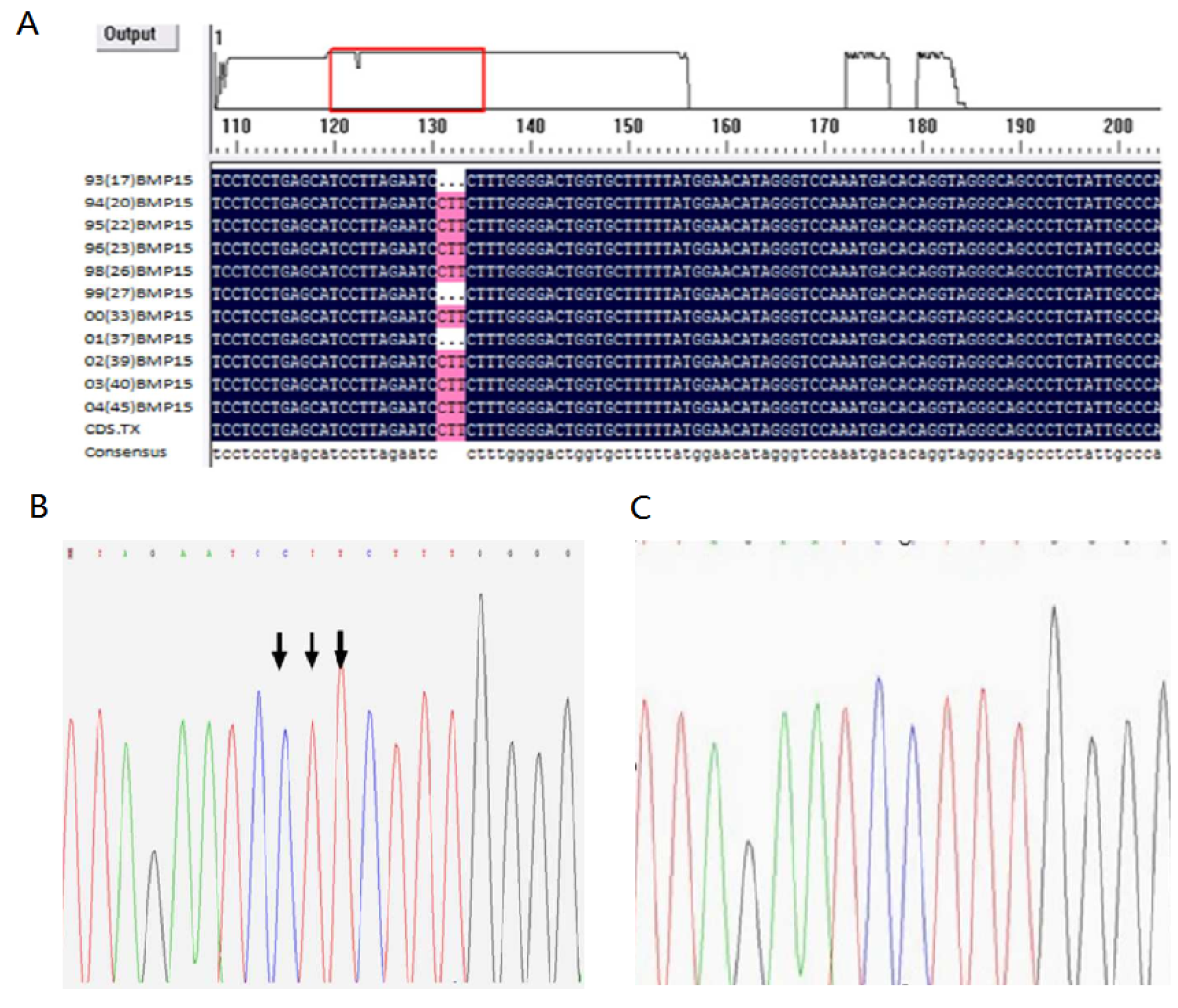
3.3. Mutation of the BMP15 Gene Polymorphism
3.4. Genotypic and Allelic Frequencies of the Mutation of the BMP15 Gene in Cele Black Sheep
3.5. Effect of Genotypes on Litter Size
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdoli, R.; Mirhoseini, S.Z.; Ghavi Hossein-Zadeh, N.; Zamani, P.; Gondro, C. Genome-wide association study to identify genomic regions affecting prolificacy in Lori-Bakhtiari sheep. Anim. Genet. 2018, 49, 488–491. [Google Scholar] [CrossRef]
- Narjess, L.; Zohra, B.; Florent, W.; Ahmed, R.; Mohamed, A.; Mourad, R.; Stephane, F.; Sonia, B.R. FecX a Novel BMP15 mutation responsible for prolificacy and female sterility in Tunisian Barbarine Sheep. BMC Genet. 2017, 18, 43. [Google Scholar]
- Morteza, M.; Shahram, N.; Davood, S.H. Mutation in BMPR-IB gene is associated with litter size in Iranian Kalehkoohi sheep. Anim. Reprod. Sci. 2014, 147, 93–98. [Google Scholar]
- McNatty, K.P.; Juengell, J.L.; Wilson, T.; Galloway, S.M.; Davis, G.H.; Hudson, N.L.; Moeller, C.L.; Cranfield, M.; Reade, K.L.; Laitinen, M.P.; et al. Oocyte-derived growth factors and ovulation rate in sheep. Reproduction 2003, 61, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Davis, G.H. Major genes affecting ovulation rate in sheep. Genet. Sel. Evol. 2005, 37 (Suppl. 1), S11–S23. [Google Scholar] [CrossRef] [Green Version]
- Hiam, N.; Karima, G.M.M.; Mohamed, M.M.K.; Nermeen, A.H.; Shawky, S.I.; Mahmoud, F.N.; Othman, E. PCR-RFLP of bone morphogenetic protein 15 (BMP15/FecX) gene as a candidate for prolificacy in sheep. Int. J. Vet. Sci. Med. 2018, 6, S68–S72. [Google Scholar]
- Shi, H.; Bai, J.; Niu, Z.; Esha, M.; Fen, L.; Jia, B. Study on Candidate Gene for Fecundity Traits in Xingjiang Cele Black Sheep. Afr. J. Biotechnol. 2010, 9, 8498–8505. [Google Scholar]
- Galloway, S.M.; McNatty, K.P.; Cambridge, L.M.; Laitinen, M.P.E.; Juengel, J.L.; Jokiranta, T.S.; McLaren, R.J.; Luiro, K.; Dodds, K.G.; Montgomery, G.W.; et al. Mutations in an oocyte-derived growth factor gene (BMP15) cause increased ovulation rate and infertility in a dosage-sensitive manner. Nat. Genet. 2000, 25, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.H.; Mcewan, J.C.; Fennessy, P.; Dodds, K.G. Discovery of the Inverdale gene (FecX). In Proceedings of the New Zealand Society of Animal Production conference, Mosgiel, New Zealand, 27–30 August 1995. [Google Scholar]
- Davis, G.H.; Bruce, G.D.; Dodds, K.G. Ovulation rate and litter size of prolific Inverdale (FecX I) and Hanna (FecX H) sheep. Proc. Assoc. Adv. Anim. Breed Genet. 2001, 14, 175–178. [Google Scholar]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef]
- Bodin, L.; Di-Pasquale, E.; Fabre, S.; Bontoux, M.; Monget, P.; Persani, L.; Mulsant, P. A novel mutation in the bone morphogenetic protein 15 gene causing defective protein secretion is associated with both increased ovulation rate and sterility in Lacaune sheep. J. Endocrinol. 2007, 148, 393–400. [Google Scholar] [CrossRef]
- Martinez-Royo, A.; Jurado, J.J.; Smulders, J.P.; Marti, J.I.; Alabart, J.L.; Roche, A.; Fantova, E.; Bodin, L.; Mulsant, P.; Serrano, M.; et al. A deletion in the bone morphogenetic protein 15 gene causes sterility and increased prolificacy in Rasa Aragonesa sheep. Anim. Genet. 2008, 39, 294–297. [Google Scholar] [CrossRef]
- Monteagudo, L.V.; Ponz, R.; Tejedor, M.T.; Lavina, A.; Sierra, I. A 17 bp deletion in the Bone Morphogenetic Protein 15 (BMP15) gene is associated to increased prolificacy in the Rasa Aragonesa sheep breed. Anim. Reprod. Sci. 2009, 110, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Amini, H.R.; Ajaki, A.; Farahi, M.; Heidari, M.; Pirali, A.; Forouzanfar, M.; Eghbalsaied, S. The novel t755c mutation in BMP15 is associated with the litter size of iranian afshari, ghezel, and shal breeds. Arch. Anim. Breed. 2013, 61, 153–160. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Purification of nucleic acids by extraction with phenol:chloroform. Cold Spring Harb. Protoc. 2006, 2006. [Google Scholar] [CrossRef] [PubMed]
- Dube, J.L.; Wang, P.; Elvin, J.; Lyons, K.M.; Celeste, A.J.; Matzuk, M.M. The Bone Morphogenetic Protein 15 Gene Is X-Linked and ExPressed in Oocytes. Mol. Endocrinol. 1998, 12, 1809–1817. [Google Scholar] [CrossRef]
- Juengel, J.L.; Hudson, N.L.; Heath, D.A.; Smith, P.; Reader, K.L.; Lawrence, S.B.; O’Connell, A.R.; Laitinen, M.P.; Cranfield, M.; Groome, N.P.; et al. Growth differentiation factor 9 and bone morphogenetic protein 15 are essential for ovarian follicular development in sheep. Biol. Reprod. 2002, 67, 1777–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juengel, J.L.; Hudson, N.L.; Berg, M.; Hamel, K.; Smith, P.; Lawrence, S.B.; Whiting, L.; McNatty, K.P. Effects of active immunization against growth differentiation factor 9 and/or bone morphogenetic protein 15 on ovarian function in cattle. Reproduction 2009, 138, 107–114. [Google Scholar] [CrossRef] [Green Version]
- McNatty, K.P.; Hudson, N.L.; Whiting, L.; Reader, K.L.; Lun, S.; Western, A.; Heath, D.A.; Smith, P.; Moore, L.G.; Juengel, J.L. The effects of immunizing sheep with different BMP15 or GDF9 peptide sequences on ovarian follicular activity and ovulation rate. Biol. Reprod. 2007, 76, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Juengel, J.L.; Hudson, N.L.; Whiting, L.; McNatty, K.P. Effects of immunization against bone morphogenetic protein 15 and growth differentiation factor 9 on ovulation rate, fertilization, and pregnancy in ewes. Biol. Reprod. 2004, 70, 557–561. [Google Scholar] [CrossRef]
- Shimasaki, S.; Moore, R.K.; Otsuka, F.; Erickson, G.F. The bone morphogenetic protein system in mammalian reproduction. Endocr. Rev. 2004, 25, 72–101. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Primer Sequence | Size |
|---|---|---|
| FecXG | Forward 5′-CACTGTCTTCTTGTTACTGTATTTCAATGAGAC-3′ Reverse 5′-GATGCAATACTGCCTGCTTG-3′ | 141 bp |
| FecXB | Forward 5′-GCCTTCCTGTGTCCCTTATAAGTATGTTCCCCTTA-3′ Reverse 5′-TACTTTCAGGCCCATCATGCTCC-3′ | 153 bp |
| FecXI | Forward 5′-GAAAGTAACCAGTGTTCCCTCCACCCTTTTCT-3′ Reverse 5′-CATGATTGGGAGAATTGAGACC-3′ | 150 bp |
| FecXH | Forward 5′-TATTTCAATGACACTCAGAG-3′ Reverse 5′-GAGCAATGATCCAGTGATCCCA-3′ | 240 bp |
| Site | Primer Sequence | Size |
|---|---|---|
| E1+28-30 | Forward 5′-TGTTACCCATGTAAAAGGAAAGG-3′ Reverse 5′-AAAAGCACCAGTCCCCAAAGAAG-3′ Forward 5′-TGTTACCCATGTAAAAGGAAAGG-3′ Reverse 5′-ACCGTAAGGGATGCCCTAAGACC-3′ | 134 bp 189 bp 286 bp |
| E2+755 | Forward 5′-GAAGACCAAACCTCTCCCTAAGG-3′ Reverse 5′-TACTTTCAGGCCCATCATGCTCC-3′ | 140 bp |
| Site | Ewe Number | X2 | Genotypic Frequency (Number) | Allelic Frequency | |||
|---|---|---|---|---|---|---|---|
| ++ | B+/E+ | BB/EE | B/E | + | |||
| E1+28-30 | 200 | X2 = 13.699 (p = 0.001) | 0.615 (123) | 0.275 (55) | 0.11 (22) | 0.247 | 0.753 |
| E2+755 | 205 | X2 = 26.46 (p = 0.0001) | 0.795 (163) | 0.205 (42) | 0 | 0.102 | 0.898 |
| Scheme | Ewe Number | Genotype | Litter Size |
|---|---|---|---|
| E1+28-30 (B1) | 22 | BB | 1.64 ± 0.181 a |
| 55 | B+ | 2.15 ± 0.114 b | |
| 123 | ++ | 1.90 ± 0.077 ab | |
| E2+755 | 42 | E+ | 2.20 ± 0.129 b |
| 163 | ++ | 1.87 ± 0.066 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Z.-g.; Qin, J.; Jiang, Y.; Ding, X.-D.; Ding, Y.-g.; Tang, S.; Shi, H.-c. The Identification of Mutation in BMP15 Gene Associated with Litter Size in Xinjiang Cele Black Sheep. Animals 2021, 11, 668. https://doi.org/10.3390/ani11030668
Niu Z-g, Qin J, Jiang Y, Ding X-D, Ding Y-g, Tang S, Shi H-c. The Identification of Mutation in BMP15 Gene Associated with Litter Size in Xinjiang Cele Black Sheep. Animals. 2021; 11(3):668. https://doi.org/10.3390/ani11030668
Chicago/Turabian StyleNiu, Zhi-gang, Jin Qin, Yao Jiang, Xiang-Dong Ding, Yu-gong Ding, Sen Tang, and Hong-cai Shi. 2021. "The Identification of Mutation in BMP15 Gene Associated with Litter Size in Xinjiang Cele Black Sheep" Animals 11, no. 3: 668. https://doi.org/10.3390/ani11030668
APA StyleNiu, Z.-g., Qin, J., Jiang, Y., Ding, X.-D., Ding, Y.-g., Tang, S., & Shi, H.-c. (2021). The Identification of Mutation in BMP15 Gene Associated with Litter Size in Xinjiang Cele Black Sheep. Animals, 11(3), 668. https://doi.org/10.3390/ani11030668