
Hibernation Conditions Contribute to the Differential Resistance to Cadmium between Urban and Forest Ant Colonies



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Measured Variables
2.3. Statistical Analyses
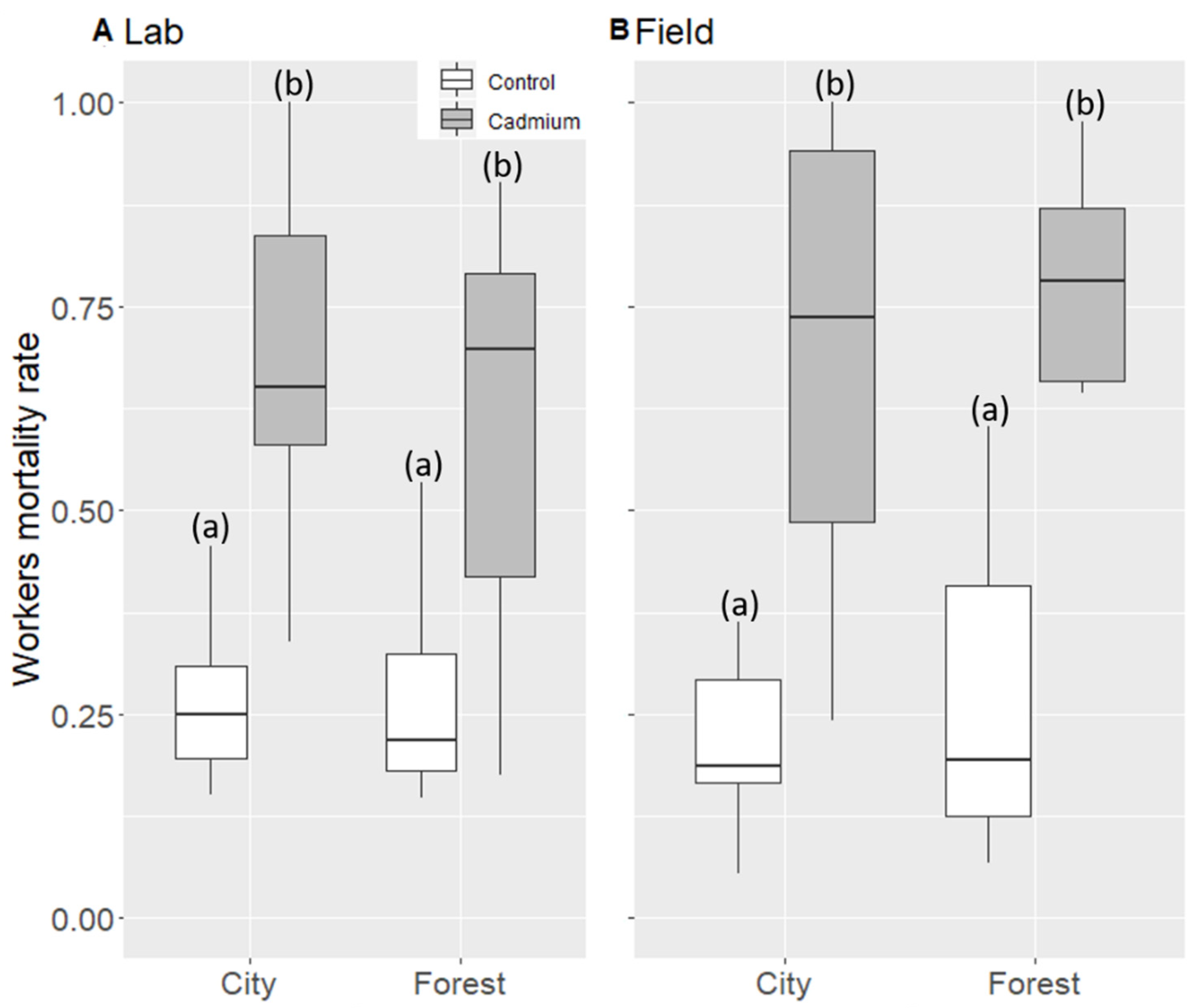
3. Results
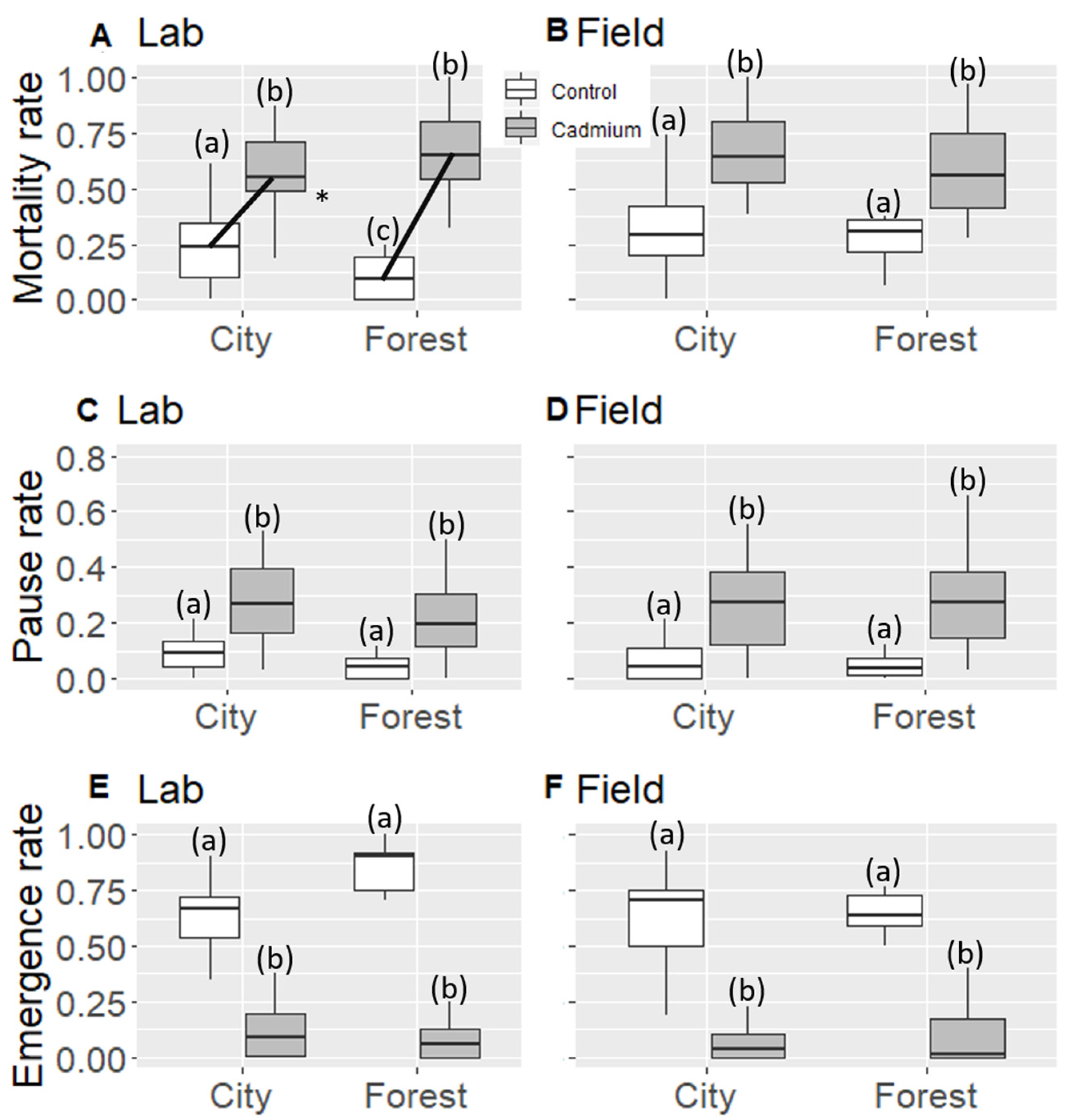
3.1. Worker Mortality Rate
3.2. Larvae Mortality Rate
3.3. Larval Paused Development Rate
3.4. Emergence Rate
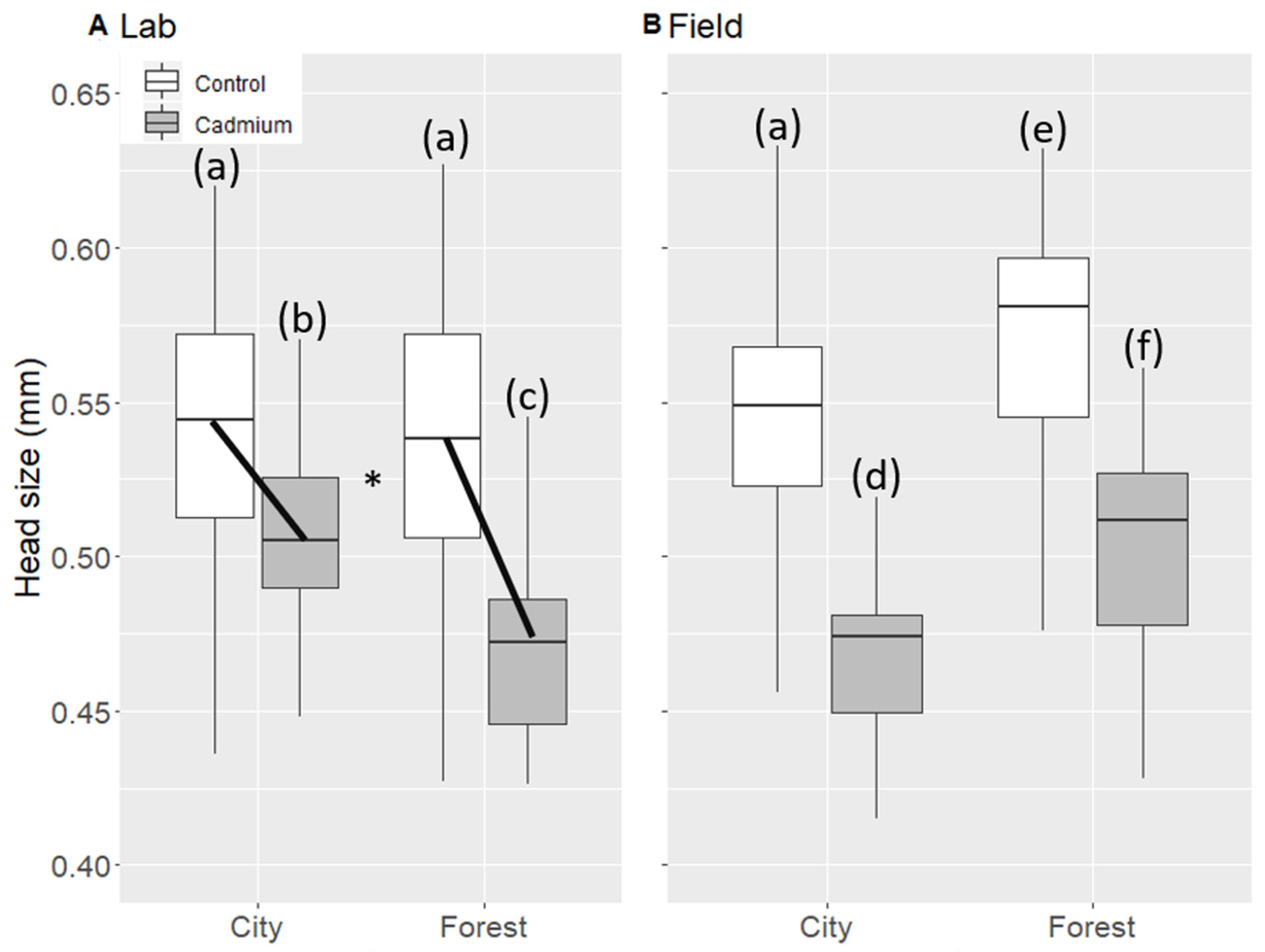
3.5. Size of Lab Workers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guxens, M.; Sunyer, J. A review of epidemiological studies on neuropsychological effects of air pollution. Swiss Med. Wkly. 2012, 141, 13322. [Google Scholar]
- Isaksson, C. Pollution and Its Impact on Wild Animals: A Meta-Analysis on Oxidative Stress. EcoHealth 2010, 7, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, P.; Qi, X.; Zhou, Q.; Zheng, L.; Sun, T.; Yang, Y. DNA changes in barley (Hordeum vulgare) seedlings induced by cadmium pollution using RAPD analysis. Chemosphere 2005, 61, 158–167. [Google Scholar] [CrossRef] [PubMed]
- den Besten, P.J.; Herwig, H.J.; Zandee, D.I.; Voogt, P.A. Effects of cadmium and PCBs on reproduction of the sea star Asterias rubens: Aberrations in the early development. Ecotoxicol. Environ. Saf. 1989, 18, 173–180. [Google Scholar] [CrossRef]
- Malakar, C.; Ganguly, A.; Haldar, P. Influence of cadmium on growth, survival and clutch size of a common Indian short horned grasshopper, Oxya fuscovittata. Am. J. Toxicol. Sci. 2009, 1, 32–36. [Google Scholar]
- Pascoe, D.; Williams, K.A.; Green, D.W.J. Chronic toxicity of cadmium to Chironomus riparius Meigen? Effects upon larval development and adult emergence. Hydrobiologia 1989, 175, 109–115. [Google Scholar] [CrossRef]
- Scott, G.R.; A Sloman, K. The effects of environmental pollutants on complex fish behaviour: Integrating behavioural and physiological indicators of toxicity. Aquat. Toxicol. 2004, 68, 369–392. [Google Scholar] [CrossRef]
- Alexandre, C.V.; Esteves, K.E.; de Moura e Mello, M.A.M. Analysis of fish communities along a rural–urban gradient in a neotropical stream (Piracicaba River Basin, São Paulo, Brazil). Hydrobiologia 2010, 641, 97–114. [Google Scholar] [CrossRef]
- Foti, L.; Dubs, F.; Gignoux, J.; Lata, J.-C.; Lerch, T.Z.; Mathieu, J.; Nold, F.; Nunan, N.; Raynaud, X.; Abbadie, L.; et al. Trace element concentrations along a gradient of urban pressure in forest and lawn soils of the Paris region (France). Sci. Total Environ. 2017, 598, 938–948. [Google Scholar] [CrossRef]
- van Straalen, N.M.; Burghouts, T.B.A.; Doornhof, M.; Groot, G.; Janssen, M.; Joosse, E.; Meerendonk, J.; Theeuven, J.; Verhoef, H.; Zoomer, R. Efficiency of Lead and Cadmium Excretion in Populations of Orchesella cincta (Collembola) from Various Contami-nated Forest Soils. J. Appl. Ecol. 1987, 24, 953–968. [Google Scholar] [CrossRef]
- Mouneyrac, C.; Mastain, O.; Amiard, J.C.; Amiardtriquet, C.; Beaunier, P.; Jeantet, A.-Y.; Smith, B.D.; Rainbow, P.S. Trace-metal detoxification and tolerance of the estuarine worm Hediste diversicolor chronically exposed in their environment. Mar. Biol. 2003, 143, 731–744. [Google Scholar] [CrossRef]
- Roelofs, D.; Janssens, T.K.S.; Timmermans, M.J.T.N.; Nota, B.; Mariën, J.; Bochdanovits, Z.; Ylstra, B.; Van Straalen, N.M. Adaptive differences in gene expression associated with heavy metal tolerance in the soil arthropod Orchesella cincta. Mol. Ecol. 2009, 18, 3227–3239. [Google Scholar] [CrossRef]
- Karouna-Renier, N.K.; Zehr, J.P. Short-term exposures to chronically toxic copper concentrations induce HSP70 proteins in midge larvae (Chironomus tentans). Sci. Total Environ. 2003, 312, 267–272. [Google Scholar] [CrossRef]
- Posthuma, L.; Van Straalen, N.M. Heavy-metal adaptation in terrestrial invertebrates: A review of occurrence, genetics, physiology and ecological consequences. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1993, 106, 11–38. [Google Scholar] [CrossRef]
- Suzuki, K.T.; Yamamura, M.; Mori, T. Cadmium-binding proteins induced in the earthworm. Arch. Environ. Contam. Toxicol. 1980, 9, 415–424. [Google Scholar] [CrossRef]
- Popović, Ž.D.; Subotić, A.; Nikolić, T.V.; Radojicic, R.; Blagojević, D.P.; Grubor-Lajšić, G.; Košťál, V. Expression of stress-related genes in diapause of European corn borer (Ostrinia nubilalis Hbn.). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 186, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, D.L.; Rinehart, J.P.; Yocum, G.D. Stress proteins: A role in insect diapause? In Insect Timing: Circadian Rhythmicity to Seasonality; Denlinger, D.L., Giebultowicz, J.M., Saunders, D.S., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2001; pp. 155–171. [Google Scholar]
- King, A.M.; Macrae, T.H. Insect Heat Shock Proteins during Stress and Diapause. Annu. Rev. Ѐntomol. 2015, 60, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Koštál, V. Eco-physiological phases of insect diapause. J. Insect Physiol. 2006, 52, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Yocum, G. Differential expression of two HSP70 transcripts in response to cold shock, thermoperiod, and adult diapause in the Colorado potato beetle. J. Insect Physiol. 2001, 47, 1139–1145. [Google Scholar] [CrossRef]
- Hahn, D.A.; Denlinger, D.L. Energetics of Insect Diapause. Annu. Rev. Ѐntomol. 2011, 56, 103–121. [Google Scholar] [CrossRef]
- Sinclair, B.J. Linking energetics and overwintering in temperate insects. J. Therm. Biol. 2015, 54, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Grześ, I.M. Ants and heavy metal pollution—A review. Eur. J. Soil Biol. 2010, 46, 350–355. [Google Scholar] [CrossRef]
- Kipyatkov, V.E. Seasonal life cycles and the forms of dormancy in ants (Hymenoptera: Formicoidea). Acta Soc. Zool. Bohem 2001, 65, 211–238. [Google Scholar]
- Haatanen, M.-K.; Van Ooik, T.; Sorvari, J. Effects of overwintering temperature on the survival of the black garden ant (Lasius niger). J. Therm. Biol. 2015, 49–50, 112–118. [Google Scholar] [CrossRef]
- Sorvari, J.; Haatanen, M.-K.; Vesterlund, S.-R. Combined effects of overwintering temperature and habitat degradation on the survival of boreal wood ant. J. Insect Conserv. 2010, 15, 727–731. [Google Scholar] [CrossRef]
- Martelli, A.; Rousselet, E.; Dycke, C.; Bouron, A.; Moulis, J.-M. Cadmium toxicity in animal cells by interference with essential metals. Biochimie 2006, 88, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.J.; Nida, D.L.; Dean, D.A.; England, M.W.; Jacobson, K.B. Resistance of drosophila to cadmium: Biochemical factors in resistant and sensitive strains. Toxicology 1989, 56, 315–321. [Google Scholar] [CrossRef]
- Donker, M.; Bogert, C. Adaptation to cadmium in three populations of the isopod Porcellio scaber. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1991, 100, 143–146. [Google Scholar] [CrossRef]
- Jacquier, L.; Doums, C.; Four-Chaboussant, A.; Peronnet, R.; Tirard, C.; Molet, M. Urban colonies are more resistant to a trace metal than their forest counterparts in the ant Temnothorax nylanderi. Urban Ecosyst. 2020, 1–10. [Google Scholar] [CrossRef]
- Herbers, J.M. Community structure in north temperate ants: Temporal and spatial variation. Oecologia 1989, 81, 201–211. [Google Scholar] [CrossRef]
- Bornstein, R.D. Observations of the Urban Heat Island Effect in New York City. J. Appl. Meteorol. 1968, 7, 575–582. [Google Scholar] [CrossRef]
- Rosenfeld, D. Suppression of Rain and Snow by Urban and Industrial Air Pollution. Science 2000, 287, 1793–1796. [Google Scholar] [CrossRef] [PubMed]
- Scharf, I.; Modlmeier, A.P.; Beros, S.; Foitzik, S. Ant Societies Buffer Individual-Level Effects of Parasite Infections. Am. Nat. 2012, 180, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.B.; Tschinkel, W.R. Worker Allometry in Relation to Colony Size and Social form in the Fire Ant Solenopsis invicta. J. Insect Sci. 2010, 10, 1–10. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- RC Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Australia, 2019; Available online: https://www.R-project.org\ (accessed on 2 January 2021).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; RC Team. NLME: Linear and Nonlinear Mixed Effects Models, 2015; R Package Version 3.1-122. Available online: http://CRAN.R289project.org/package=nlme (accessed on 21 January 2021).
- Hadley, W. Ggplot2: Elegrant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Andrew, S.C.; Taylor, M.P.; Lundregan, S.; Lien, S.; Jensen, H.; Griffith, S.C. Signs of adaptation to trace metal contamination in a common urban bird. Sci. Total Environ. 2019, 650, 679–686. [Google Scholar] [CrossRef]
- Williams, J.B.; Shorthouse, J.D.; Lee, R.E. Deleterious effects of mild simulated overwintering temperatures on survival and potential fecundity of rose-galling Diplolepis wasps (Hymenoptera: Cynipidae). J. Exp. Zool. 2003, 298, 23–31. [Google Scholar] [CrossRef]
- Fisker, K.V.; Holmstrup, M.; Sørensen, J.G. Freezing of body fluids induces metallothionein gene expression in earthworms (Dendrobaena octaedra). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 179, 44–48. [Google Scholar] [CrossRef]
- Reid, N.M.; Proestou, D.A.; Clark, B.W.; Warren, W.C.; Colbourne, J.K.; Shaw, J.R.; Karchner, S.I.; Hahn, M.E.; Nacci, D.; Oleksiak, M.F.; et al. The genomic landscape of rapid repeated evolutionary adaptation to toxic pollution in wild fish. Science 2016, 354, 1305–1308. [Google Scholar] [CrossRef] [Green Version]
- Khimoun, A.; Doums, C.; Molet, M.; Kaufmann, B.; Peronnet, R.; Eyer, P.A.; Mona, S. Urbanization without isolation: The absence of genetic structure among cities and forests in the tiny acorn ant Temnothorax nylanderi. Biol. Lett. 2020, 16, 20190741. [Google Scholar] [CrossRef] [Green Version]
- Watson, H.; Videvall, E.; Andersson, M.N.; Isaksson, C. Transcriptome analysis of a wild bird reveals physiological responses to the urban environment. Sci. Rep. 2017, 7, 44180. [Google Scholar] [CrossRef] [PubMed]
- Cassone, B.J.; Kamdem, C.; Cheng, C.; Tan, J.C.; Hahn, M.W.; Costantini, C.; Besansky, N.J. Gene expression divergence between malaria vector sibling species Anopheles gambiae and An. coluzzii from rural and urban Yaoundé Cameroon. Mol. Ecol. 2014, 23, 2242–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, S.E.; Munshi-South, J.; Obergfell, C.; O’Neill, R. Signatures of Rapid Evolution in Urban and Rural Transcriptomes of White-Footed Mice (Peromyscus leucopus) in the New York Metropolitan Area. PLoS ONE 2013, 8, e74938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guengerich, F.P. Common and Uncommon Cytochrome P450 Reactions Related to Metabolism and Chemical Toxicity. Chem. Res. Toxicol. 2001, 14, 611–650. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Worker Mortality Rate | |
|---|---|
| T:H:C | F125,119 = 0.87 |
| p = 0.52 | |
| Treatment–Habitat | F125,123 = 0.0099 |
| p = 0.90 | |
| Treatment–Conditions | F125,123 = 1.98 |
| p = 0.14 | |
| Habitat–Conditions | F125,122 = 0.75 |
| p = 0.52 | |
| Treatment | F125,126 = 4.91 |
| p = 0.028 | |
| Habitat | F125,124 = 0.058 |
| P = 0.81 | |
| Conditions | F125,124 = 1.45 |
| p = 0.23 | |
| Colony Size–Treatment | F125,126 = 6.2 |
| p = 0.014 | |
| Colony Size | NA |
| Larvae Mortality Rate | Size of Lab Workers | |||
|---|---|---|---|---|
| Treatment–Habitat | Lab | Field | Lab | Field |
| F56,57 = 5.33 | F62,60 = 0.18 | X1 = 4.06 | X1 = 0.50 | |
| p = 0.024 | p = 0.83 | p = 0.043 | p = 0.47 | |
| Treatment | Z = −7.59 | F62,63 = 18.41 | t65 = 6.52 | X1 = 50.73 |
| p < 0.001 | p < 0.0001 | p < 0.001 | p < 0.0001 | |
| Habitat | Z = −1.30 | F61,62 = 0.20 | t63 = 0.88 | X1 = 12.84 |
| p = 0.19 | p = 0.65 | p = 0.38 | p = 0.0003 | |
| Colony Size–Treatment | F57,58 = 1.69 | F62,63 = 4.57 | X1 = 0.17 | X2 = 3.05 |
| p = 0.19 | p = 0.036 | p = 0.67 | p = 0.22 | |
| Colony Size | F58,59 = 3.63 | NA | X1 = 4.46 | X1 = 2.91 |
| p = 0.061 | p = 0.035 | p = 0.088 | ||
| Pause Rate | Emergence Rate | |
|---|---|---|
| T:H:C | F121,127 = 0.78 | F126,121 = 1.29 |
| p = 0.58 | p = 0.26 | |
| Treatment–Habitat | F127,125 = 1.64 | F126,124 = 1.33 |
| p = 0.19 | p = 0.26 | |
| Treatment–Conditions | F127,125 = 0.37 | F126,125 = 0.040 |
| p=0.69 | p = 0.84 | |
| Habitat–Conditions | F127,124 = 0.87 | F126,124 = 1.09 |
| p = 0.46 | p = 0.33 | |
| Treatment | F127,128 = 100.14 | F126,127 = 257.29 |
| p < 0.0001 | p < 0.0001 | |
| Habitat | F127,126 = 1.69 | F126,125 = 1.82 |
| p = 0.19 | p = 0.17 | |
| Conditions | F127,126 = 0.034 | F126,127 = 6.64 |
| p = 0.85 | p = 0.011 | |
| Colony Size–Treatment | F127,125 = 0.17 | F126,124 = 0.50 |
| p = 0.84 | p = 0.60 | |
| Colony Size | F127,126 = 0.32 | F126,125 = 0.23 |
| p = 0.57 | p = 0.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacquier, L.; Molet, M.; Bocquet, C.; Doums, C. Hibernation Conditions Contribute to the Differential Resistance to Cadmium between Urban and Forest Ant Colonies. Animals 2021, 11, 1050. https://doi.org/10.3390/ani11041050
Jacquier L, Molet M, Bocquet C, Doums C. Hibernation Conditions Contribute to the Differential Resistance to Cadmium between Urban and Forest Ant Colonies. Animals. 2021; 11(4):1050. https://doi.org/10.3390/ani11041050
Chicago/Turabian StyleJacquier, Lauren, Mathieu Molet, Céline Bocquet, and Claudie Doums. 2021. "Hibernation Conditions Contribute to the Differential Resistance to Cadmium between Urban and Forest Ant Colonies" Animals 11, no. 4: 1050. https://doi.org/10.3390/ani11041050
APA StyleJacquier, L., Molet, M., Bocquet, C., & Doums, C. (2021). Hibernation Conditions Contribute to the Differential Resistance to Cadmium between Urban and Forest Ant Colonies. Animals, 11(4), 1050. https://doi.org/10.3390/ani11041050