
A Machine Learning Approach to Study Demographic Alterations in Honeybee Colonies Using SDS–PAGE Fingerprinting

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Single Cohort Colonies Setup and Sample Collection
- On day −21, four mated sister queens were caged on four different combs, drawn from organic-certified residue free wax, and placed inside four fully developed and healthy colonies;
- On day −19 the queens were removed from the cages, in order to have a maximum difference of 48 h among the brood laid;
- On day +1, newly eclosed workers were gently brushed from the combs, mixed to eliminate the mother colony factor and used to prepare the two SCCs. Each SCCs was made with 250 g of Apis mellifera ligustica bees (equivalent to approximately 2500 individuals), one queen (of the same subspecies) and two combs drawn from the same wax mentioned above: one empty and one with plenty of honey and pollen. The SCCs were kept closed in a protected and shaded environment to allow complete maturation of the workers;
- On day +3, 3 days post-eclosion, the SCCs were moved to the outdoor apiary.
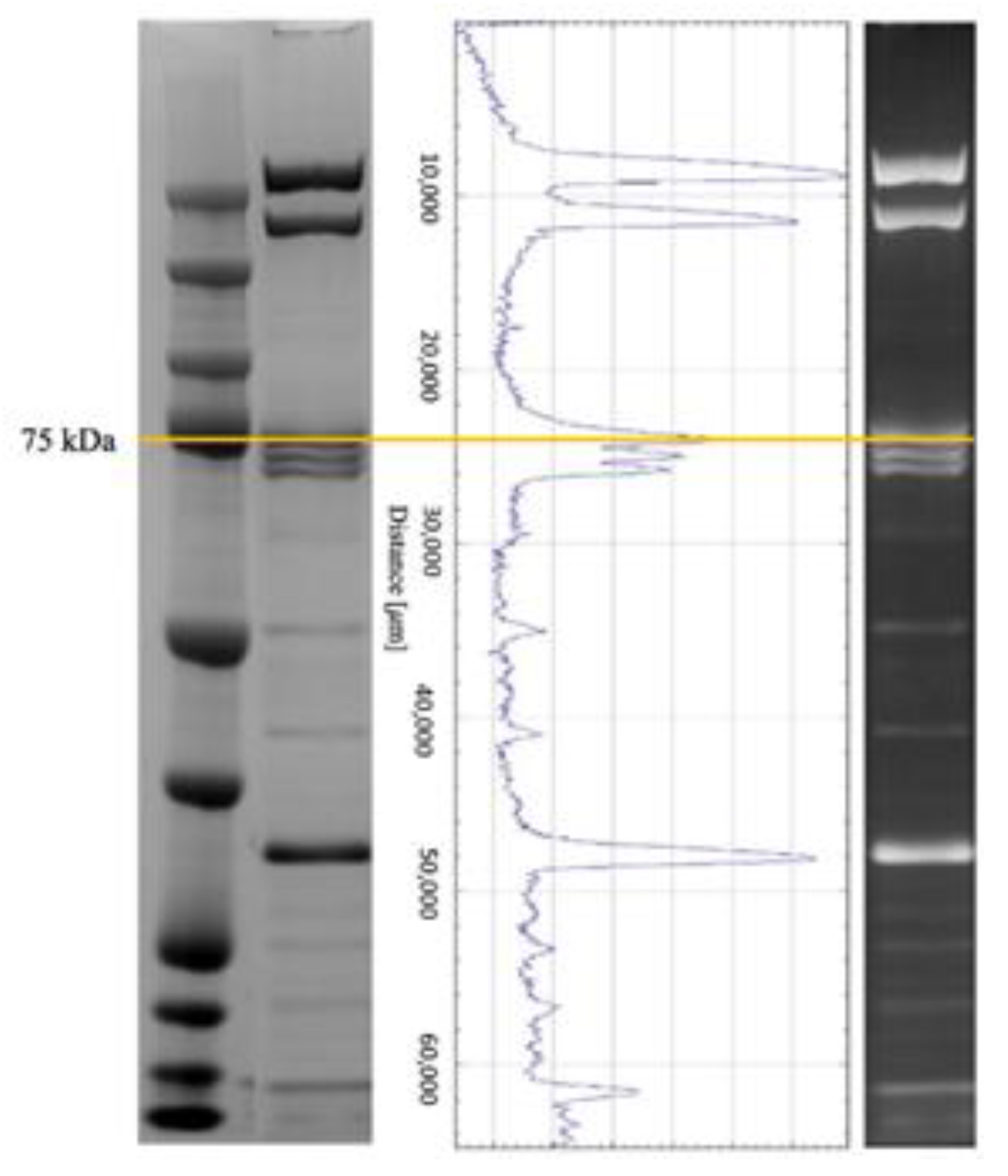
2.2. Haemolymph Collection and SDS-PAGE Electrophoresis
2.3. Data Preparation
2.4. Statistical Analysis
3. Results
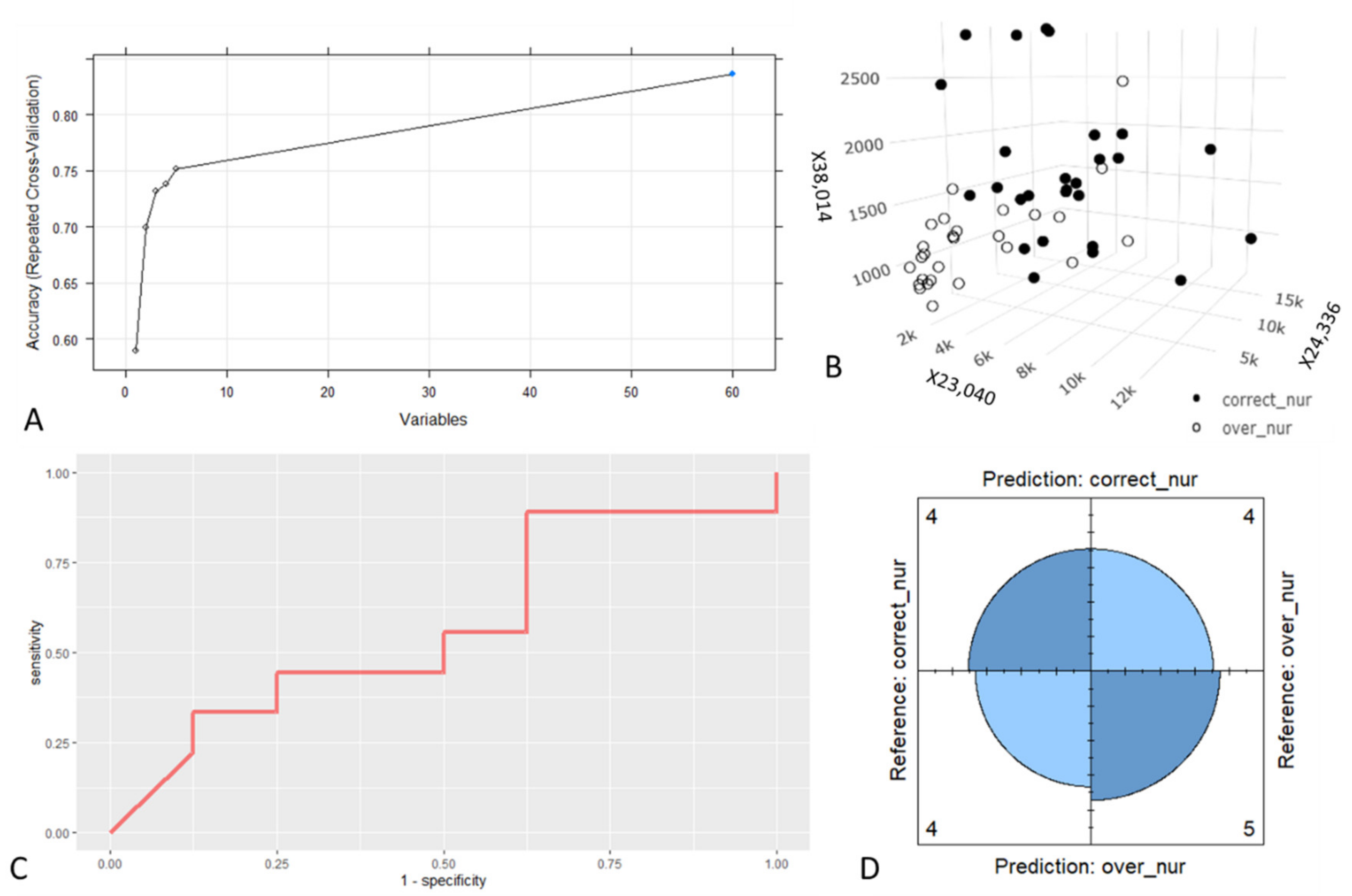
3.1. Nurses
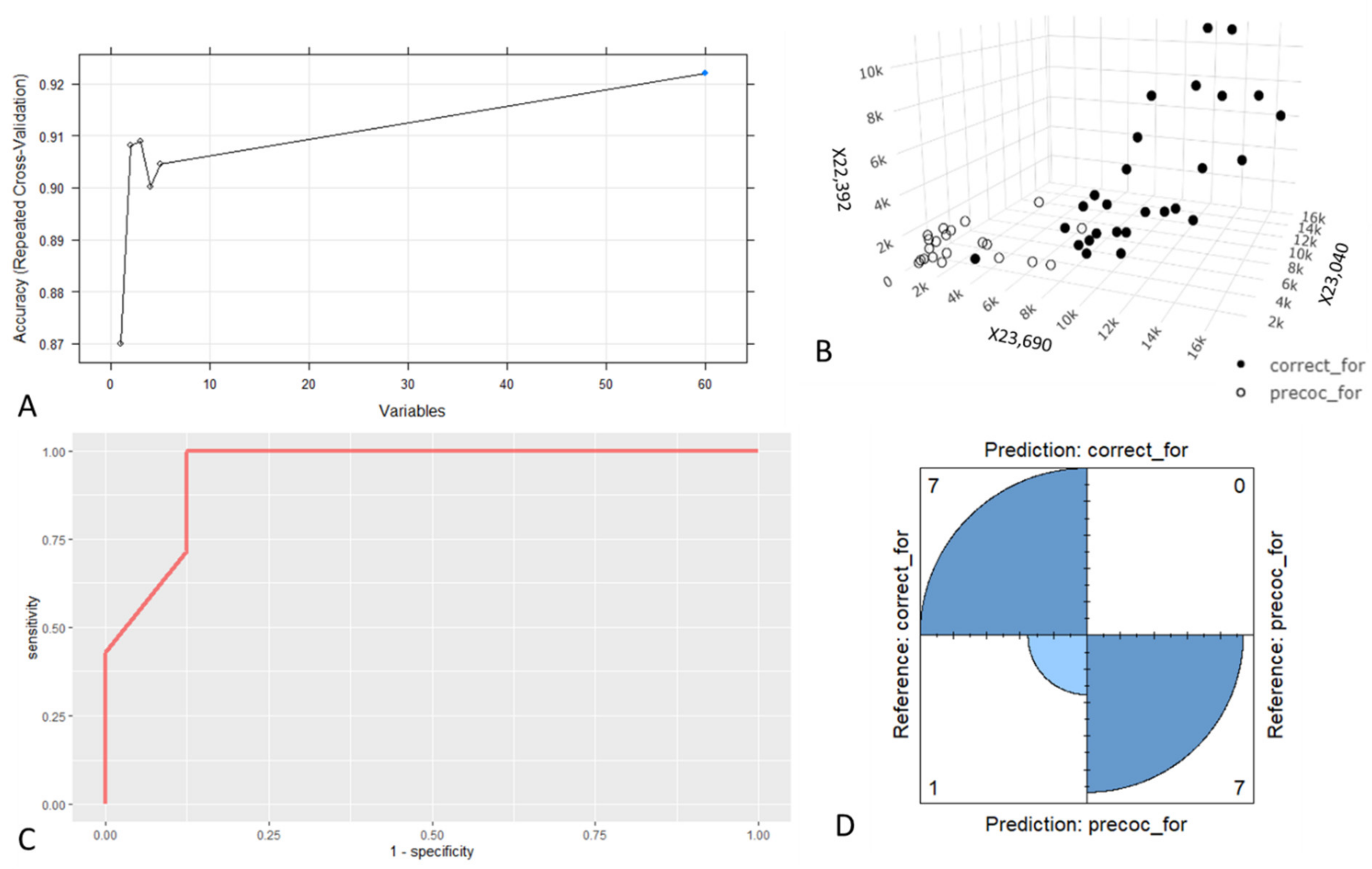
3.2. Foragers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Potts, S.G.; Roberts, S.P.M.; Dean, R.; Marris, G.; Brown, M.A.; Jones, R.; Neumann, P.; Settele, J. Declines of managed honey bees and beekeepers in Europe. J. Apic. Res. 2010, 49, 15–22. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Delaplane, K.S. Emergent properties in the honey bee superorganism. Bee World 2017, 94, 8–15. [Google Scholar] [CrossRef]
- Amdam, G.V.; Page, R.E. Intergenerational transfers may have decoupled physiological and chronological age in a eusocial insect. Ageing Res. Rev. 2005, 4, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Cabbri, R.; Ferlizza, E.; Nanetti, A.; Monari, E.; Andreani, G.; Galuppi, R.; Isani, G. Biomarkers of nutritional status in honeybee haemolymph: Effects of different biotechnical approaches for Varroa destructor treatment and wintering phase. Apidologie 2018, 49, 606–618. [Google Scholar] [CrossRef] [Green Version]
- Omholt, S.W.; Amdam, G.V. Epigenetic regulation of aging in honeybee workers. Sci. Aging Knowl. Environ. 2004, 2004, pe28. [Google Scholar] [CrossRef] [PubMed]
- Maurizio, A. The influence of pollen feeding and brood rearing on the lenght of life and physiological condition of the honeybee. Bee World 1950, 31, 9–12. [Google Scholar] [CrossRef]
- Naiem, E.S.; Hrassnigg, N.; Crailsheim, K. Nurse bees support the physiological development of young bees (Apis mellifera L.). J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 1999, 169, 271–279. [Google Scholar] [CrossRef]
- Amdam, G.V. Social context, stress, and plasticity of aging. Aging Cell 2011, 10, 18–27. [Google Scholar] [CrossRef]
- Seeley, T.D. Adaptive significance of the age polyethism schedule in honeybee colonies. Behav. Ecol. Sociobiol. 1982, 11, 287–293. [Google Scholar] [CrossRef]
- Amdam, G.V.; Norberg, K.; Omholt, S.W.; Kryger, P.; Lourenço, A.P.; Bitondi, M.M.G.; Simões, Z.L.P. Higher vitellogenin concentrations in honey bee workers may be an adaptation to life in temperate climates. Insectes Soc. 2005, 52, 316–319. [Google Scholar] [CrossRef]
- Amdam, G.V.; Omholt, S.W. The hive bee to forager transition in honeybee colonies: The double repressor hypothesis. J. Theor. Biol. 2003, 223, 451–464. [Google Scholar] [CrossRef]
- Guidugli, K.R.; Nascimento, A.M.; Amdam, G.V.; Barchuk, A.R.; Omholt, S.; Simões, Z.L.P.; Hartfelder, K. Vitellogenin regulates hormonal dynamics in the worker caste of a eusocial insect. FEBS Lett. 2005, 579, 4961–4965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.Y.; Robinson, G.E. Honeybee colony integration: Worker-worker interactions mediate hormonally regulated plasticity in division of labor. Proc. Natl. Acad. Sci. USA 1992, 89, 11726–11729. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.-Y.; Robinson, G.E. Regulation of honey bee division of labor by colony age demography. Behav. Ecol. Sociobiol. 1996, 39, 147–158. [Google Scholar] [CrossRef]
- Fergusson, L.A.; Winston, M.L. The influence of wax deprivation on temporal polyethism in honey bee (Apis mellifera L.) colonies. Can. J. Zool. 1988, 66, 1997–2001. [Google Scholar] [CrossRef]
- Janmaat, A.F.; Winston, M.L. The influence of pollen storage area and Varroa jacobsoni Oudemans parasitism on temporal caste structure in honey bees (Apis mellifera L.). Insectes Soc. 2000, 47, 177–182. [Google Scholar] [CrossRef]
- Schulz, D.J.; Huang, Z.Y.; Robinson, G.E. Effects of colony food shortage on behavioral development in honey bees. Behav. Ecol. Sociobiol. 1998, 42, 295–303. [Google Scholar] [CrossRef]
- Toth, A.L.; Robinson, G.E. Worker nutrition and division of labour in honeybees. Anim. Behav. 2005, 69, 427–435. [Google Scholar] [CrossRef]
- Vance, J.T.; Williams, J.B.; Elekonich, M.M.; Roberts, S.P.R. The effects of age and behavioral development on honey bee (Apis mellifera) flight performance. J. Exp. Biol. 2009, 212, 2604–2611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goblirsch, M.; Huang, Z.Y.; Spivak, M. Physiological and behavioral changes in honey bees (Apis mellifera) induced by Nosema ceranae infection. PLoS ONE 2013, 8, e58165. [Google Scholar] [CrossRef]
- Higes, M.; Martín-Hernández, R.; Botías, C.; Bailón, E.G.; González-Porto, A.V.; Barrios, L.; Del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J.; Palencia, P.G.; et al. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Doublet, V.; Poeschl, Y.; Gogol-Döring, A.; Alaux, C.; Annoscia, D.; Aurori, C.; Barribeau, S.M.; Bedoya-Reina, O.C.; Brown, M.J.F.; Bull, J.C.; et al. Unity in defence: Honeybee workers exhibit conserved molecular responses to diverse pathogens. BMC Genom. 2017, 18, 207. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.J.; Søvik, E.; Myerscough, M.R.; Barron, A.B. Rapid behavioral maturation accelerates failure of stressed honey bee colonies. Proc. Natl. Acad. Sci. USA. 2015, 112, 3427–3432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alaux, C.; Soubeyrand, S.; Prado, A.; Peruzzi, M.; Maisonnasse, A.; Vallon, J.; Hernandez, J.; Jourdan, P.; Le Conte, Y. Measuring biological age to assess colony demographics in honeybees. PLoS ONE 2018, 13, e0209192. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.E.; Page, R.E.; Strambi, C.; Strambi, A. Hormonal and genetic control of behavioral integration in honey bee colonies. Science 1989, 246, 109–112. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- RStudio Team. Rstudio: Integrated Development Environment for R; RStudio: PBC, Boston, MA, USA, 2020. [Google Scholar]
- Ottensmann, M.; Stoffel, M.A.; Nichols, H.J.; Hoffman, J.I. GCalignR: An R package for aligning gas-chromatography data for ecological and evolutionary studies. PLoS ONE 2018, 13, e0198311. [Google Scholar] [CrossRef] [Green Version]
- Ambroise, C.; McLachlan, G.J. Selection bias in gene extraction on the basis of microarray gene-expression data. Proc. Natl. Acad. Sci. USA 2002, 99, 6562–6566. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Wegener, J.; Lorenz, M.W.; Bienefeld, K. Physiological consequences of prolonged nursing in the honey bee. Insectes Soc. 2009, 56, 85–93. [Google Scholar] [CrossRef]
- Mattila, H.R.; Reeve, H.K.; Smith, M.L. Promiscuous honey bee queens increase colony productivity by suppressing worker selfishness. Curr. Biol. 2012, 22, 2027–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbs, S.L.; Ryan, R.O.; Schmidt, J.O.; Keim, P.S.; Law, J.H. Lipophorin of the larval honeybee, Apis mellifera L. J. Lipid Res. 1985, 26, 241–247. [Google Scholar] [CrossRef]
- Kawooya, J.K.; Meredith, S.C.; Wells, M.A.; Kézdy, F.J.; Law, J.H. Physical and surface properties of insect apolipophorin III. J. Biol. Chem. 1986, 261, 13588–13591. [Google Scholar] [CrossRef]
- Ryan, R.O.; van der Horst, D.J. Lipid transport biochemistry and its role in energy production. Annu. Rev. Entomol. 2000, 45, 233–260. [Google Scholar] [CrossRef]
- Wen, D.; Luo, H.; Li, T.; Wu, C.; Zhang, J.; Wang, X.; Zhang, R. Cloning and characterization of an insect apolipoprotein (apolipophorin-II/I) involved in the host immune response of Antheraea pernyi. Dev. Comp. Immunol. 2017, 77, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Dolan, J.; Walshe, K.; Alsbury, S.; Hokamp, K.; O’Keeffe, S.; Okafuji, T.; Miller, S.F.C.; Tear, G.; Mitchell, K.J. The extracellular Leucine-rich repeat superfamily; a comparative survey and analysis of evolutionary relationships and expression patterns. BMC Genom. 2007, 8, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, G.; Liu, Z.H.; Zhao, M.; Wang, H.L.; Zhou, B. Transferrin 1 functions in iron trafficking and genetically interacts with ferritin in Drosophila melanogaster. Cell Rep. 2019, 26, 748–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iatsenko, I.; Marra, A.; Boquete, J.P.; Peña, J.; Lemaitre, B. Iron sequestration by transferrin 1 mediates nutritional immunity in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2020, 117, 7317–7325. [Google Scholar] [CrossRef] [PubMed]
- Vannette, R.L.; Mohamed, A.; Johnson, B.R. Forager bees (Apis mellifera) highly express immune and detoxification genes in tissues associated with nectar processing. Sci. Rep. 2015, 5, 16224. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Nurses | Foragers | |||||
|---|---|---|---|---|---|---|
| Model | AUC | Sensitivity | Specificity | AUC | Sensitivity | Specificity |
| SVM | 0.87 (a) | 0.76 (a) | 0.83 (a) | 0.98 (a) | 0.96 (a) | 0.86 (a) |
| KNN | 0.83 (a) | 0.9 (b) | 0.67 (b) | 0.98 (a) | 0.96 (a) | 0.87 (a) |
| RF | 0.82 (a) | 0.82 (ab) | 0.75 (ab) | 0.96 (a) | 0.89 (b) | 0.90 (a) |
| Nurses | Foragers | |
|---|---|---|
| RF | KNN | |
| Accuracy | 0.53 | 0.93 |
| Accuracy Lower | 0.28 | 0.68 |
| Accuracy Upper | 0.77 | 1 |
| Accuracy Null | 0.53 | 0.53 |
| Accuracy p-Value | 0.6 | 0.00113 |
| Sensitivity | 0.5 | 0.88 |
| Specificity | 0.56 | 1 |
| AUC | 0.57 | 0.95 |
| Accession 1 | Description | Mass (kD) 2 | Score 3 | Pep 4 | Pep (sig) 5 | Seq 6 | Seq (sig) 7 | Protein Homologous 8 | % Identity 9 | Species 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| A0A088AS56 | Uncharacterised protein | 369 | 3848 | 410 | 224 | 48 | 35 | Apolipophorins | 91.8 | Apis cerana |
| A0A088AQB0 | Uncharacterised protein | 76 | 1001 | 98 | 59 | 19 | 13 | Leucine-rich repeat-containing protein 15 | 98.4 | Apis cerana |
| A0A088AFH7 | Transferrin | 80 | 340 | 55 | 24 | 19 | 9 | 100 | Apis mellifera |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabbri, R.; Ferlizza, E.; Bellei, E.; Andreani, G.; Galuppi, R.; Isani, G. A Machine Learning Approach to Study Demographic Alterations in Honeybee Colonies Using SDS–PAGE Fingerprinting. Animals 2021, 11, 1823. https://doi.org/10.3390/ani11061823
Cabbri R, Ferlizza E, Bellei E, Andreani G, Galuppi R, Isani G. A Machine Learning Approach to Study Demographic Alterations in Honeybee Colonies Using SDS–PAGE Fingerprinting. Animals. 2021; 11(6):1823. https://doi.org/10.3390/ani11061823
Chicago/Turabian StyleCabbri, Riccardo, Enea Ferlizza, Elisa Bellei, Giulia Andreani, Roberta Galuppi, and Gloria Isani. 2021. "A Machine Learning Approach to Study Demographic Alterations in Honeybee Colonies Using SDS–PAGE Fingerprinting" Animals 11, no. 6: 1823. https://doi.org/10.3390/ani11061823
APA StyleCabbri, R., Ferlizza, E., Bellei, E., Andreani, G., Galuppi, R., & Isani, G. (2021). A Machine Learning Approach to Study Demographic Alterations in Honeybee Colonies Using SDS–PAGE Fingerprinting. Animals, 11(6), 1823. https://doi.org/10.3390/ani11061823