
Genetic Estimates for Growth and Shape-Related Traits in the Flatfish Senegalese Sole

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
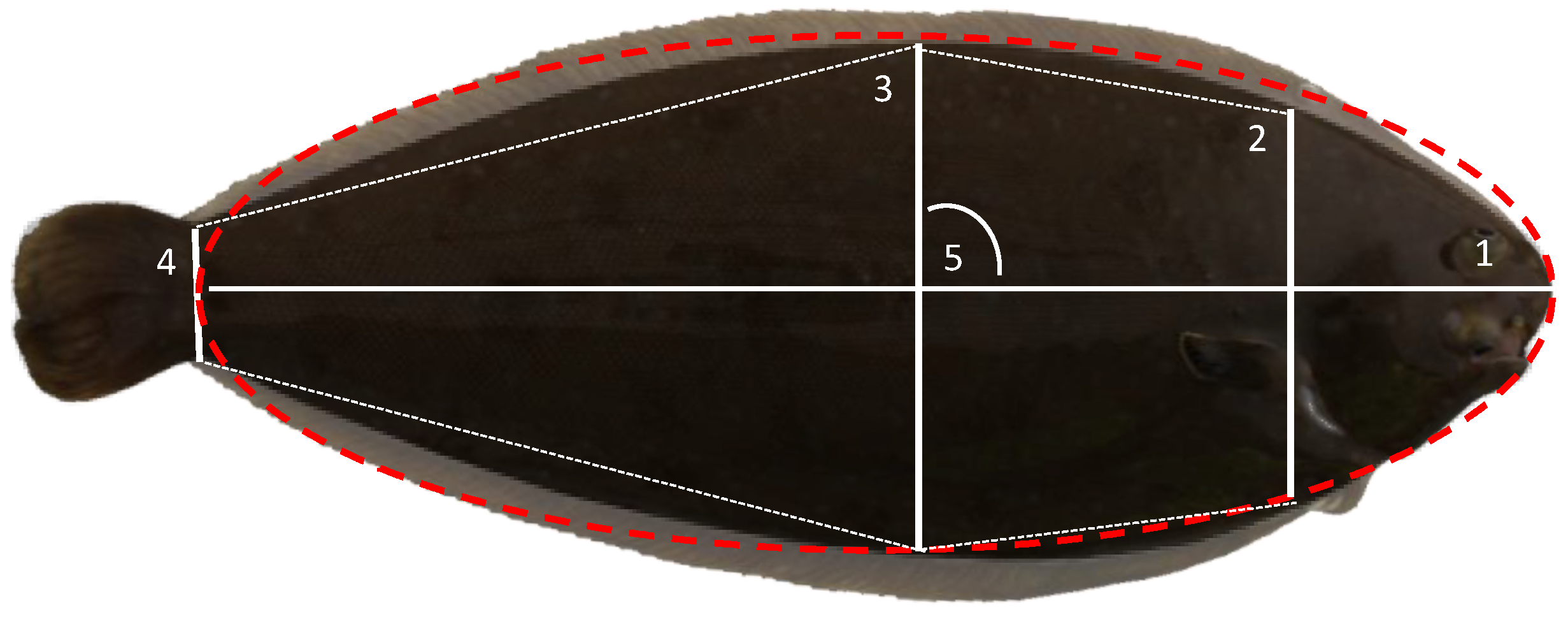
2. Materials and Methods
2.1. Animals
2.2. DNA Isolation and Parentage Assignment
2.3. Statistical Analysis and Genetic Parameters
3. Results
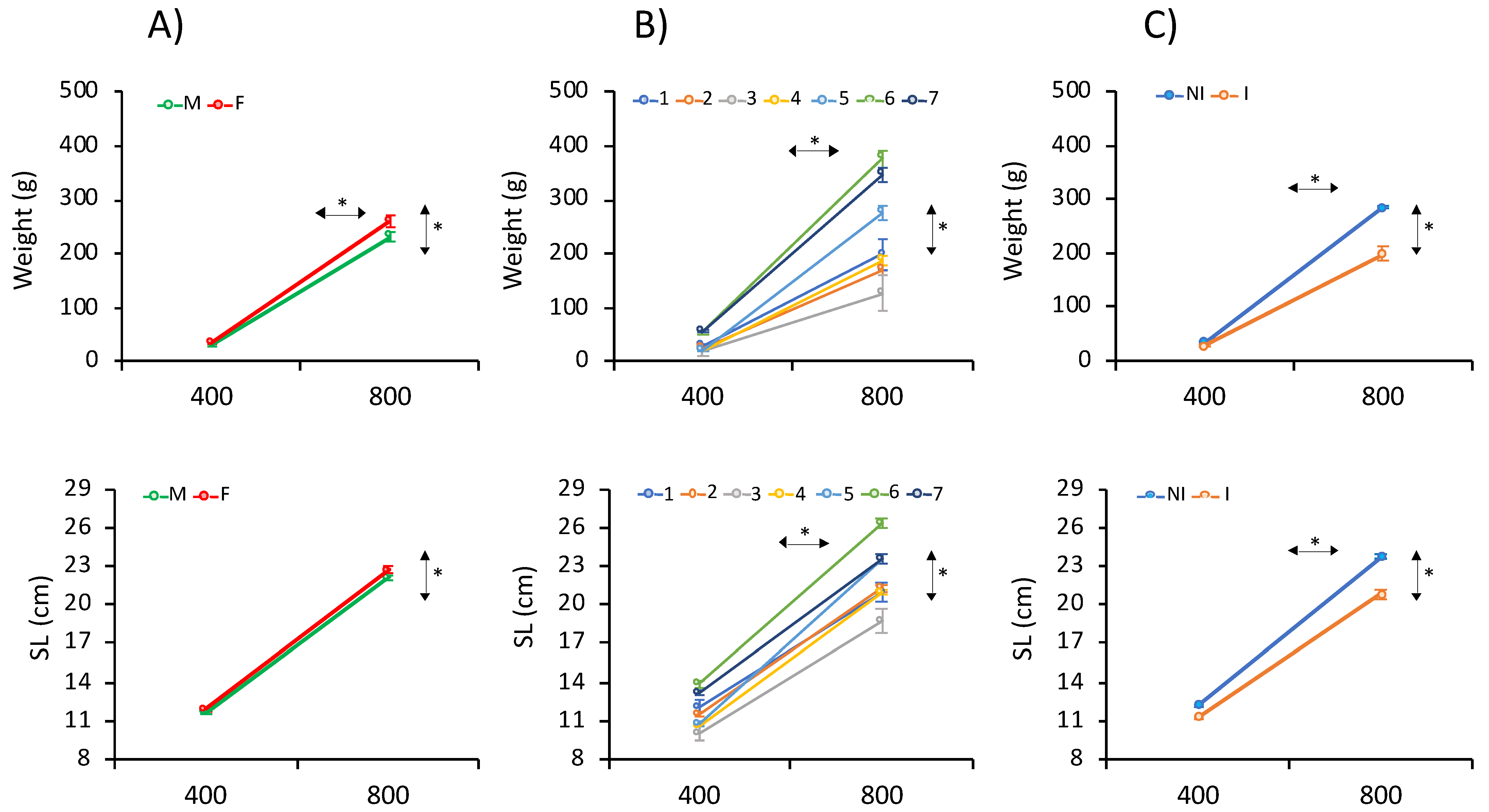
3.1. Phenotypic Data for Growth Traits
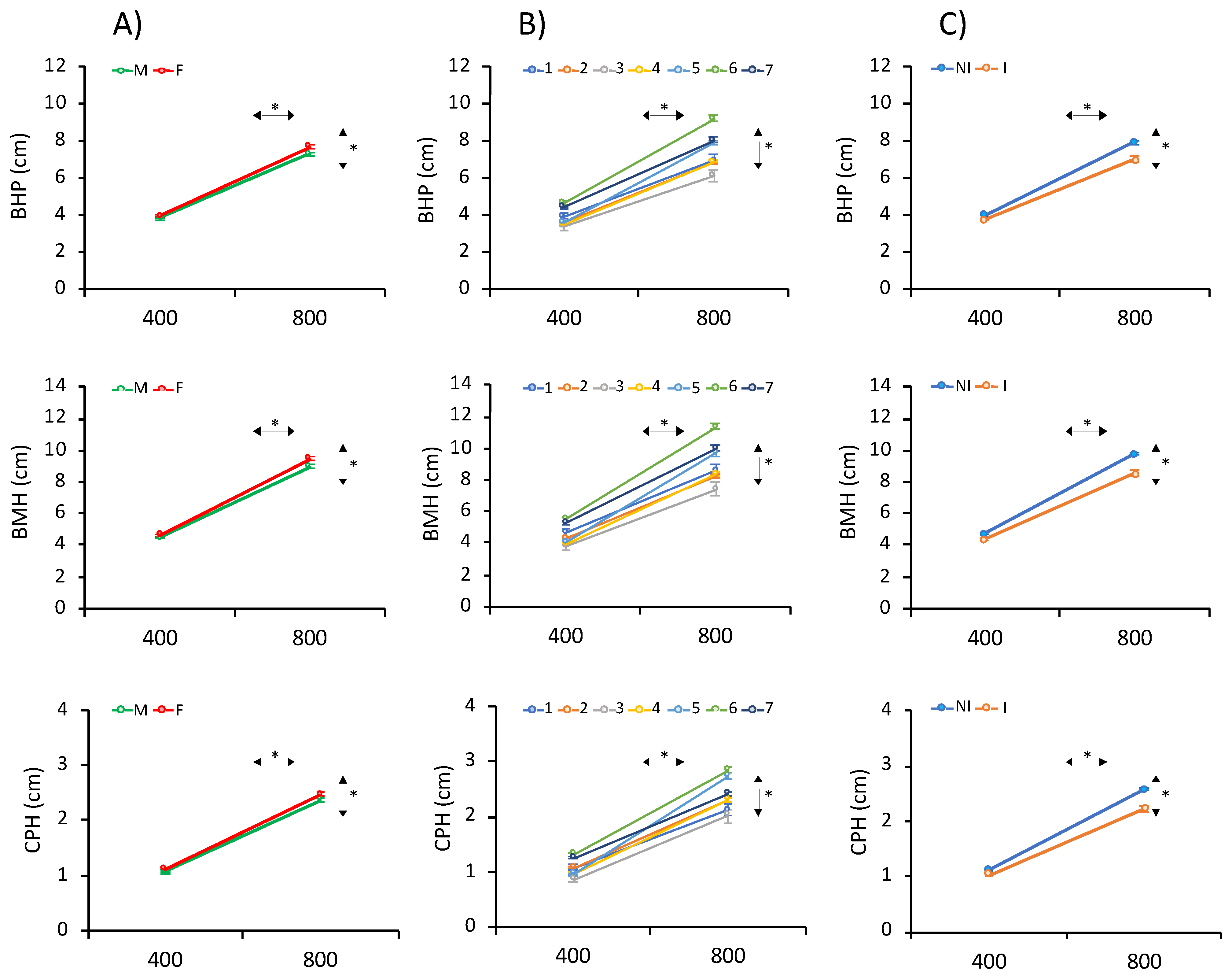
3.2. Phenotypic Data for Height Traits
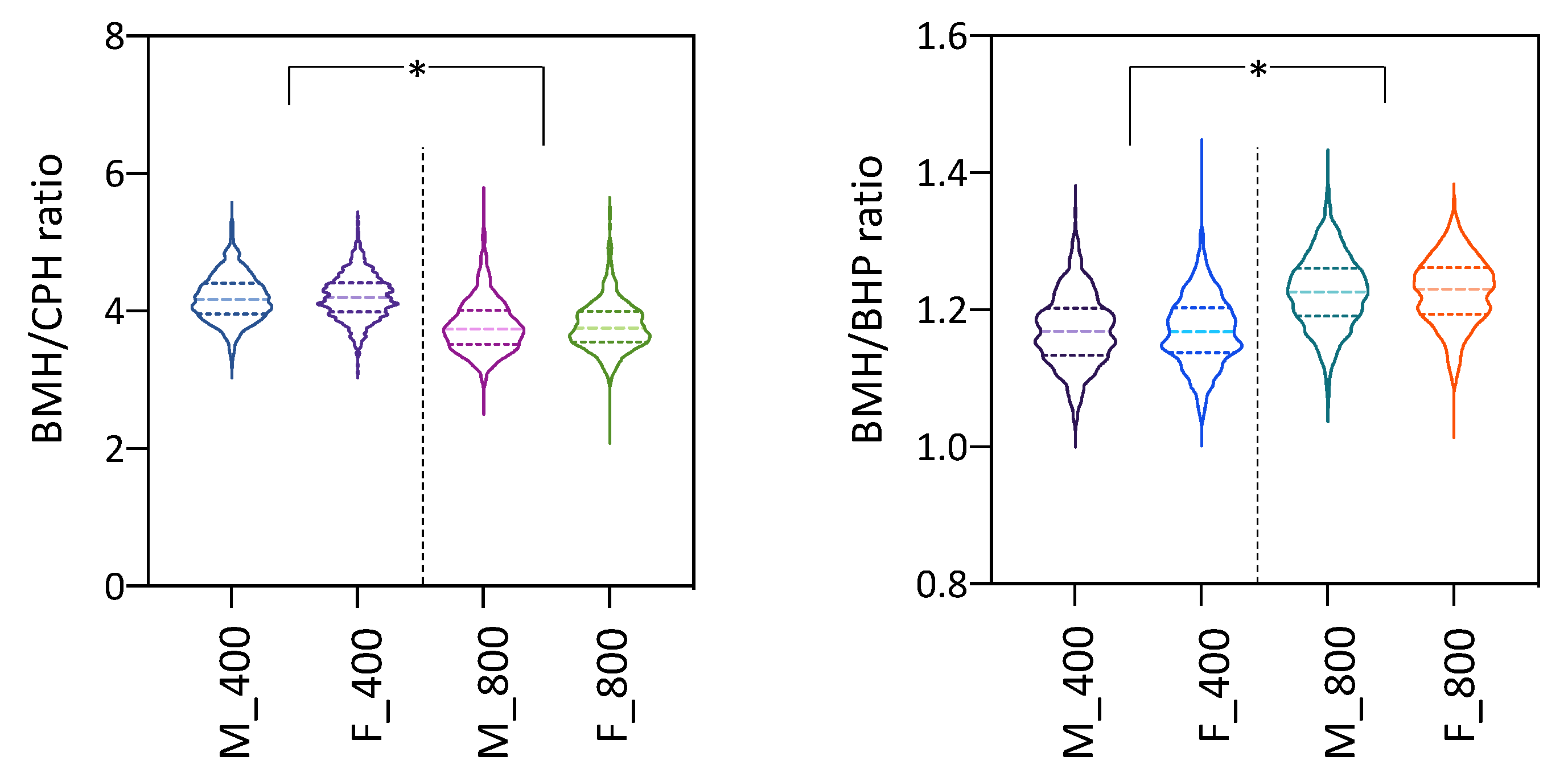
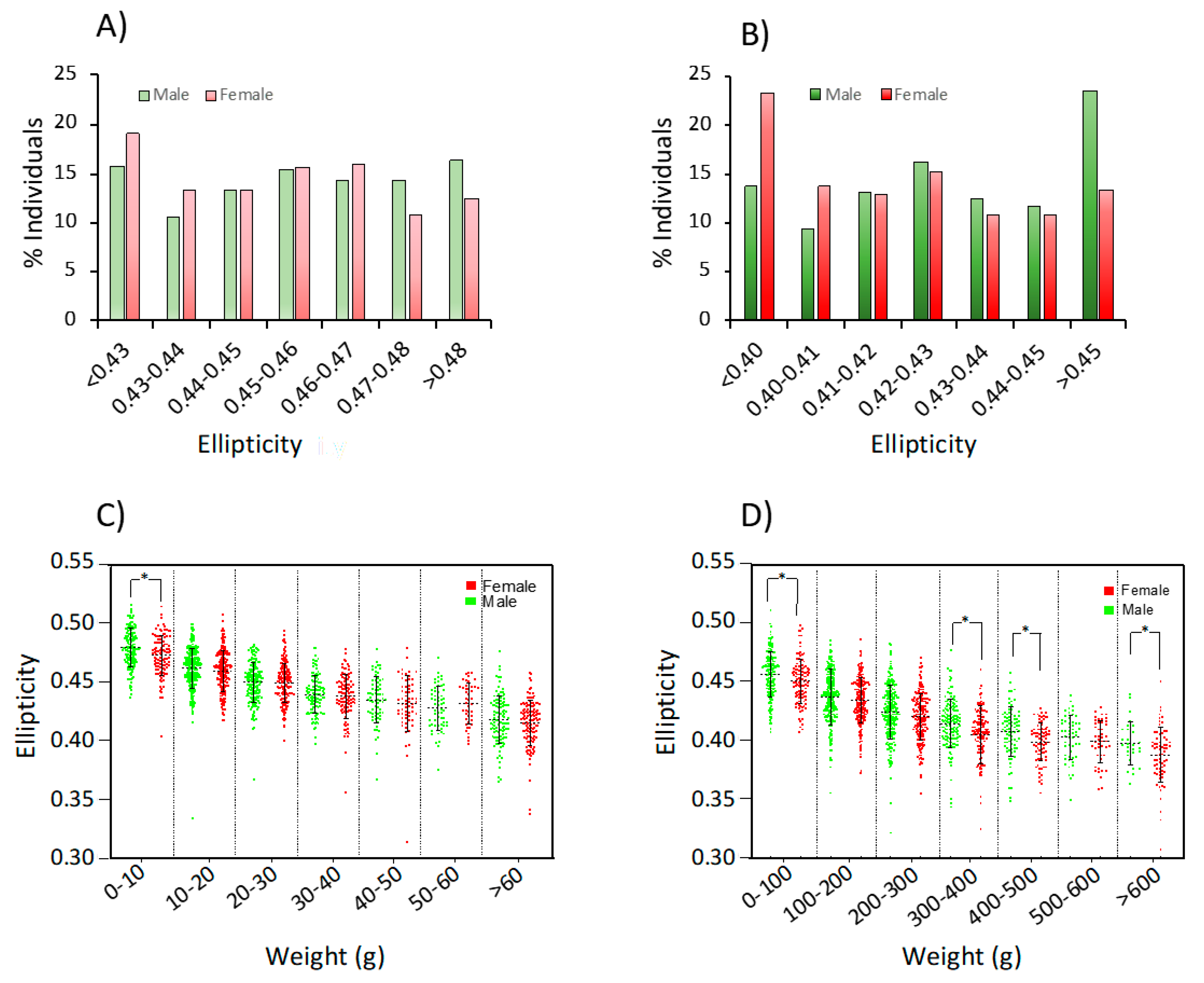
3.3. Phenotypic Data for Ellipticity
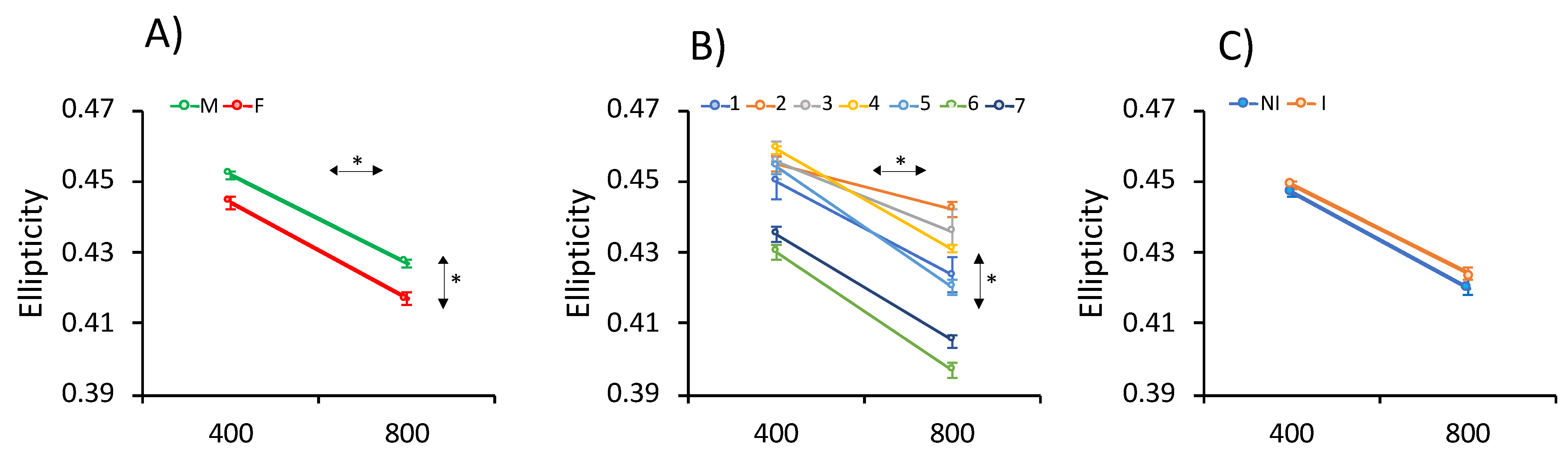
3.4. Genetic Estimates
3.4.1. Heritability
3.4.2. Genetic Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akkaynak, D.; Siemann, L.A.; Barbosa, A.; Mathger, L.M. Changeable camouflage: How well can flounder resemble the colour and spatial scale of substrates in their natural habitats? R. Soc. Open Sci. 2017, 4, 160824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, C.R.; Berendzen, P.B. Shared ecological traits influence shape of the skeleton in flatfishes (Pleuronectiformes). Peer.J. 2020, 8, e8919. [Google Scholar] [CrossRef] [Green Version]
- Reinders, M.J.; Banovi, M.; Guerrero, L.; Krystallis, A. Consumer perceptions of farmed fish: A cross-national segmentation in five European countries. Br. Food J. 2016, 118, 2581–2597. [Google Scholar] [CrossRef] [Green Version]
- de Azevedo, A.M.; Losada, A.P.; Barreiro, A.; Barreiro, J.D.; Ferreiro, I.; Riaza, A.; Vazquez, S.; Quiroga, M.I. Skeletal anomalies in reared Senegalese sole Solea senegalensis juveniles: A radiographic approach. Dis. Aquat. Organ. 2017, 124, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, I.; Ortiz-Delgado, J.B.; Darias, M.J.; Hontoria, F.; Andree, K.B.; Manchado, M.; Sarasquete, C.; Gisbert, E. Vitamin A affects flatfish development in a thyroid hormone signaling and metamorphic stage dependent manner. Front. Physiol. 2017, 8, 458. [Google Scholar] [CrossRef] [Green Version]
- de Azevedo, A.M.; Losada, A.P.; Barreiro, A.; Vazquez, S.; Quiroga, M.I. Skeletal anomalies in senegalese sole (Solea senegalensis), an anosteocytic boned flatfish species. Vet. Pathol. 2019, 56, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Dionisio, G.; Campos, C.; Valente, L.M.P.; Conceicao, L.E.C.; Cancela, M.L.; Gavaia, P.J. Effect of egg incubation temperature on the occurrence of skeletal deformities in Solea senegalensis. J. Appl. Ichthyol. 2012, 28, 471–476. [Google Scholar] [CrossRef]
- Losada, A.P.; de Azevedo, A.M.; Barreiro, A.; Barreiro, J.D.; Ferreiro, I.; Riaza, A.; Quiroga, M.I.; Vazquez, S. Skeletal malformations in Senegalese sole (Solea senegalensis Kaup, 1858): Gross morphology and radiographic correlation. J. Appl. Ichthyol. 2014, 30, 804–808. [Google Scholar] [CrossRef]
- Gavaia, P.; Domingues, S.; Engrola, S.; Drake, P.; Sarasquete, C.; Dinis, M.T.; Cancela, M.L. Comparing skeletal development of wild and hatchery-reared Senegalese sole (Solea senegalensis, Kaup 1858): Evaluation in larval and postlarval stages. Aquac. Res. 2009, 40, 1585–1593. [Google Scholar] [CrossRef]
- Fernández, I.; Pimentel, M.S.; Ortiz Delgado, J.B.; Hontoria, F.; Sarasquete, C.; Estévez, A.; Zambonino Infante, J.L.; Gisbert, E. Effect of dietary vitamin A on Senegalese sole (Solea senegalensis) skeletogenesis and larval quality. Aquaculture 2009, 295, 250–265. [Google Scholar] [CrossRef] [Green Version]
- Ambrosio, P.P.; Costa, C.; Sánchez, P.; Flos, R. Stocking density and its influence on shape of Senegalese sole adults. Aquac. Int. 2008, 16, 333–343. [Google Scholar] [CrossRef]
- Blonk, R.J.; Komen, H.; Tenghe, A.; Kamstra, A.; van Arendonk, J.A.M. Heritability of shape in common sole, Solea solea, estimated from image analysis data. Aquaculture 2010, 307, 6–11. [Google Scholar] [CrossRef]
- Guerrero-Cozar, I.; Jimenez-Fernandez, E.; Berbel, C.; Cordoba-Caballero, J.; Claros, M.G.; Zerolo, R.; Manchado, M. Genetic parameter estimates and identification of SNPs associated with growth traits in Senegalese sole. Aquaculture 2021, 539, 736665. [Google Scholar] [CrossRef]
- Martin, I.; Rasines, I.; Gomez, M.; Rodriguez, C.; Martinez, P.; Chereguini, O. Evolution of egg production and parental contribution in Senegalese sole, Solea senegalensis, during four consecutive spawning seasons. Aquaculture 2014, 424-425, 45–52. [Google Scholar] [CrossRef]
- Fatsini, E.; Gonzalez, W.; Ibarra-Zatarain, Z.; Napuchi, J.; Duncan, N. The presence of wild Senegalese sole breeders improves courtship and reproductive success in cultured conspecifics. Aquaculture 2020, 519, 734922. [Google Scholar] [CrossRef]
- Cañavate, J.P.; Fernandez-Diaz, C. Influence of co-feeding larvae with live and inert diets on weaning the sole Solea senegalensis onto commercial dry feeds. Aquaculture 1999, 174, 255–263. [Google Scholar] [CrossRef]
- Roman-Padilla, J.; Rodríguez-Rua, A.; Ponce, M.; Manchado, M.; Hachero-Cruzado, I. Effects of dietary lipid profile on larval performance and lipid management in Senegalese sole. Aquaculture 2017, 468, 80–93. [Google Scholar] [CrossRef]
- Carballo, C.; Berbel, C.; Guerrero-Cozar, I.; Jimenez-Fernandez, E.; Cousin, X.; Bégout, M.L.; Manchado, M. Evaluation of different tags on survival, growth and stress response in the flatfish Senegalese sole. Aquaculture 2018, 494, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Navarro, A.; Lee-Montero, I.; Santana, D.; Henríquez, P.; Ferrer, M.A.; Morales, A.; Soula, M.; Badilla, R.; Negrin-Baez, D.; Zamorano, M.J.; et al. IMAFISH_ML: A fully-automated image analysis software for assessing fish morphometric traits on gilthead seabream (Sparus aurata L.), meagre (Argyrosomus regius) and red porgy (Pagrus pagrus). Comput. Electron. Agr. 2016, 121, 66–73. [Google Scholar] [CrossRef]
- Guerrero-Cozar, I.; Perez-Garcia, C.; Benzekri, H.; Sánchez, J.; Seoane, P.; Cruz, F.; Gut, M.; Zamorano, M.J.; Claros, M.G.; Manchado, M. Development of whole-genome multiplex assays and construction of an integrated genetic map using SSR markers in Senegalese sole. Sci. Rep. 2020, 10, 21905. [Google Scholar] [CrossRef]
- Vandeputte, M.; Mauger, S.; Dupont-Nivet, M. An evaluation of allowing for mismatches as a way to manage genotyping errors in parentage assignment by exclusion. Mol. Ecol. Notes 2006, 6, 265–267. [Google Scholar] [CrossRef]
- Meyer, K. WOMBAT: A tool for mixed model analyses in quantitative genetics by restricted maximum likelihood (REML). J. Zhejiang Univ. Sci. B. 2007, 8, 815–821. [Google Scholar] [CrossRef] [Green Version]
- Martin, I.; Carazo, I.; Rasines, I.; Rodriguez, C.; Fernandez, R.; Martinez, P.; Norambuena, P.; Chereguini, O.; Duncan, N. Reproductive performance of captive Senegalese sole, Solea senegalensis, according to the origin (wild or cultured) and gender. Span. J. Agric. Res. 2019, 17, 2171–9292. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, P.; Ambrosio, P.P.; Flos, R. Stocking density and sex influence individual growth of Senegalese sole (Solea senegalensis). Aquaculture 2010, 300, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Constenla, M.; Padros, F. Histopathological and ultrastructural studies on a novel pathological condition in Solea senegalensis. Dis. Aquat. Organ. 2010, 90, 191–196. [Google Scholar] [CrossRef]
- Cadrin, S.X.; Silva, V.M. Morphometric variation of yellowtail flounder. ICES J. Mar. Sci. 2005, 62, 683–694. [Google Scholar] [CrossRef]
- Shikano, T. Quantitative genetic parameters for growth-related and morphometric traits of hatchery-produced Japanese founder Paralichthys olivaceus in the wild. Aquac. Res. 2007, 38, 1248–1253. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, B.; Yang, Y.; Chen, S. Estimation of genetic parameters for juvenile growth performance traits in olive flounder (Paralichthys olivaceus). Aquac. Fish 2019, 4, 48–52. [Google Scholar] [CrossRef]
- Liu, Y.-X.; Wang, G.-X.; Wang, Y.-F.; Si, F.; Sun, Z.-H.; Zhang, X.-Y.; Wang, J.-D.; Liu, H.-J. Estimation of genetic parameters for growth traits of Japanese flounder Paralichthys olivaceus using an animal model. Fish Sci. 2011, 77, 87–93. [Google Scholar] [CrossRef]
- Liu, F.; Chen, S.L.; Wang, L.; Zhang, Y.P.; Tian, Y.S.; Chen, H.L. Estimation of genetic parameters for growth related traits at different stages of development in Paralichthys olivaceus (Temminck & Schlegel, 1846). Indian J. Fish 2016, 63, 70–75. [Google Scholar]
- Blonk, R.J.; Komen, H.; Kamstra, A.; van Arendonk, J.A. Estimating breeding values with molecular relatedness and reconstructed pedigrees in natural mating populations of common sole, Solea solea. Genetics 2010, 184, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imsland, A.K.; Foss, A.; Conceiçao, L.E.C.; Dinis, M.T.; Delbare, D.; Schram, E.; Kamstra, A.; Rema, P.; White, P. A review of the culture potential of Solea solea and S. senegalensis. Rev. Fish Biol. Fish 2003, 13, 379–407. [Google Scholar] [CrossRef]
- Salas-Leiton, E.; Anguis, V.; Martin-Antonio, B.; Crespo, D.; Planas, J.V.; Infante, C.; Canavate, J.P.; Manchado, M. Effects of stocking density and feed ration on growth and gene expression in the Senegalese sole (Solea senegalensis): Potential effects on the immune response. Fish Shellfish Immunol. 2010, 28, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Salas-Leiton, E.; Anguis, V.; Rodriguez-Rua, A.; Cañavate, J.P. Stocking homogeneous size groups does not improve growth performance of Senegalese sole (Solea senegalensis, Kaup 1858) juveniles: Individual growth related to fish size. Aquac. Eng. 2010, 43, 108–113. [Google Scholar] [CrossRef]
- Mas-Muñoz, J.; Blonk, R.J.; Schrama, J.W.; van Arendonk, J.; Komen, H. Genotype by environment interaction for growth of sole (Solea solea) reared in an intensive aquaculture system and in a semi-natural environment. Aquaculture 2013, 410, 230–235. [Google Scholar] [CrossRef]
- Dupont-Nivet, M.; Chevassus, B.; Mauger, S.; Haffray, P.; Vandeputte, M. Side effects of sexual maturation on heritability estimates in rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2010, 41, e878–e880. [Google Scholar] [CrossRef]
- Omasaki, S.K.; Charo-Karisa, H.; Kahi, A.K.; Komen, H. Genotype by environment interaction for harvest weight, growth rateand shape between monosex and mixed sex Nile tilapia (Oreochromis niloticus). Aquaculture 2016, 458, 75–81. [Google Scholar] [CrossRef]
- Mengistu, S.B.; Mulder, H.A.; Benzie, J.A.H.; Khaw, H.L.; Megens, H.; Trinh, T.Q.; Komen, H. Genotype by environment interaction between aerated and non-aerated ponds and the impact of aeration on genetic parameters in Nile tilapia (Oreochromis niloticus). Aquaculture 2020, 529, 735704. [Google Scholar] [CrossRef]
- Assumpção, L.; Makrakis, M.C.; Makrakis, S.; Wagner, R.; Silva, P.S.; Lima, A.F.; Kashiwaqui, E.A.L. The use of morphometric analysis to predict the swimming efficiency of two Neotropical long-distance migratory species in fish passage. Neotrop. Ichthyol. 2012, 10, 797–804. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.; Hogan, J.D. Morphological predictors of swimming speed: A case study of pre-settlement juvenile coral reef fishes. J. Exp. Biol. 2007, 210, 2436–2443. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 400 days (n = 1840) | Male (n = 1007) | Female (n = 833) | Mean |
|---|---|---|---|
| Weight | 30.7 ± 28.0 | 34.4 ± 30.4 | 32.4 ± 29.2 |
| SL | 11.85 ± 2.87 | 12.18 ± 2.86 | 12.00 ± 2.87 |
| BHP | 3.84 ± 1.00 | 4.00 ± 1.02 | 3.91 ± 1.01 |
| BMH | 4.54 ± 1.33 | 4.71 ± 1.34 | 4.62 ± 1.34 |
| CPH | 1.13 ± 0.34 | 1.10 ± 0.34 | 1.11 ± 0.34 |
| BMH/BHP | 0.45 ± 0.03 | 0.45 ± 0.03 | 0.45 ± 0.03 |
| BMH/CPH | 1.17 ± 0.06 | 1.17 ± 0.06 | 1.17 ± 0.05 |
| Ellipticity | 4.165 ± 0.358 | 4.183 ± 0.335 | 4.173 ± 0.347 |
| 800 days (n = 1840) | Male (n = 1007) | Female (n = 833) | Mean |
| Weight | 244.0 ± 153.0 | 290.3 ± 189.3 | 264.9 ± 171.9 |
| SL | 22.91 ± 4.64 | 23.88 ± 4.91 | 23.35 ± 4.79 |
| BHP | 7.51 ± 1.66 | 8.01 ± 1.85 | 7.74 ± 1.77 |
| BMH | 9.24 ± 2.22 | 9.87 ± 2.46 | 9.53 ± 2.35 |
| CPH | 2.55 ± 0.66 | 2.68 ± 0.67 | 2.61 ± 0.67 |
| BMH/BHP | 0.426 ± 0.027 | 0.419 ± 0.028 | 0.424 ± 0.028 |
| BMH/CPH | 1.225 ± 0.054 | 1.227 ± 0.050 | 1.226 ± 0.052 |
| Ellipticity | 3.650 ± 0.325 | 3.699 ± 0.320 | 3.673 ± 0.323 |
| 400 d | W | SL | BHP | BMH | CPH | BMH/BHP | BMH/CPH | E |
|---|---|---|---|---|---|---|---|---|
| W | 0.625 ± 0.109 | 0.991 ± 0.004 | 0.988 ± 0.004 | 0.992 ± 0.003 | 0.990 ± 0.005 | 0.874 ± 0.057 | 0.161 ± 0.189 | −0.768 ± 0.073 |
| SL | 0.983 ± 0.002 | 0.567 ± 0.104 | 0.981 ± 0.007 | 0.986 ± 0.005 | 0.984 ± 0.007 | 0.881 ± 0.054 | 0.167 ± 0.189 | −0.724 ± 0.085 |
| BHP | 0.981 ± 0.002 | 0.976 ± 0.002 | 0.623 ± 0.110 | 0.991 ± 0.001 | 0.948 ± 0.005 | 0.858 ± 0.064 | 0.284 ± 0.180 | −0.738 ± 0.025 |
| BMH | 0.988 ± 0.001 | 0.982 ± 0.002 | 0.999 ± 0.001 | 0.621 ± 0.109 | 0.955 ± 0.004 | 0.878 ± 0.055 | 0.247 ± 0.183 | −0.828 ± 0.056 |
| CPH | 0.952 ± 0.004 | 0.953 ± 0.004 | 0.974 ± 0.010 | 0.979 ± 0.008 | 0.576 ± 0.105 | 0.881 ± 0.055 | 0.044 ± 0.193 | −0.749 ± 0.079 |
| BMH/BHP | 0.591 ± 0.024 | 0.61 ± 0.023 | 0.528 ± 0.028 | 0.622 ± 0.023 | 0.587 ± 0.024 | 0.270 ± 0.069 | 0.102 ± 0.035 | −0.521 ± 0.034 |
| BMH/CPH | 0.062 ± 0.045 | 0.047 ± 0.043 | 0.09 ± 0.044 | 0.094 ± 0.044 | −0.191 ± 0.044 | 0.076 ± 0.204 | 0.303 ± 0.076 | −0.254 ± 0.045 |
| E | −0.673 ± 0.031 | −0.284 ± 0.033 | −0.838 ± 0.054 | −0.750 ± 0.025 | −0.677 ± 0.031 | −0.662 ± 0.115 | −0.487 ± 0.153 | 0.774 ± 0.117 |
| 800d | W | SL | BHP | BMH | CPH | BMH/BHP | BMH/CPH | E |
| W | 0.486 ± 0.099 | 0.983 ± 0.007 | 0.974 ± 0.01 | 0.978 ± 0.008 | 0.983 ± 0.008 | 0.546 ± 0.162 | 0.016 ± 0.198 | −0.608 ± 0.115 |
| SL | 0.975 ± 0.002 | 0.433 ± 0.094 | 0.961 ± 0.015 | 0.957 ± 0.016 | 0.964 ± 0.014 | 0.412 ± 0.183 | 0.011 ± 0.198 | −0.509 ± 0.137 |
| BHP | 0.967 ± 0.003 | 0.948 ± 0.005 | 0.549 ± 0.105 | 0.996 ± 0.002 | 0.953 ± 0.018 | 0.506 ± 0.177 | 0.181 ± 0.193 | −0.703 ± 0.092 |
| BMH | 0.982 ± 0.002 | 0.962 ± 0.004 | 0.983 ± 0.001 | 0.515 ± 0.102 | 0.961 ± 0.016 | 0.586 ± 0.155 | 0.182 ± 0.192 | −0.733 ± 0.085 |
| CPH | 0.928 ± 0.005 | 0.918 ± 0.006 | 0.911 ± 0.007 | 0.926 ± 0.006 | 0.463 ± 0.097 | 0.612 ± 0.151 | −0.073 ± 0.195 | −0.586 ± 0.123 |
| BMH/BHP | 0.479 ± 0.027 | 0.486 ± 0.027 | 0.345 ± 0.031 | 0.508 ± 0.026 | 0.474 ± 0.026 | 0.144 ± 0.046 | 0.178 ± 0.211 | −0.389 ± 0.031 |
| BMH/CPH | −0.003 ± 0.042 | −0.015 ± 0.04 | 0.046 ± 0.043 | 0.050 ± 0.043 | −0.314 ± 0.040 | 0.048 ± 0.029 | 0.306 ± 0.075 | −0.217 ± 0.044 |
| E | −0.557 ± 0.037 | −0.447 ± 0.042 | −0.644 ± 0.033 | −0.662 ± 0.030 | −0.548 ± 0.038 | −0.719 ± 0.117 | −0.534 ± 0.144 | 0.735 ± 0.115 |
| Genetic | 800 d | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| W | SL | BHP | BMH | CPH | BMH/BHP | BMH/CPH | E | ||
| 400 d | W | 0.843 ± 0.054 | 0.831 ± 0.060 | 0.832 ± 0.057 | 0.838 ± 0.055 | 0.849 ± 0.054 | 0.509 ± 0.169 | 0.041 ± 0.194 | −0.554 ± 0.122 |
| SL | 0.828 ± 0.058 | 0.837 ± 0.057 | 0.813 ± 0.062 | 0.817 ± 0.061 | 0.826 ± 0.057 | 0.509 ± 0.161 | 0.049 ± 0.194 | −0.511 ± 0.134 | |
| BHP | 0.868 ± 0.047 | 0.859 ± 0.052 | 0.874 ± 0.045 | 0.876 ± 0.044 | 0.853 ± 0.053 | 0.524 ± 0.166 | 0.213 ± 0.186 | −0.637 ± 0.105 | |
| BMH | 0.853 ± 0.051 | 0.856 ± 0.050 | 0.862 ± 0.048 | 0.870 ± 0.046 | 0.846 ± 0.055 | 0.542 ± 0.162 | 0.180 ± 0.188 | −0.618 ± 0.109 | |
| CPH | 0.814 ± 0.063 | 0.821 ± 0.062 | 0.786 ± 0.070 | 0.811 ± 0.063 | 0.825 ± 0.062 | 0.523 ± 0.167 | 0.025 ± 0.195 | −0.533 ± 0.126 | |
| BMH/BHP | 0.633 ± 0.123 | 0.604 ± 0.133 | 0.575 ± 0.141 | 0.653 ± 0.121 | 0.668 ± 0.120 | 0.687 ± 0.139 | 0.038 ± 0.206 | −0.541 ± 0.141 | |
| BMH/CPH | 0.296 ± 0.184 | 0.275 ± 0.186 | 0.408 ± 0.167 | 0.372 ± 0.172 | 0.194 ± 0.193 | −0.009 ± 0.229 | 0.663 ± 0.140 | −0.442 ± 0.161 | |
| E | −0.762 ± 0.079 | −0.712 ± 0.095 | −0.849 ± 0.054 | −0.858 ± 0.051 | −0.733 ± 0.088 | −0.601 ± 0.151 | −0.492 ± 0.152 | 0.912 ± 0.032 | |
| Phenotypic | W | SL | BHP | BMH | CPH | BMH/BHP | BMH/CPH | E | |
| 400 d | W | 0.786 ± 0.018 | 0.765 ± 0.019 | 0.778 ± 0.021 | 0.783 ± 0.019 | 0.740 ± 0.021 | 0.337 ± 0.031 | 0.015 ± 0.044 | −0.474 ± 0.046 |
| SL | 0.791 ± 0.018 | 0.790 ± 0.017 | 0.781 ± 0.020 | 0.787 ± 0.019 | 0.746 ± 0.020 | 0.358 ± 0.028 | 0.013 ± 0.039 | −0.430 ± 0.046 | |
| BHP | 0.798 ± 0.017 | 0.777 ± 0.019 | 0.809 ± 0.017 | 0.809 ± 0.016 | 0.752 ± 0.020 | 0.328 ± 0.032 | 0.059 ± 0.044 | −0.536 ± 0.041 | |
| BMH | 0.798 ± 0.018 | 0.796 ± 0.017 | 0.803 ± 0.018 | 0.810 ± 0.017 | 0.751 ± 0.020 | 0.354 ± 0.031 | 0.056 ± 0.044 | −0.533 ± 0.042 | |
| CPH | 0.761 ± 0.020 | 0.765 ± 0.020 | 0.748 ± 0.022 | 0.764 ± 0.020 | 0.739 ± 0.021 | 0.336 ± 0.031 | −0.036 ± 0.042 | −0.464 ± 0.045 | |
| BMH/BHP | 0.478 ± 0.030 | 0.474 ± 0.027 | 0.313 ± 0.031 | 0.496 ± 0.029 | 0.458 ± 0.030 | 0.385 ± 0.026 | 0.032 ± 0.035 | −0.362 ± 0.038 | |
| BMH/CPH | 0.081 ± 0.043 | 0.072 ± 0.041 | 0.130 ± 0.044 | 0.118 ± 0.043 | −0.001 ± 0.042 | 0.005 ± 0.031 | 0.306 ± 0.031 | −0.197 ± 0.045 | |
| E | −0.615 ± 0.035 | −0.521 ± 0.037 | −0.672 ± 0.029 | 0.001 ± 0.028 | −0.574 ± 0.035 | −0.274 ± 0.036 | −0.175 ± 0.047 | 0.797 ± 0.021 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrero-Cozar, I.; Jimenez-Fernandez, E.; Berbel, C.; Espinosa, E.; Claros, M.G.; Zerolo, R.; Manchado, M. Genetic Estimates for Growth and Shape-Related Traits in the Flatfish Senegalese Sole. Animals 2021, 11, 1206. https://doi.org/10.3390/ani11051206
Guerrero-Cozar I, Jimenez-Fernandez E, Berbel C, Espinosa E, Claros MG, Zerolo R, Manchado M. Genetic Estimates for Growth and Shape-Related Traits in the Flatfish Senegalese Sole. Animals. 2021; 11(5):1206. https://doi.org/10.3390/ani11051206
Chicago/Turabian StyleGuerrero-Cozar, Israel, Eduardo Jimenez-Fernandez, Concha Berbel, Elena Espinosa, Manuel Gonzalo Claros, Ricardo Zerolo, and Manuel Manchado. 2021. "Genetic Estimates for Growth and Shape-Related Traits in the Flatfish Senegalese Sole" Animals 11, no. 5: 1206. https://doi.org/10.3390/ani11051206
APA StyleGuerrero-Cozar, I., Jimenez-Fernandez, E., Berbel, C., Espinosa, E., Claros, M. G., Zerolo, R., & Manchado, M. (2021). Genetic Estimates for Growth and Shape-Related Traits in the Flatfish Senegalese Sole. Animals, 11(5), 1206. https://doi.org/10.3390/ani11051206