
Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea

 ,
,  ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
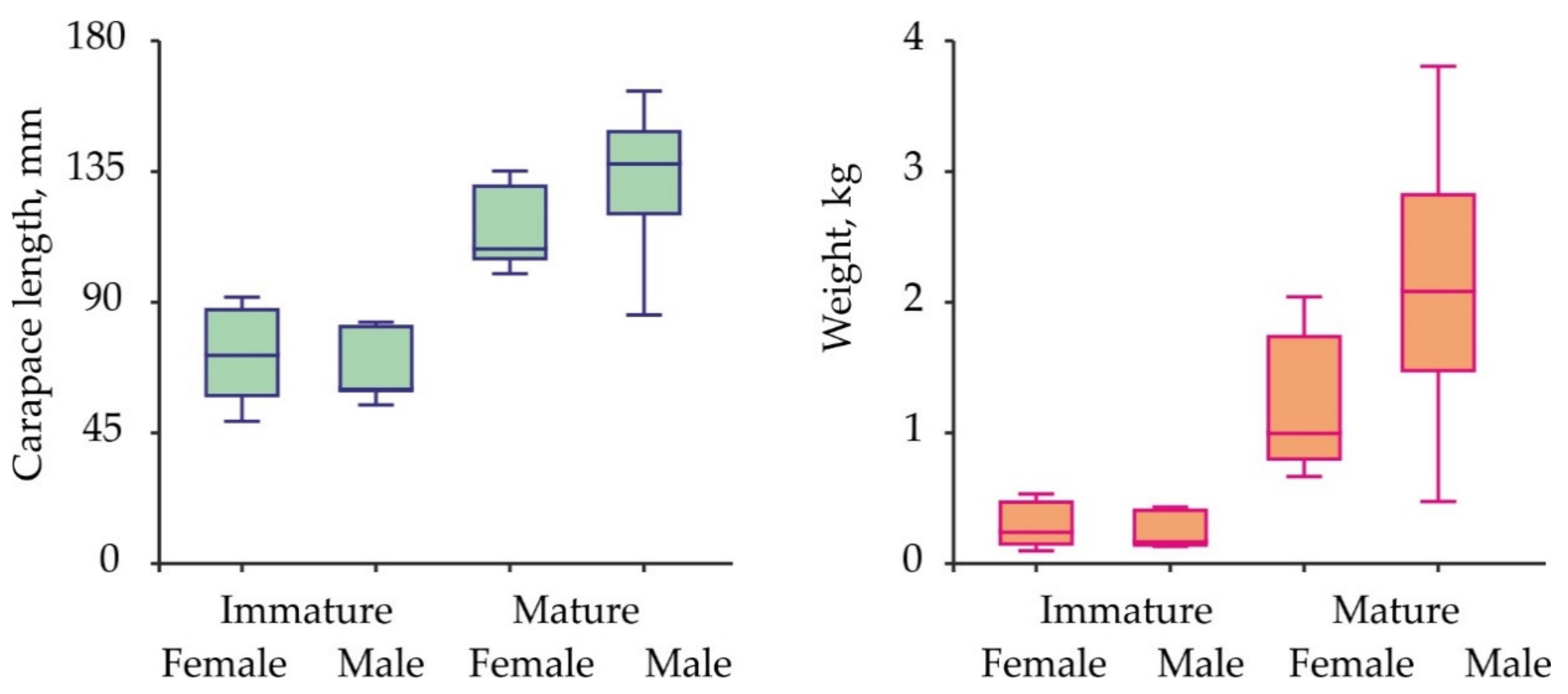
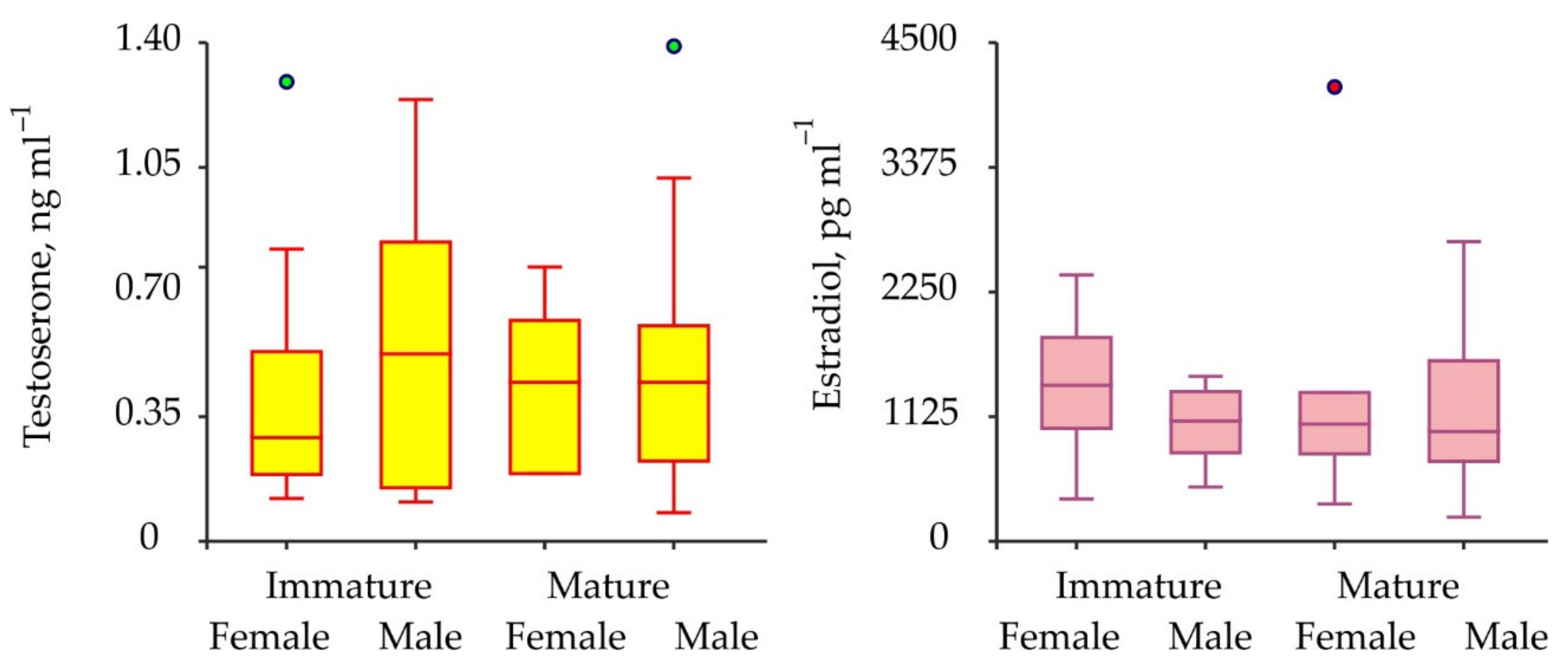
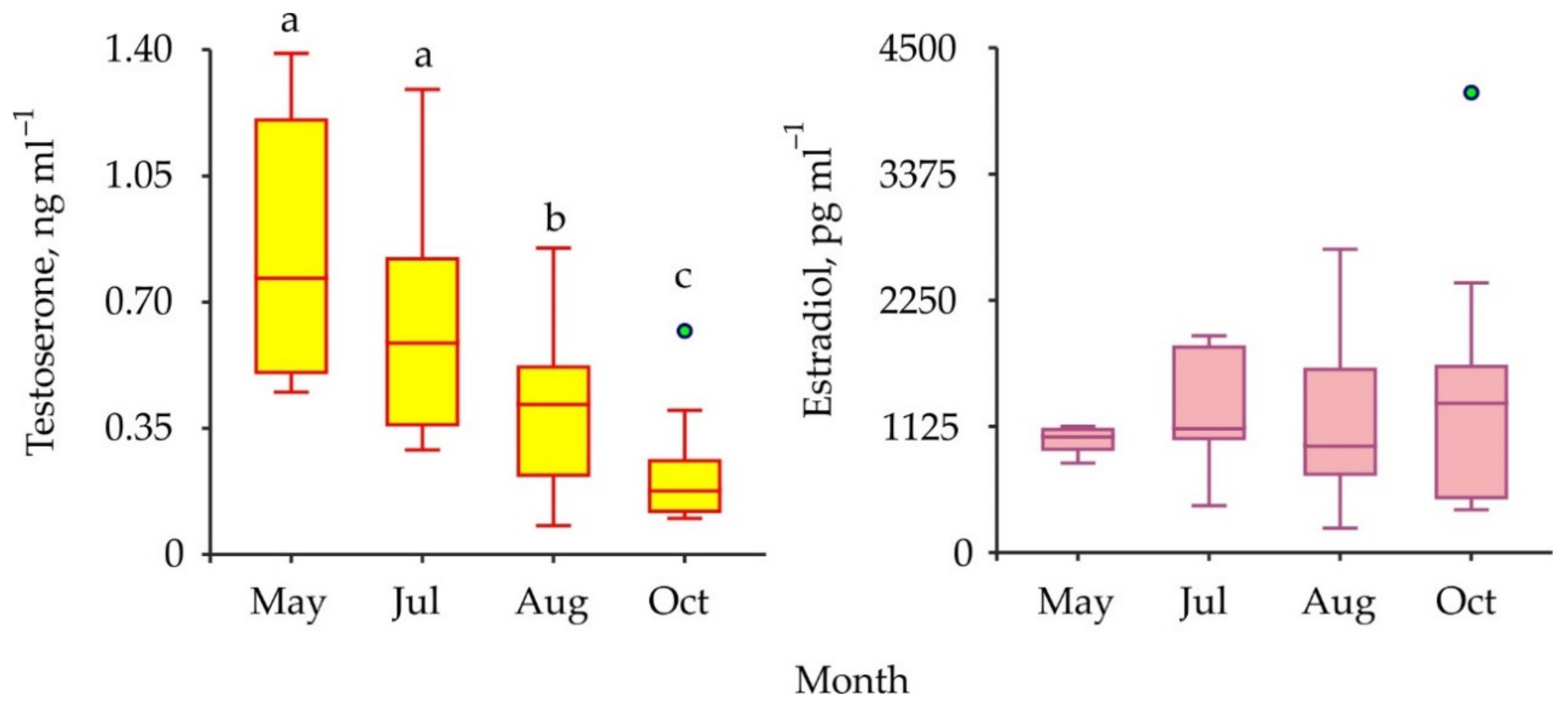
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2018, 28, 331–353. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition of the Barents Sea red king crab (Paralithodes camtschaticus) leg meat. J. Food Compos. Anal. 2021, 98, 103826. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef]
- Kuzmin, S.A.; Gudimova, E.N. Introduction of the Kamchatka (Red King) Crab in the Barents Sea. Pecularities of Biology, Perspectives of Fishery; Publishing Kola Science Centre Russian Academy of Sciences: Apatity, Russia, 2002. (In Russian) [Google Scholar]
- Pavlova, L.V. Red king crab trophic relations and its influence on bottom biocenoses. In Biology and Physiology of the Red King Crab from the Coastal Zone of the Barents Sea; Matishov, G.G., Ed.; Publishing Kola Scientific Centre Russian Academy of Sciences: Apatity, Russia, 2008; pp. 77–104. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population dynamics of the invasive lithodid crab, Paralithodes camtschaticus, in a typical bay of the Barents Sea. ICES J. Mar. Sci. 2013, 70, 1255–1262. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size-at-age of juvenile red king crab (Paralithodes camtschaticus) in the coastal Barents Sea. Cah. Biol. Mar. 2014, 55, 43–48. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size at maturity of female red king crab, Paralithodes camtschaticus, from the costal zone of Kola Peninsula (southern Barents Sea). Cah. Biol. Mar. 2015, 56, 49–54. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Hemolymph molting hormone concentrations in red king crabs from the Barents Sea. Polar Biol. 2010, 33, 1293–1298. [Google Scholar] [CrossRef]
- Moiseeva, S.A.; Moiseev, S.I. Relation between muscular tissue condition in limbs and hemocyanin concentration in the hemolymph of red king crab (Paralithodes camtschaticus) in the Barents Sea. Probl. Fish. 2008, 9, 200–217, (In Russian with English Abstract). [Google Scholar]
- Kutzler, M.A. Possible relationship between long-term adverse health effects of gonad-removing surgical sterilization and luteinizing hormone in dogs. Animals 2020, 10, 599. [Google Scholar] [CrossRef] [Green Version]
- Am-in, N.; Suwimonteerabutr, J.; Kirkwood, R.N. Serum anti-mullerian hormone and estradiol concentrations in gilts and their age at puberty. Animals 2020, 10, 2189. [Google Scholar] [CrossRef] [PubMed]
- Hyndman, T.H.; Algar, K.L.; Woodward, A.P.; Coiacetto, F.; Hampton, J.O.; Nickels, D.; Hamilton, N.; Barnes, A.; Algar, D. Estradiol-17β pharmacokinetics and histological assessment of the ovaries and uterine horns following intramuscular administration of estradiol cypionate in feral cats. Animals 2020, 10, 1708. [Google Scholar] [CrossRef]
- Massoud, D.; Lao-Pérez, M.; Ortega, E.; Burgos, M.; Jiménez, R.; Barrionuevo, F.J. Divergent seasonal reproductive patterns in syntopic populations of two murine species in southern Spain, Mus spretus and Apodemus sylvaticus. Animals 2021, 11, 243. [Google Scholar] [CrossRef]
- García, M.-L.; Muelas, R.; Argente, M.-J.; Peiró, R. Relationship between prenatal characteristics and body condition and endocrine profile in rabbits. Animals 2021, 11, 95. [Google Scholar] [CrossRef]
- Verslycke, T.; De Wasch, K.; De Brabander, H.F.; Janssen, C.R. Testosterone metabolism in the estuarine mysid Neomysis integer (Crustacea; Mysidacea): Identification of testosterone metabolites and endogenous vertebrate-type steroids. Gen. Comp. Endocrinol. 2002, 126, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janer, G.; Porte, C. Sex steroids and potential mechanisms of non-genomic endocrine disruption in invertebrates. Ecotoxicology 2007, 16, 145–160. [Google Scholar] [CrossRef]
- Swevers, L.; Lambert, J.G.; De Loof, A. Metabolism of vertebrate-type steroids by tissues of three crustacean species. Comp. Biochem. Physiol. 1991, 99B, 35–41. [Google Scholar] [CrossRef]
- Blanchet, M.-F.; Ozon, R.; Meusy, J.J. Metabolism of steroids, in vitro, in the male crab Carcinus maenas Linne. Comp. Biochem. Physiol. 1972, 41B, 251–261. [Google Scholar] [CrossRef]
- Shih, J.T.; Liao, C.F. Conversion of cholesterol to sex steroid-like substances by tissues of Mictyris brevidactylus in vitro. Zool. Stud. 1998, 37, 102–110. [Google Scholar]
- Charmantier, G.; Charmantier-Daures, M.; Van Herp, F. Hormonal regulation of growth and reproduction in crustaceans. In Recent Advances in Marine Biotechnology; Fingerman, M., Nagabhushanam, R., Eds.; Oxford & IBH: New Delhi, India, 1997; Volume 1, pp. 109–161. [Google Scholar]
- Swetha, C.H.; Sainath, S.B.; Reddy, P.R.; Reddy, P.S. Reproductive Endocrinology of Female Crustaceans: Perspective and Prospective. J. Mar. Sci. Res. Dev. 2011, 3, 1–13. [Google Scholar]
- Quinitio, E.T.; Yamauchi, K.; Hara, A.; Fuji, A. Profiles of progesterone-and estradiol-like substances in the hemolymph of female Pandalus kessleri during an annual reproductive cycle. Gen. Comp. Endocrinol. 1991, 81, 343–348. [Google Scholar] [CrossRef]
- Gunamalai, V.; Kirubagaran, R.; Subramoniam, T. Vertebrate steroids and the control of female reproduction in two decapods, Emertia asiatica and Macrobrachium rosenbergii. Curr. Sci. 2006, 90, 119–123. [Google Scholar]
- Huiyang, H.; Haihui, Y.; Shizhao, H.; Guizhong, W. Profiles of gonadotropins and steroid hormone-like substances in the hemolymph of mud crab Scylla paramamosain during the reproduction cycle. Mar. Freshw. Behav. Physiol. 2009, 42, 297–305. [Google Scholar] [CrossRef]
- Lewis, S.E.; Freund, J.G.; Riddell, G.E.; Wankowski, J.L.; Dick, J.T.A.; Baldridge, M.G. Interspecific comparison of estrogen and testosterone concentrations in three species of amphipods (Gammarus duebeni celticus, G. pseudolimnaeus, and G. pulex). J. Crustacean Biol. 2015, 35, 789–792. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs. Fairbanks, Alaska Sea Grant College Program; University of Alaska: Fairbanks, AK, USA, 2005. [Google Scholar]
- WoRMS. World Register of Marine Species. 2021. Available online: http://www.marinespecies.org/ (accessed on 22 February 2021).
- Fairs, N.J.; Evershed, R.P.; Quinlan, P.T.; Goad, L.J. Detection of unconjugated and conjugated steroids in the ovary, eggs, and haemolymph of the decapod crustacean Nephrops norvegicus. Gen. Comp. Endocrinol. 1989, 4, 199–208. [Google Scholar] [CrossRef]
- Warrier, S.R.; Tirumalai, R.; Subramoniam, T. Occurrence of vertebrate steroids, estradiol 17 β and progesterone in the reproducing females of the mud crab Scylla serrata. Comp. Biochem. Physiol. Part A 2001, 130, 283–294. [Google Scholar] [CrossRef]
- Wei, W.; Wei, H.; Liu, Q. Effect of estradiol in hemolymph and gonad on precociousness of Eriocheir sinensis. J. Fish. China 2005, 29, 826–865. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-SCKX200506020.htm (accessed on 20 May 2020). (In Chinese with English Abstract).
- Swetha, C.H.; Girish, B.P.; Reddy, P.S. Elucidation of the role of estradiol and progesterone in regulating reproduction in the edible crab, Oziothelphusa senex senex. RSC Adv. 2016, 6, 24959. [Google Scholar] [CrossRef]
- Shih, J.-T. Sex steroid-like substances in the ovaries, hepatopancreases, and body fluid of female Mictyris brevidactylus. Zool. Stud. 1997, 36, 136–145. [Google Scholar]
- Amin-Safwan, A.; Muhd-Farouk, H.; Mardhiyyah, M.P.; Nadirah, M.; Ikhwanuddin, M. Does water salinity affect the level of 17β-estradiol and ovarian physiology of orange mud crab, Scylla olivacea (Herbst, 1796) in captivity? J. King Saud. Univ. Sci. 2019, 31, 827–835. [Google Scholar] [CrossRef]
- Quinitio, E.T.; Hara, A.; Yamauchi, K.; Nakao, S. Changes in the steroid hormone and vitellogenin levels during the gametogenic cycle of the giant tiger shrimp, Penaeus monodon. Comp. Biochem. Physiol. 1994, 109C, 21–26. [Google Scholar] [CrossRef]
- Okumura, T.; Sakiyama, K. Hemolymph levels of vertebrate-type steroid hormones in female kuruma prawn Marsupenaeus japonicus (Crustacea: Decapoda: Penaeidae) during natural reproductive cycle and induced ovarian development by eyestalk ablation. Fish. Sci. 2004, 70, 372–380. [Google Scholar] [CrossRef]
- Burns, B.G.; Sangalang, G.B.; Freeman, H.C.; McMenemy, M. Isolation and identification of testosterone from the serum and testes of the American lobster (Homarus americanus). Gen. Comp. Endocrinol. 1984, 54, 429–432. [Google Scholar] [CrossRef]
- Huang, D.-J.; Chen, H.-C. Effects of chlordane and lindane on testosterone and vitellogenin levels in green neon shrimp (Neocaridina denticulata). Int. J. Toxicol. 2004, 23, 91–95. [Google Scholar] [CrossRef]
- Tuck, I.D.; Atkinson, R.J.A.; Chapman, C.J. Population biology of the Norway lobster, Nephrops norvegicus (L.) in the Firth of Clyde, Scotland II: Fecundity and size at onset of sexual maturity. ICES J. Mar. Sci. 2000, 57, 1227–1239. [Google Scholar] [CrossRef]
- Prasad, P.N.; Neelakantan, B. Maturity and breeding of the mud crab, Scylla serrata (Forskal) (Decapoda: Brachyura: Portunidae). Proc. Indian Acad. Sci. Anim. Sci. 1989, 98, 341–349. [Google Scholar] [CrossRef]
- Tanglin, Z.; Zhongjie, L.; Yibo, C. Survival, growth, sex ratio, and maturity of the Chinese mitten crab (Eriocheir sinensis) reared in a Chinese pond. J. Freshw. Ecol. 2001, 16, 633–640. [Google Scholar]
- Takeda, S. Sexual differences in behaviour during the breeding seasonin the soldier crab (Mictyris brevidactylus). J. Zool. Lond. 2005, 266, 197–204. [Google Scholar] [CrossRef]
- Subramoniam, T. Steroidal control of vitellogenesis in Crustacea: A new understanding for improving shrimp hatchery production. Proc. Indian Acad. Sci. Anim. Sci. 2017, 83, 595–610. [Google Scholar]
- Pan, J.; Liu, M.; Chen, T.; Yongxu, C.; Xugan, W. Immunolocalization and changes of 17beta-estradiol during ovarian development of Chinese mitten crab Eriocheir Sinensis. Cell Tissue Res. 2018, 373, 509–520. [Google Scholar]
- Yano, I.; Hoshino, R. Effects of 17β-estradiol on the vitellogenin synthesis and oocyte development in the ovary of kuruma prawn (Marsupenaeus japonicus). Comp. Biochem. Physiol. Part A 2006, 144, 18–23. [Google Scholar] [CrossRef]
- Pakdeenarong, N. Effect of estradiol-17β on embryonic tolerance, growth, and muscular compactness of giant freshwater prawn, Macrobrachium rosenbergii. J. Appl. Biol. Biotechnol. 2019, 7, 16–20. [Google Scholar]
- Coccia, E.; De Lisa, E.; Di Cristo, C.; Di Cosmo, A.; Paolucci, M. Effects of estradiol and progesterone on the reproduction of the freshwater crayfish Cherax albidus. Biol. Bull. 2010, 218, 36–47. [Google Scholar] [CrossRef]
- Zapata, V.; Lopez Greco, L.S.; Medesani, D.; Rodriguez, E.M. Ovarian growth in the crab, Chasmagnathus granulata induced by hormones and neuroregulators throughout the year. In vivo and in vitro studies. Aquaculture 2003, 224, 1–4. [Google Scholar] [CrossRef]
- Subramoniam, T.; Kirubagaran, R. Endocrine regulation of vitellogenesis in lobsters. J. Mar. Biol. Assoc. India 2010, 52, 229–236. [Google Scholar]
- Nagabhushanam, R.; Kulkarni, G.K. Effect of exogenous testosterone on the androgenic gland and testis of a marine penaeid prawn, Parapenaeopsis hardwickii (Miers) (Crustacea, Decapoda, Penaeidae). Aquaculture 1981, 23, 19–27. [Google Scholar] [CrossRef]
- Maheswarudu, G.; Rajkumar, U.; Sreeram, M.P.; Chakravarty, M.S.; Sajeev, C.K. Effect of testosterone hormone on performance of male broodstock of black tiger shrimp Penaeus monodon Fabricius, 1798. J. Vet. Sci. Photon 2015, 116, 446–456. [Google Scholar]
- Koskela, R.W.; Greenwood, J.G.; Rothlisberg, P.C. The influence of prostaglandin E2 and the steroid hormones, 17α-hydroxyprogesterone and 17β-estradiol on moulting and ovarian development in the tiger prawn, Penaeus esculentus Haswell, 1879 (Crustacea: Decapoda). Comp. Biochem. Physiol. Part A 1992, 101, 295–299. [Google Scholar] [CrossRef]
- Fiordelmondo, E.; Magi, G.E.; Mariotti, F.; Bakiu, R.; Roncarati, A. Improvement of the water quality in rainbow trout farming by means of the feeding type and management over 10 years (2009–2019). Animals 2020, 10, 1541. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Aquaculture of green sea urchin in the Barents Sea: A brief review of Russian studies. Rev. Aquac. 2020, 12, 1280–1290. [Google Scholar] [CrossRef]
- Samat, N.A.; Yusoff, F.M.; Rasdi, N.W.; Karim, M. Enhancement of live food nutritional status with essential nutrients for improving aquatic animal health: A review. Animals 2020, 10, 2457. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, A.G.; Dvoretsky, V.G. Does spine removal affect molting process in the king red crab (Paralithodes camtschaticus) in the Barents Sea? Aquaculture 2012, 326–329, 173–177. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hormone | Comparisons | d | H | p |
|---|---|---|---|---|
| Testosterone | M-im vs. M-mat | 1 | 0.436 | 0.509 |
| F-im vs. F-mat | 1 | 0.875 | 0.349 | |
| M-tot vs. F-tot | 1 | 0.750 | 0.386 | |
| May vs. July vs. August vs. October | 3 | 24.030 | 0.00002 | |
| 17β-estradiol | M-im vs. M-mat | 1 | 0.001 | 0.982 |
| F-im vs. F-mat | 1 | 0.698 | 0.404 | |
| M-tot vs. F-tot | 1 | 1.032 | 0.310 | |
| May vs. July vs. August vs. October | 3 | 0.950 | 0.813 |
| Steroid | Species | Sex | Matrix | Levels | Reference |
|---|---|---|---|---|---|
| E | Nephrops norvegicus | F | Eggs | 900 pg g–1 | [31] |
| E | Nephrops norvegicus | F | Hemolymph | 800 pg mL–1 | [31] |
| E | Scylla serrata | F | Hemolymph | 230–1040 pg g–1 | [32] |
| E | Scylla serrata | F | Ovary | 180–1250 pg g–1 | [32] |
| E | Scylla serrata | F | Hepatopancreas | 300–4100 pg mg–1 | [32] |
| E | Scylla olivacea | F | Hemolymph | 970–3520 pg mL–1 | [36] |
| E | Pandalus latirostris | F | Hemolymph | 18.1–54.3 pg mL–1 | [25] |
| E | Penaeus monodon | F | Hemolymph | 30.4 pg mL–1 | [37] |
| E | Penaeus monodon | F | Ovary | 30–77 pg g–1 | [37] |
| E | Penaeus monodon | F | Hepatopancreas | 20–752.6 pg mg–1 | [37] |
| E | Penaeus japonicus | F | Hemolymph | 2.8–8.7 pg mL–1 | [38] |
| E | Gammarus duebeni | F | Hemolymph | 86.6 pg mL–1 | [28] |
| E | Gammarus duebeni | M | Hemolymph | 41.6 pg mL–1 | [28] |
| E | Gammarus pulex | F | Hemolymph | 38.8 pg mL–1 | [28] |
| E | Gammarus pulex | M | Hemolymph | 47.4 pg mL–1 | [28] |
| E | Gammarus pseudolimnaeus | F | Hemolymph | 18.8 pg mL–1 | [28] |
| E | Gammarus pseudolimnaeus | M | Hemolymph | 29.3 pg mL–1 | [28] |
| E | Scylla paramamosain | F | Hemolymph | 18.5–75.4 pg mL–1 | [27] |
| E | Emertia emeritus | F | Ovary | 2000–8000 pg g–1 | [26] |
| E | Emertia emeritus | F | Hepatopancreas | 2900–9700 pg g–1 | [26] |
| E | Macrobrachium rosenbergii | F | Ovary | 67–341 pg g–1 | [26] |
| E | Macrobrachium rosenbergii | F | Hepatopancreas | 168–663 pg g–1 | [26] |
| E | Eriocheir sinensis | F | Hemolymph | 118.8–291.8 pg mL–1 | [33] |
| E | Eriocheir sinensis | F | Ovary | 232.2–257.8 pg g–1 | [33] |
| E | Eriocheir sinensis | M | Hemolymph | 122.7–237.6 pg mL–1 | [33] |
| E | Eriocheir sinensis | M | Testes | 128.5–150.0 pg g–1 | [33] |
| E | Spiralothelphusa senex | F | Hemolymph | 27.9–134.2 pg mL–1 | [34] |
| E | Mictyris brevidactylus | F | Hemolymph | 0.2–0.6 pg g–1 | [35] |
| E | Mictyris brevidactylus | F | Ovary | 8.8–70.0 pg g–1 | [35] |
| E | Mictyris brevidactylus | F | Hepatopancreas | 0.6–122.9 pg g–1 | [35] |
| T | Nephrops norvegicus | F | Ovary | 19000 pg g–1 | [31] |
| T | Homarus americanus | M | Testes | 14300 pg g–1 | [39] |
| T | Gammarus duebeni | F | Hemolymph | 25.3 pg mL–1 | [28] |
| T | Gammarus duebeni | M | Hemolymph | 15.6 pg mL–1 | [28] |
| T | Gammarus pulex | F | Hemolymph | 22.1 pg mL–1 | [28] |
| T | Gammarus pulex | M | Hemolymph | 22.7 pg mL–1 | [28] |
| T | Gammarus pseudolimnaeus | F | Hemolymph | 21.7 pg mL–1 | [28] |
| T | Gammarus pseudolimnaeus | M | Hemolymph | 31.4 pg mL–1 | [28] |
| T | Scylla paramamosain | M | Hemolymph | 394–607 pg mL–1 | [27] |
| T | Penaeus japonicus | F | Hemolymph | 2.2–4.4 pg mL–1 | [38] |
| T | Neocaridina denticulata | ND | Hemolymph | 2700 pg g–1 | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, A.G.; Tipisova, E.V.; Elfimova, A.E.; Alikina, V.A.; Dvoretsky, V.G. Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea. Animals 2021, 11, 2149. https://doi.org/10.3390/ani11072149
Dvoretsky AG, Tipisova EV, Elfimova AE, Alikina VA, Dvoretsky VG. Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea. Animals. 2021; 11(7):2149. https://doi.org/10.3390/ani11072149
Chicago/Turabian StyleDvoretsky, Alexander G., Elena V. Tipisova, Aleksandra E. Elfimova, Viktoria A. Alikina, and Vladimir G. Dvoretsky. 2021. "Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea" Animals 11, no. 7: 2149. https://doi.org/10.3390/ani11072149
APA StyleDvoretsky, A. G., Tipisova, E. V., Elfimova, A. E., Alikina, V. A., & Dvoretsky, V. G. (2021). Sex Hormones in Hemolymph of Red King Crabs from the Barents Sea. Animals, 11(7), 2149. https://doi.org/10.3390/ani11072149