
Myostatin and Related Factors Are Involved in Skeletal Muscle Protein Breakdown in Growing Broilers Exposed to Constant Heat Stress

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds and Treatments
2.2. Sampling Collection and Chemical Analysis
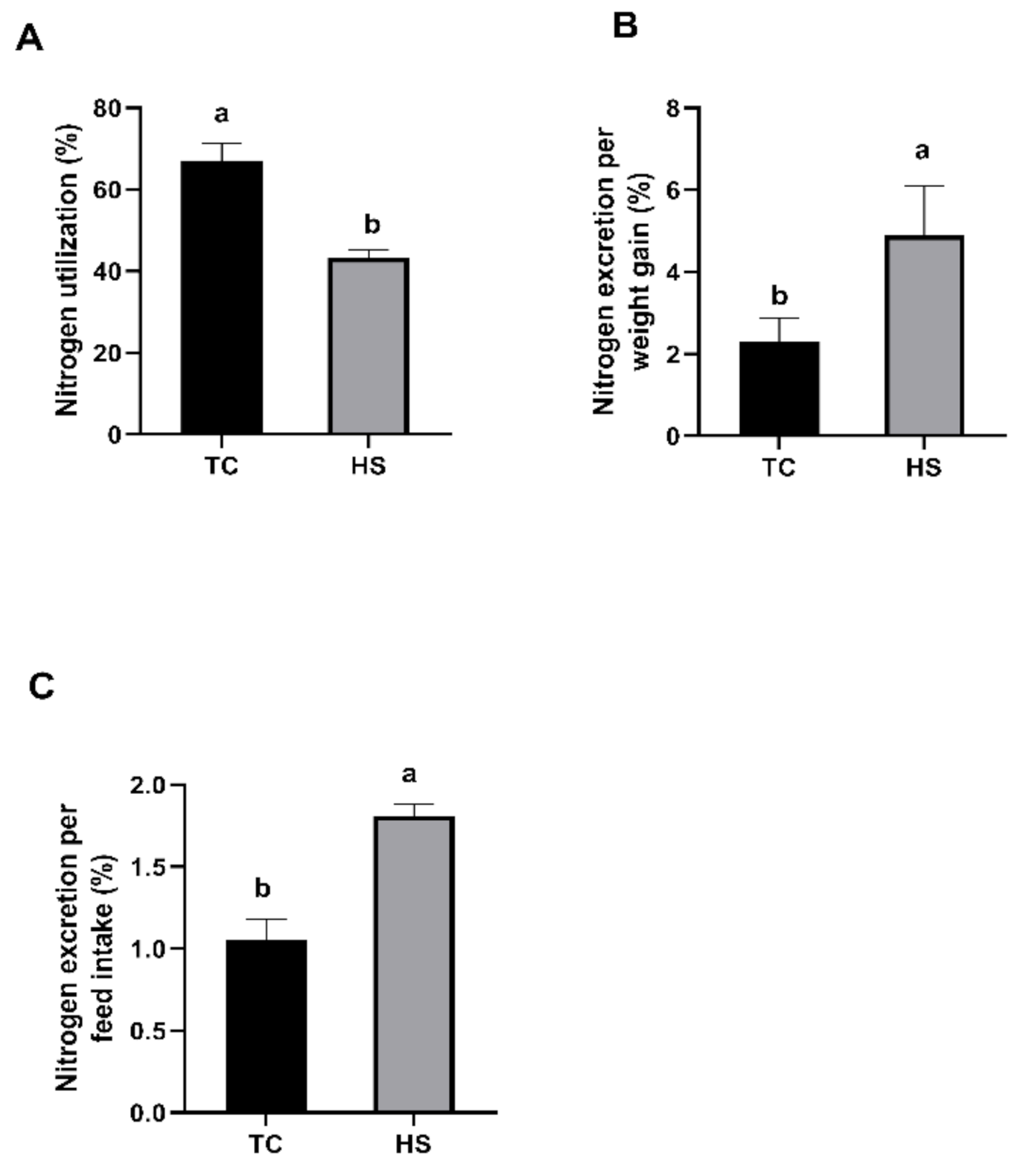
2.2.1. Nitrogen Utilization and Nitrogen Excretion
2.2.2. Blood Measurements
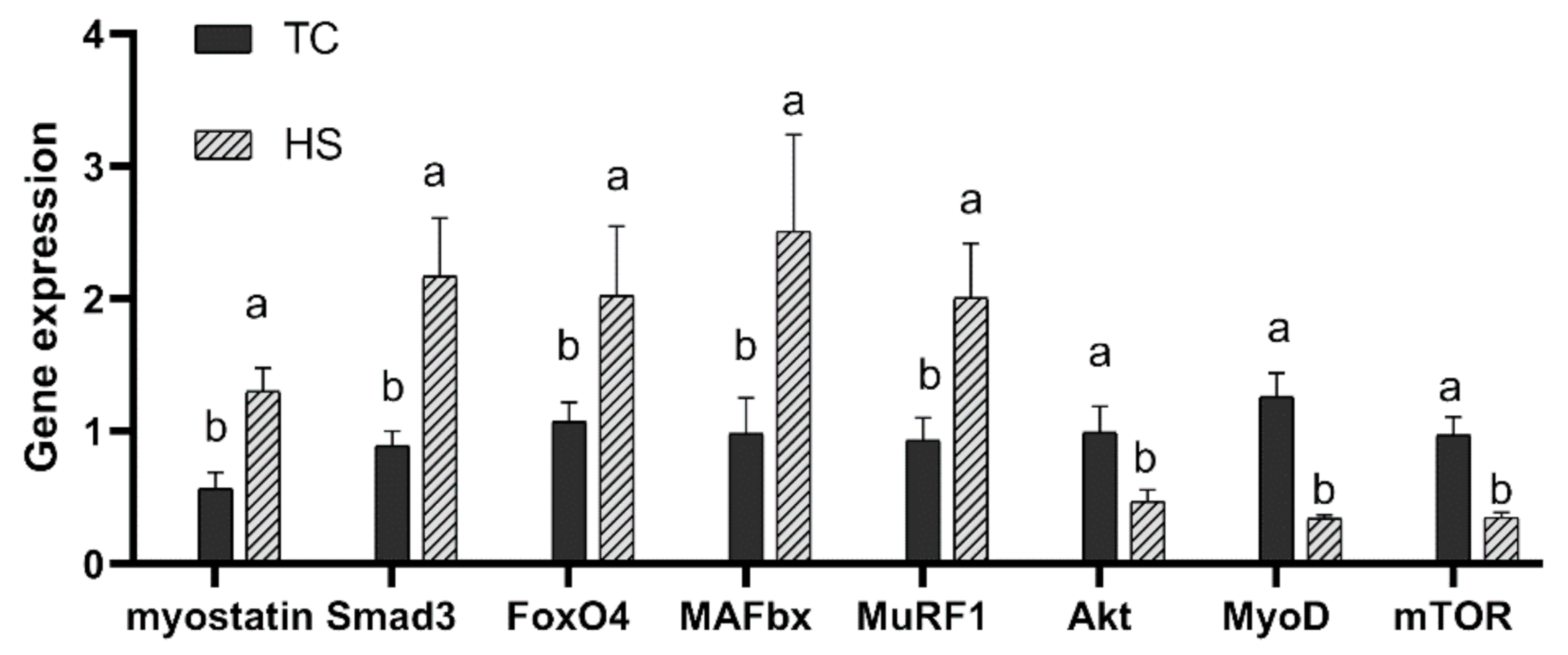
2.2.3. Regulatory Factors Gene Expression
2.3. Statistical Analysis
3. Results
3.1. Growth Performance, Breast Muscle Yield, Nitrogen Utilization and Nitrogen Excretion
3.2. Blood Biochemical Indexes
3.3. Regulatory Factors Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zuo, J.; Xu, M.; Abdullahi, Y.A.; Ma, L.; Zhang, Z.; Feng, D. Constant heat stress reduces skeletal muscle protein deposition in broilers. J. Sci. Food Agric. 2015, 95, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Orhan, C.; Tuzcu, M.; Sahin, N.; Hayirli, A.; Bilgili, S.; Kucuk, O. Lycopene activates antioxidant enzymes and nuclear transcription factor systems in heat-stressed broilers. Poult. Sci. 2016, 95, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhai, W.; Corzo, A.; Mejia, L.; Zumwalt, C.D.; Peebles, E.D. Effects of dietary amino acid density and metabolizable energy level on the growth and meat yield of summer-reared broilers. J. Appl. Poult. Res. 2014, 23, 501–515. [Google Scholar] [CrossRef]
- Awad, A.E.; Idrus, Z.; Farjam, A.S.; Bello, A. Growth performance, duodenal morphology and the caecal microbial population in female broiler chickens fed glycine-fortified low protein diets under heat stress conditions. Br. Poult. Sci. 2018, 59, 340–348. [Google Scholar] [CrossRef]
- Sifa, D.; Bai, X.; Zhang, D.; Hu, H.; Wu, X.; Wen, A.; He, S.; Zhao, L. Dietary glutamine improves meat quality, skeletal muscle antioxidant capacity and glutamine metabolism in broilers under acute heat stress. J. Appl. Anim. Res. 2018, 46, 1412–1417. [Google Scholar] [CrossRef]
- Petracci, M.; Cavani, C. Muscle Growth and Poultry Meat Quality Issues. Nutrients 2012, 4, 1–12. [Google Scholar] [CrossRef]
- Matsakas, A.; Patel, K. Skeletal muscle fibre plasticity in response to selected environmental and physiological stimuli. Histol. Histopathol. 2009, 24, 611–629. [Google Scholar]
- Yunianto, V.D.; Hayashit, K.; Kaiwda, S.; Ohtsuka, A.; Tomita, Y. Effect of environmental temperature on muscle protein turnover and heat production in tube-fed broiler chickens. Br. J. Nutr. 1997, 77, 897–909. [Google Scholar] [CrossRef]
- Dai, S.F.; Gao, F.; Zhang, W.H.; Song, S.X.; Xu, X.L.; Zhou, G.H. Effects of dietary glutamine and gamma-aminobutyric acid on performance, carcass characteristics and serum parameters in broilers under circular heat stress. Anim. Feed Sci. Technol. 2011, 168, 51–60. [Google Scholar] [CrossRef]
- Zhang, Z.; Jia, G.; Zuo, J.; Zhang, Y.; Lei, J.; Ren, L.; Feng, D. Effects of constant and cyclic heat stress on muscle metabolism and meat quality of broiler breast fillet and thigh meat. Poult. Sci. 2012, 91, 2931–2937. [Google Scholar] [CrossRef]
- Akşit, M.; Yalçin, S.; Ozkan, S.; Metin, K.; Ozdemir, D. Effects of Temperature During Rearing and Crating on Stress Parameters and Meat Quality of Broilers. Poult. Sci. 2006, 85, 1867. [Google Scholar] [CrossRef]
- Rosa, P.; Filho, D.F.; Dahlke, F.; Vieira, B.; Macari, M.; Furlan, R. Performance and carcass characteristics of broiler chickens with different growth potential and submitted to heat stress. Rev. Bras. De Ciência Avícola 2007, 9, 181–186. [Google Scholar] [CrossRef]
- Li, Y.; Li, F.; Duan, Y.; Guo, Q.; Wang, W.; Wen, C.; Huang, X.; Yin, Y. The Protein and Energy Metabolic Response of Skeletal Muscle to the Low-Protein Diets in Growing Pigs. J. Agric. Food Chem. 2017, 65, 8544–8551. [Google Scholar] [CrossRef]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Lozier, N.R.; Kopchick, J.J.; Lacalle, S.D. Relative Contributions of Myostatin and the GH/IGF-1 Axis in Body Composition and Muscle Strength. Front. Physiol. 2018, 9, 1418. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; He, X.; Lu, Z.; Lin, Z.; Li, J.; Yun, J.; Zhou, G.; Gao, F. Chronic heat stress affects muscle hypertrophy, muscle protein synthesis and uptake of amino acid in broilers via insulin like growth factor-mammalian target of rapamycin signal pathway. Poult. Sci. 2018, 97, 4150–4158. [Google Scholar] [CrossRef]
- Gomes, M.D.; Lecker, S.H.; Jagoe, R.T.; Navon, A.; Goldberg, A.L. Atrogin-1, a muscle-specific F-box protein highly expressed during muscle atrophy. Proc. Natl. Acad. Sci. USA 2001, 98, 14440–14445. [Google Scholar] [CrossRef] [Green Version]
- Rebbapragada, A.; Benchabane, H.; Wrana, J.; Celeste, A.; Attisano, L. Myostatin signals through a transforming growth factor β-like signaling pathway to block adipogenesis. Mol. Cell. Biol. 2003, 23, 7230–7242. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Mcpherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef] [Green Version]
- Trendelenburg, A.U.; Meyer, A.; Rohner, D.; Boyle, J.; Hatakeyama, S.; Glass, D.J. Myostatin reduces Akt/TORC1/p70S6K signaling, inhibiting myoblast differentiation and myotube size. Am. J. Physiol. Cell Physiol. 2009, 296, C1258–C1270. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Yang, M.; Wei, Z.; Gu, M.; Li, G. MSTN Mutant Promotes Myogenic Differentiation by Increasing Demethylase TET1 Expression via the SMAD2/SMAD3 Pathway. Int. J. Biol. Sci. 2020, 16, 1324–1334. [Google Scholar] [CrossRef] [Green Version]
- Saneyasu, T.; Honda, K.; Kamisoyama, H. Myostatin Increases Smad2 Phosphorylation and Atrogin-1 Expression in Chick Embryonic Myotubes. J. Poult. Sci. 2019, 56, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Del Vesco, A.P.; Eliane, G.; de Oliveira Grieser, D.; Zancanela, V.; Soares, M.A.M.; de Oliveira Neto, A.R. Effects of methionine supplementation on the expression of oxidative stress-related genes in acute heat stress-exposed broilers. Br. J. Nutr. 2015, 113, 549–559. [Google Scholar] [CrossRef]
- Furukawa, K.; Kikusato, M.; Kamizono, T.; Toyomizu, M. Time-course changes in muscle protein degradation in heat-stressed chickens: Possible involvement of corticosterone and mitochondrial reactive oxygen species generation in induction of the ubiquitin-proteasome system. Gen. Comp. Endocrinol. 2016, 228, 105–110. [Google Scholar] [CrossRef]
- Helrich, K. Official Methods of Analysis of the Association of Official Analytical Chemists; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zulkifili, I.; Akmal, A.F.; Soleimani, A.F.; Hossain, M.A.; Awad, E.A. Effects of low-protein diets on acute phase proteins and heat shock protein 70 responses, and growth performance in broiler chickens under heat stress condition. Poult. Sci. 2018, 97, 1306–1314. [Google Scholar] [CrossRef]
- Luo, J.; Song, J.; Liu, L.; Xue, B.; Tian, G.; Yang, Y. Effect of epigallocatechin gallate on growth performance and serum biochemical metabolites in heat-stressed broilers. Poult. Sci. 2018, 97, 599–606. [Google Scholar] [CrossRef]
- Kumar, P.; Patra, A.K.; Mandal, G.P.; Samanta, I.; Pradhan, S. Effect of black cumin seeds on growth performance, nutrient utilization, immunity, gut health and nitrogen excretion in broiler chickens. J. Sci. Food Agric. 2017, 97, 3742–3751. [Google Scholar] [CrossRef]
- Hadad, Y.; Cahaner, A.; Halevy, O. Featherless and feathered broilers under control versus hot conditions. 2. Breast muscle development and growth in pre-and posthatch periods. Poult. Sci. 2014, 93, 1076–1088. [Google Scholar] [CrossRef]
- Piestun, Y.; Patael, T.; Yahav, S.; Velleman, S.G.; Halevy, O. Early posthatch thermal stress affects breast muscle development and satellite cell growth and characteristics in broilers. Poult. Sci. 2017, 96, 2877–2888. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Sui, S.J.; Jiao, H.C.; Buyse, J.; Decuypere, E. Impaired development of broiler chickens by stress mimicked by corticosterone exposure. Comp. Biochem. Physiol. Part. A Mol. Integr. Physiol. 2006, 143, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Decuypere, E.; Buyse, J. Oxidative stress induced by corticosterone administration in broiler chickens (Gallus gallus domesticus) 1. Chronic exposure. Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2004, 139, 737–744. [Google Scholar] [CrossRef]
- Xiao, Y.; Wu, C.; Li, K.; Gui, G.; Zhang, G.; Yang, H. Association of growth rate with hormone levels and myogenic gene expression profile in broilers. J. Anim. Sci. Biotechnol. 2017, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataria, N.; Kataria, A.K.; Gahlot, A.K. Ambient temperature associated variations in serum hormones and interrelated analytes of broiler chickens in arid tract. Slov. Vet. Res. 2008, 45, 127–34616612. [Google Scholar]
- Sohail, M.U.; Hume, M.E.; Byrd, J.A.; Nisbet, D.J.; Ijaz, A.; Sohail, A.; Shabbir, M.Z.; Rehman, H. Effect of supplementation of prebiotic mannan-oligosaccharides and probiotic mixture on growth performance of broilers subjected to chronic heat stress. Poult. Sci. 2012, 91, 2235. [Google Scholar] [CrossRef]
- Lara, L.; Rostagno, M. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Loyau, T.; Bedrani, L.; Berri, C.; Métayer-Coustard, S.; Praud, C.; Coustham, V.; Mignon-Grasteau, S.; Duclos, M.J.; Tesseraud, S.; Rideau, N. Cyclic variations in incubation conditions induce adaptive responses to later heat exposure in chickens: A review. Animal 2015, 9, 76–85. [Google Scholar] [CrossRef]
- Jiang, S.; Mohammed, A.A.; Jacobs, J.A.; Cramer, T.A.; Cheng, H.W. Effect of synbiotics on thyroid hormones, intestinal histomorphology, and heat shock protein 70 expression in broiler chickens reared under cyclic heat stress-ScienceDirect. Poult. Sci. 2020, 99, 142–150. [Google Scholar] [CrossRef]
- Geraert, P.A.; Padilha, J.; Guillaumin, S. Metabolic and endocrine changes induced by chronic heatexposure in broiler chickens: Growth performance, body composition and energy retention. Br. J. Nutr. 1996, 75, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Mcfarlane, C.; Plummer, E.; Thomas, M.; Hennebry, A.; Ashby, M.; Ling, N.; Smith, H.; Sharma, M.; Kambadur, R. Myostatin induces cachexia by activating the ubiquitin proteolytic system through an NF-κB-independent, FoxO1-dependent mechanism. J. Cell. Physiol. 2006, 209, 501–514. [Google Scholar] [CrossRef]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Yancopoulos, G.D. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Forbes, D.; Jackman, M.; Bishop, A.; Thomas, M.; Kambadur, R.; Sharma, M. Myostatin auto-regulates its expression by feedback loop through Smad7 dependent mechanism. J. Cell. Physiol. 2006, 206, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Yueping, C.; Ping, W.; Wang, T.; Zhou, Y. MSTN, mTOR and FoxO4 are involved in the enhancement of breast muscle growth by methionine in broilers with lower hatching weight. PLoS ONE 2014, 9, e114236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, T.; Kitamura, Y.I.; Funahashi, Y.; Shawber, C.J.; Castrillon, D.H.; Kollipara, R.; De Pinho, R.A.; Kitajewski, J.; Accili, D. A Foxo/Notch pathway controls myogenic differentiation and fiber type specification. J. Clin. Investig. 2007, 117, 2477–2485. [Google Scholar] [CrossRef] [Green Version]
- Yi, C.; Yao, Z.; Sarkar, D.; Lawrence, M.; Sanchez, G.J.; Parker, M.H.; Macquarrie, K.L.; Davison, J.; Morgan, M.T.; Ruzzo, W.L. Genome-wide MyoD Binding in Skeletal Muscle Cells: A Potential for Broad Cellular Reprogramming. Dev. Cell 2010, 18, 662–674. [Google Scholar]
- Hennebry, A.; Berry, C.; Siriett, V.; O’Callaghan, P.; Chau, L.; Watson, T.; Sharma, M.; Kambadur, R. Myostatin regulates fiber-type composition of skeletal muscle by regulating MEF2 and MyoD gene expression. Am. J. Physiol. Cell Physiol. 2009, 296, C525–C534. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name 1 | Primer Sequence 2 5′-3′ | Product Size (bp) | GenBank Accession Number |
|---|---|---|---|
| β-actin | F: TGCTGTGTTCCCATCTATCG | 150 | NM_205518 |
| R: TTGGTGACAATACCGTGTTCA | |||
| mTOR | F: AAGGATGCTGACAAACGCTATGGA | 225 | XM_417614 |
| R: ACTGACTGACTGGCTGAGTAGGAG | |||
| myostatin | F: TACCCGCTGACAGTGGATTTC | 153 | NM_001001461 |
| R: GCCTCTGGGATTTGCTTGG | |||
| MyoD | F: GGAGAGGATTTCCACAGACAACTC | 113 | NM_204214 |
| R: CTCCACTGTCACTCAGGTTTCCT | |||
| Akt | F: GCTGGCATTGTTTGGCAAGATGT | 215 | NM_205055 |
| R: GCGGTTCCACTGGCTGAATAGG | |||
| FoxO4 | F: GCTCTTCTCACACCTGGCTCTC | 186 | XM_015278657 |
| R: TGGTTCTGCCTGCTGCTCTG | |||
| Smad3 | F: GCGTTCTGGTGCTCCATATCCTAC | 192 | NM_204475 |
| R: TCCTCTTCCGATGTGCCGTCTC | |||
| MAFbx | F: CAGTGAGCCAGCCTCTTGTGATG | 114 | NM_001030956 |
| R: TTCAGCCAGTGTGACAGTCTCAGT | |||
| MuRF1 | F: GCGAGCAGGAGGACAAGACAAG | 240 | XM_424369 |
| R: CAAGACTGACTGTGAAGGCATCCA |
| Items | TC 2 | HS 2 | p-Value |
|---|---|---|---|
| ADFI (g) | 153.30 a ± 1.17 | 127.30 b ± 1.76 | 0.02 |
| ADG (g) | 82.08 a ± 0.53 | 62.07 b ± 1.01 | <0.01 |
| FCR | 1.87 b ± 0.01 | 2.05 a ± 0.01 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhang, M.; Feng, J.; Zhou, Y. Myostatin and Related Factors Are Involved in Skeletal Muscle Protein Breakdown in Growing Broilers Exposed to Constant Heat Stress. Animals 2021, 11, 1467. https://doi.org/10.3390/ani11051467
Li X, Zhang M, Feng J, Zhou Y. Myostatin and Related Factors Are Involved in Skeletal Muscle Protein Breakdown in Growing Broilers Exposed to Constant Heat Stress. Animals. 2021; 11(5):1467. https://doi.org/10.3390/ani11051467
Chicago/Turabian StyleLi, Xiumei, Minhong Zhang, Jinghai Feng, and Ying Zhou. 2021. "Myostatin and Related Factors Are Involved in Skeletal Muscle Protein Breakdown in Growing Broilers Exposed to Constant Heat Stress" Animals 11, no. 5: 1467. https://doi.org/10.3390/ani11051467
APA StyleLi, X., Zhang, M., Feng, J., & Zhou, Y. (2021). Myostatin and Related Factors Are Involved in Skeletal Muscle Protein Breakdown in Growing Broilers Exposed to Constant Heat Stress. Animals, 11(5), 1467. https://doi.org/10.3390/ani11051467